
沿江双季稻北缘区机插早稻的超高产群体特征
2021-04-21 10:44:36朱铁忠柯健姚波陈婷婷何海兵尤翠翠朱德泉武立权
中国农业科学 2021年7期
朱铁忠,柯健,姚波,陈婷婷,何海兵,尤翠翠,朱德泉,武立权,2✉
1安徽农业大学,合肥 230036;2江苏省现代作物生产协同创新中心,南京 210095
0 引言
【研究意义】双季稻是我国南方典型的水稻种植模式,较单季稻可实现周年水稻增产59%左右,在当前可耕地面积急剧下降的背景下,是保证我国粮食安全的重要稻作方式[1-2]。安徽沿江地区是我国双季稻主要种植区域,年播种面积20×104hm2,但由于地处双季稻北缘,生长季节短、温光资源紧张[3],早季稻倒春寒现象发生普遍,加之推行“早籼晚粳”品种搭配模式进一步压缩了早稻生育进程,导致早稻产量一般较晚稻低1 t·hm-2以上,极大地影响了沿江地区双季稻周年产量的提高[4-6]。近年来,随着农机农艺技术的改进、集中育秧及专业合作社的发展,机插逐渐成为双季稻主要的种植方式,极大地提高了种植效率、降低了劳动力成本,促进水稻高产群体构建,有利于水稻稳产高产[7-9]。但这种方式在一定程度上较传统手栽、抛栽方式缩短了水稻生育进程,尤其不利于沿江双季稻北缘区生育期限制下的早稻产量提高[10-11]。因此,在适宜生育期下,选用高效利用温光资源的机插高产品种是当前提高沿江双季稻北缘区早稻产量的重要思路,对稳定和发展我国双季稻生产具有重要意义。【前人研究进展】当前关于南方双季早稻机插高产品种筛选及其群体特征已有大量研究[12-15]。曾勇军等[13]认为高产类型早稻生育期110—115 d,穗型结构为每穗粒数100—130,千粒重27—29 g,单穗干重2.5 g以上。吕伟生等[14]和WU等[15]进一步阐明,机插高产类型早稻品种具有分蘖力中等、成穗率较高、全生育期特别是中后期干物质积累量大、中后期叶面积指数高和穗大粒多等基本特征。然而,随着育种技术的飞速发展,人们对水稻产量水平的期望已由传统的高产逐渐向更高产和超高产转变,且近年来农机农艺农信技术的不断融合,也为水稻产量的进一步提升带来了契机。目前,基于单季中稻的机插超高产群体形成机制已基本明确,且基本认为建立高光效群体、培育大穗、平衡源库关系、稳定结实率是机插中稻超高产的核心栽培技术[16-17]。【本研究切入点】考虑到不同季节温光特征和品种特性对水稻产量形成具有显著的互作效应[18-19],机插模式下双季早稻超高产群体形成特征与已有的常规中稻机插超高产栽培方式或可能存在差异,但相关研究并不系统。【拟解决的关键问题】本研究采用前期筛选获得的 9个早稻高产品种为供试材料,分析机插下不同品种的产量及构成、干物质积累、叶面积指数和群体光截获的差异,系统研究温光资源高效利用超高产品种的群体共性特征,以期为沿江双季稻北缘区机插超高产早稻品种的选育与精确定量栽培提供参考。
1 材料与方法
1.1 试验地点及材料
试验于 2018—2019年在安徽农业大学郭河试验基地(31°48′N,117°23′E)进行。试验地土壤为砂壤土,移栽前 0—20 cm 土壤主要理化参数为全氮2.03 g·kg-1,有机质 32.36 g·kg-1,有效磷 24.80 mg·kg-1,速效钾211.42 mg·kg-1,pH 5.11。试验期间的气象数据由基地安装的小型自动气象站(Watch Dog 2900ET,SPECTRUM,USA)提供,2年水稻生长期太阳辐射规律基本一致,2018年水稻生长期平均温度较2019年高0.7℃(图1)。
本试验采用 9个高产抗倒伏品种为供试品种材料,包括 4个杂交籼稻品种,分别为株两优 2013(ZLY2013)、陆两优 35(LLY35)、株两优 829(ZLY829)、陵两优942(LLY942),5个常规籼稻品种,分别为浙辐203(ZF203)、洪优早1号(HYZ1)、中嘉早17(ZJZ17)、湘早籼24号(XZX24)和中早35(ZZ35)。以上材料是由中国水稻所提供的 28个早稻品种在当地种植,根据产量、抗倒伏和抗病性综合筛选的最优品种类型。
1.2 试验设计
采用随机区组设计,9个品种,3次重复,共27个小区,单个小区面积75 m2(3.75 m×20 m)。所有处理施氮量为150 kg·hm-2,基肥∶分蘖肥∶穗肥=4∶4∶2,所有处理磷、钾施用量相同,分别为75 kg·hm-2和120 kg·hm-2,均一次性基施。分别于2018年3月28日和2019年3月25日播种,大棚软盘旱育秧,每盘播种芽谷120 g,于2018年4月19日和2019年4月24日采用井关PZ80D-25高速插秧机机插,移栽行株距为25 cm×14 cm,常规稻每穴4—5苗,杂交稻每穴2—3苗。采用常规水分管理,返青后保持3—5 cm田面水层,够苗期排水晒田,之后干湿交替灌溉,于成熟前一周排水自然落干。病、虫、草害防治同当地高产管理。
1.3 测定项目及方法
1.3.1 生育期判定标准 参照 XU 等[12]标准结合镜检的办法,以小区中80%的植株幼穗长0.5—1.0 mm为穗分化期,以50%的稻穗露出叶鞘为抽穗期,以每穗有90%籽粒黄熟,且稻穗基部青谷粒坚硬为成熟期。
1.3.2 冠层 PAR参数和叶面积指数 分别于穗分化期和抽穗期,在典型晴天的 11:00—13:00,使用SunScan冠层分析仪(Delta,USA)测定冠层光合有效辐射(photosynthetically active radiation,PAR)。将探头置于距冠层顶部30 cm处,垂直向上测量冠层顶部入射PAR(总光截获,PAR0),垂直向下测量冠层反射PAR(PAR1),再将探头置于冠层底部距土壤表面 5 cm处,垂直向上测量冠层底部入射 PAR(PAR2)[20]。每次测定PAR值后,各小区根据普查茎蘖数选取代表性植株3穴,使用Li-3000型自动叶面积仪(LI-COR,USA)测定植株叶面积,并计算叶面积指数。
1.3.3 干物质积累 于穗分化期、抽穗期和成熟期,根据平均茎蘖数,各小区取代表性植株5穴。将样品分为茎鞘、叶、穗(抽穗、成熟期),105℃杀青30 min,80℃烘干至恒重,测定各部分干物质重。
1.3.4 抽穗期粒叶比 粒叶比包括颖花数/叶面积、实粒数/叶面积、粒重/叶面积,其中颖花数、实粒数、粒重分别指总颖花数、总实粒数,此处叶面积指抽穗期的叶面积[21]。
1.3.5 实际产量和产量结构调查 于成熟期,各小区取代表性植株5穴考种,测定穗粒数、结实率、千粒重[22]。在每个小区中心未采样处实割100穴,晾干后测定谷物质量和含水量,然后按含水量 13.5%折算实际产量。
1.4 计算方法
式中,CGR(crop growth rate)表示群体生长速率,W1和W2为前后2次测定的干物质重,t1和t2为前后2次测定的时间。
式中,IPAR(the intercepted photosynthetically active radiation,μmol·m-2·s-1)表示有效光截获量,PAR0表示冠层顶部入射PAR,PAR1表示冠层反射PAR,PAR2表示冠层底部入射 PAR,I(the interception rate of photosynthetically active radiation,%)表示有效光截获率,PUE(the photosynthetically active radiation use efficiency,g·MJ-1)表示光截获利用率,TPAR(the total photosynthetically active radiation,MJ·m-2)表示某生育阶段总光合辐射,DM(the dry biomass,g·m-2)表示某生育阶段干物质积累量。
1.5 分析与统计方法
应用 Excel 2016进行数据输入、整理,使用OriginPro 9.1进行作图,用SPSS 22.0软件进行数据统计分析。
2 结果
2.1 水稻主要农学参数的方差分析
年份间水稻实际产量和日产量差异显著(表1),这可能与2年水稻生长季平均温度不同有关(图1)。品种间水稻实产、日产量、成熟期干物质积累量和抽穗期叶面积指数有显著差异。品种对相关农学指标的影响整体高于年份,且年份与品种对水稻主要农学指标的影响无互作效应,因此本研究采用2年数据的平均值进行下列分析。

表1 水稻主要农学参数的方差分析Table 1 Analysis of variance of main agronomic parameters of rice
2.2 水稻产量及构成因素
采用欧氏距离法对不同品种的实际产量进行系统聚类(图 2),根据聚类结果将不同品种分为超高产(9.1—9.5 t·hm-2)、更高产(8.1—8.6 t·hm-2)和高产(7.6—7.8 t·hm-2)3种产量类型。其中超高产类型品种为株两优2013、陆两优35、浙辐203和株两优829,更高产类型品种为洪优早1号、陵两优942和中嘉早17,高产类型品种为湘早籼24号和中早35。
不同产量类型间的水稻实际产量存在显著差异(表2),与更高产和高产类型品种相比,超高产类型品种实际产量分别高 12.0%和 20.8%。对产量构成因素进一步分析发现,不同产量类型间的水稻有效穗数和结实率无显著差异。与更高产与高产类型品种相比,超高产类型品种显著提高了每穗粒数、总颖花量和千粒重。由此可见,在保证有效穗数充足和结实率稳定的前提下,通过提高每穗粒数来增加总颖花量,同时提高千粒重,是机插早稻品种超高产产量形成的主要原因。
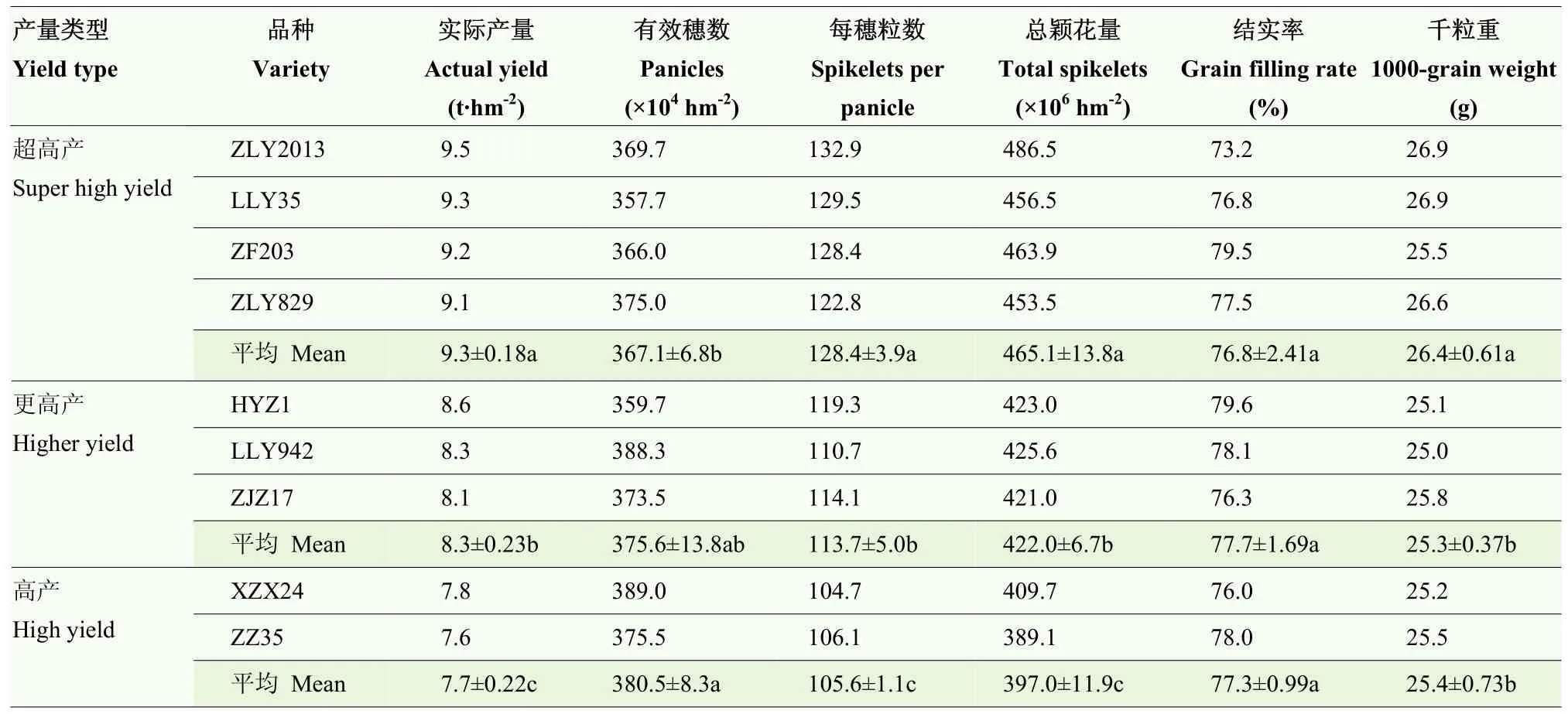
表2 不同类型水稻品种产量及构成因素Table 2 Grain yield and its components of different yield types of rice
2.3 水稻主要生育时期和日产量
不同产量类型间的水稻生育进程无显著差异(表3),其中移栽—穗分化期的天数为31—33 d,穗分化—抽穗期为25—28 d,抽穗—成熟期为22—25 d,移栽—成熟期的天数为81—83 d。结果表明,不同产量类型的水稻产量差异并不是由生育期造成的,而可能与生产效率有关。进一步比较日产量发现,不同产量类型间的水稻日产量存在显著差异,且表现超高产类型品种(82.4 kg·hm-2·d-1)显著高于更高产(74.7 kg·hm-2·d-1)和高产类型品种(68.8 kg·hm-2·d-1)。与更高产和高产类型品种相比,超高产类型品种日产量分别高出 10.2%和 19.8%。因此,除生育期以外,日产量是机插超高产早稻品种的重要筛选指标。
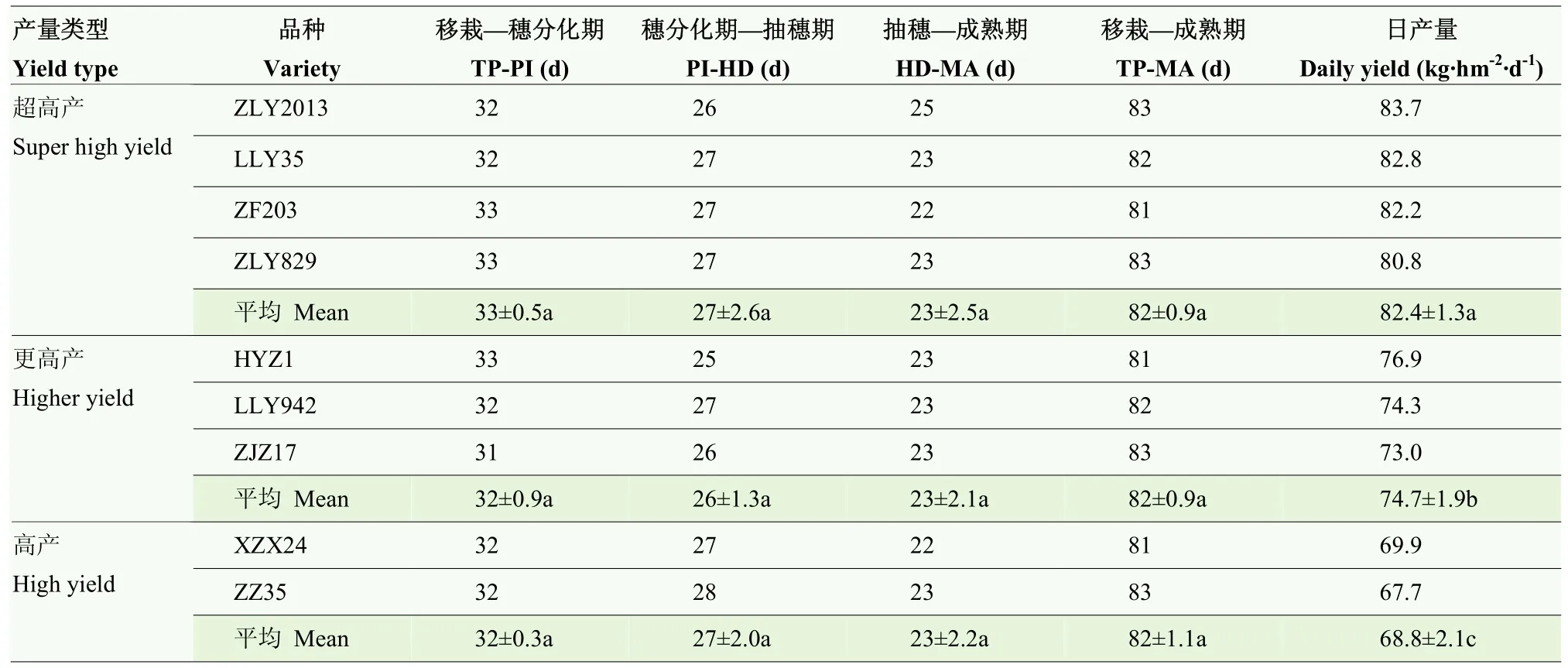
表3 水稻主要生育时期天数及日产量Table 3 The days of main growth duration and daily grain yield of rice
2.4 水稻干物质生产特性
不同产量类型间的水稻(阶段)干物质积累量和(阶段)群体生长速率具有显著差异(表 4)。超高产类型品种总干物质积累量为 16.6 t·hm-2,较更高产和高产类型品种分别高出11.4%和14.5%,差异显著,这主要是由于更高的中后期干物质阶段积累。与高产类型品种相比,超高产类型品种分别增加水稻穗分化—抽穗期和抽穗—成熟期阶段干物质积累量 15.1%和27.1%,差异显著;与更高产类型品种相比,超高产类型品种显著增加了水稻抽穗—成熟期阶段干物质积累量 41.5%。然而值得注意的是,与更高产与高产类型品种相比,超高产类型品种均显著降低了水稻移栽—穗分化期阶段干物质积累量。
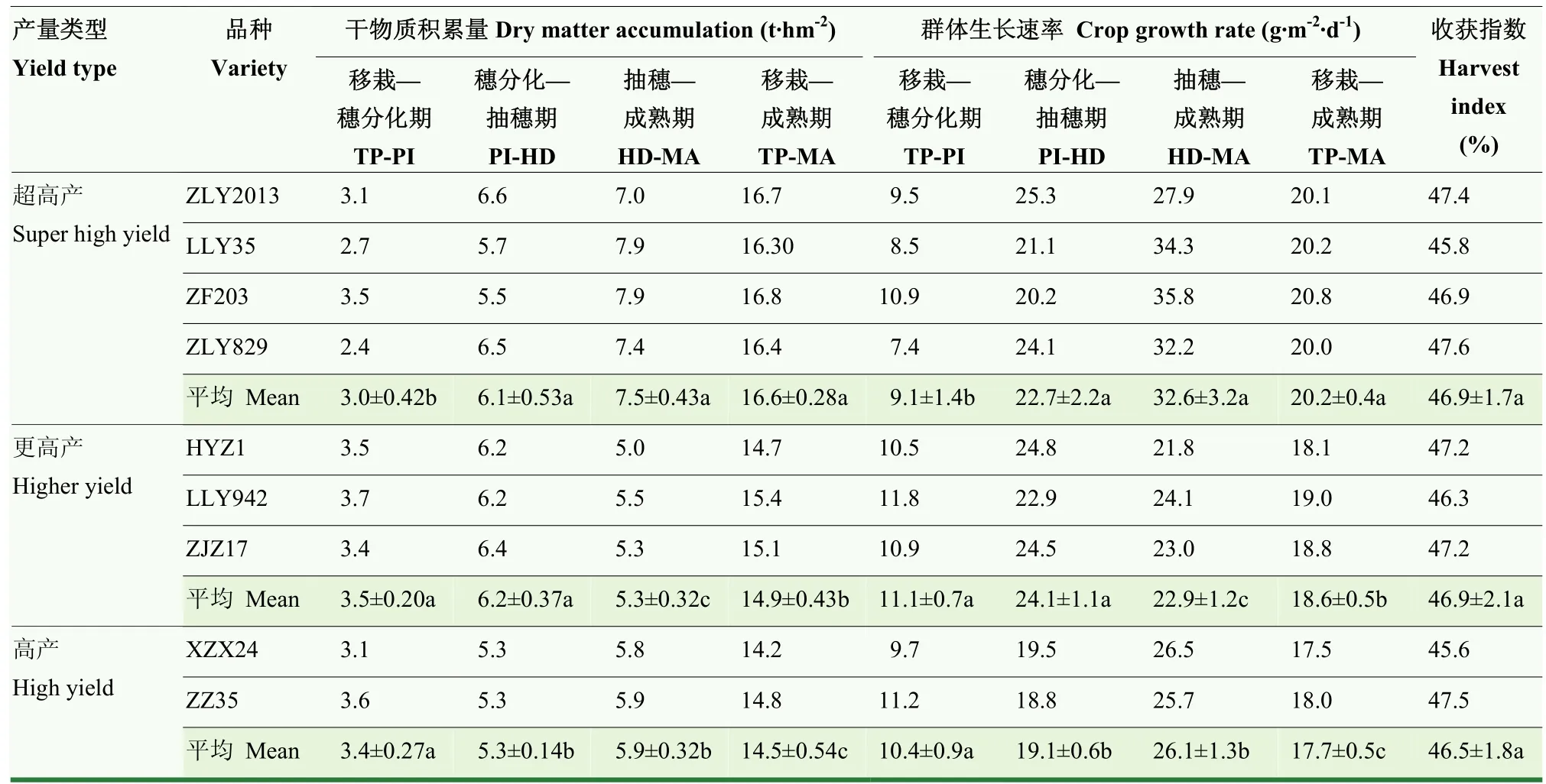
表4 不同产量类型品种水稻干物质生产特性Table 4 Dry matter production characteristics of different yield types of rice
不同产量类型间的水稻群体和阶段生长速率与对应的干物质积累规律基本一致,这与不同类型间无明显差别的生育进程有关。另外,不同产量类型间的水稻收获指数为46.5%—46.9%,无显著差异,表明干物质积累是影响不同类型水稻产量差异的主要原因。
2.5 水稻群体光合生产特性
2.5.1 叶面积指数和光截获的特征 超高产类型品种具有更高的穗分化期和抽穗期叶面积指数、有效光截获量(图 3)。与更高产和高产类型品种相比,超高产类型品种分别提高水稻穗分化期叶面积指数9.4%和18.4%,提高抽穗期叶面积指数12.5%和7.5%。同时,超高产类型品种分别提高水稻穗分化期有效光截获量0.6%和4.6%,提高抽穗期有效光截获量12.1%和15.2%。另外,水稻穗分化期和抽穗期的叶面积指数与有效光截获量均呈极显著正相关关系。由此可见,超高产类型品种在水稻穗分化期与抽穗期具有更高的光截获能力,与其在这2个时期较高的叶面积指数有关。
2.5.2 阶段光截获利用效率 不同产量类型间的水稻穗分化—抽穗期和抽穗期—成熟期阶段光截获利用率(PUE)均具有显著差异(图 4)。与高产类型品种相比,超高产类型品种分别显著增加了穗分化—抽穗期和抽穗—成熟期阶段PUE 23.7%和39.8%;与更高产类型品种相比,超高产类型品种显著增加了水稻抽穗—成熟期阶段PUE 50%。超高产类型品种不仅表现了更高的光截获效率,同时具有更高的光截获利用率。
2.5.3 光截获与每穗粒数及千粒重的关系 水稻穗分化期有效光截获量与每穗粒数,以及抽穗期有效光截获量与千粒重,均呈显著的正相关关系(图 5)。因此,超高产类型品种最高的产量成因与穗分化期和抽穗期更高的有效光截获量有关。另外,不同产量类型水稻的每穗粒数和千粒重对阶段有效光截获量的响应效率(方程斜率)并不一致。从每穗粒数来看,超高产和更高产类型品种具有更高的有效光截获量响应效率;从千粒重来看,超高产类型品种表现最高的有效光截获量响应效率。另外,上述不同产量类型间的水稻每穗粒数和千粒重光截获量响应效率与其阶段PUE规律基本一致。因此,在有效光截获量对品种穗型的影响过程中,品种的光截获利用率可能还存在一定的正向调节效应。
2.6 水稻抽穗期粒叶比
不同产量类型间的水稻粒叶比(颖花/叶、实粒/叶和总粒重/叶)具有显著差异(表 5)。超高产类型品种的颖花/叶、实粒/叶和总粒重/叶分别为0.69 cm-2、0.52 cm-2和18.06 mg·cm-2,均显著高于高产类型品种。与更高产类型品种相比,超高产类型品种显著提高了总粒重/叶的比值。另外,水稻穗分化—成熟期PUE与总粒重/叶呈极显著正相关关系(图6)。由此可见,提高水稻穗分化—成熟期的光截获利用率有利于平衡水稻源库关系。

表5 不同产量类型水稻品种的抽穗期粒叶比Table 5 Grain-leaf area ratio of different types of rice at heading stage
3 讨论
3.1 机插超高产早稻的产量及构成因素
邹应斌等[23]提出高产双季早稻产量可达8.1 t·hm-2,这与吕伟生等[14]认为早稻高产水平为 8.3 t·hm-2的结果基本一致。LOBELL等[24]通过对高产栽培条件下早晚稻产量潜力的评估,认为我国双季早、晚稻的高产潜力相近,均约为9.5—9.8 t·hm-2。然而,沿江双季稻北缘区由于受水稻品种和温光资源的限制,传统种植模式下早稻产量仅为5.5—6.4 t·hm-2左右[25]。本研究中的9个早稻品种产量区间为7.6—9.5 t·hm-2,通过系统聚类分析进一步分为 9.1—9.5 t·hm-2、8.1—8.6 t·hm-2和7.6—7.8 t·hm-23个产量等级,均达到了本区域高产水平,且第一类基本达到了文献报道的早稻产量潜力。因此,我们将3类产量水平分别定义为超高产、更高产和高产水平。
研究表明,适当减少分蘖,增加成穗率,以培育结实期高光效群体;同时增加每穗粒数,扩大库容,促进源库平衡,是当前超高产栽培的基本理论[26-27]。因此,在常规中稻种植中,选用大穗型品种,配合钵苗摆栽的群体优化栽植技术,可不断提高产量水平[7,28]。然而由于早稻营养生长期较低的温度限制,以及较短的生殖生长期,影响水稻分蘖发生和大穗形成,在过去很长的时期内基于“穗大粒多”的早稻超高产栽培理论难以实现。目前关于早稻高产栽培的理论还仍以提高有效穗数为主。WU等[15]通过比较早稻和晚稻产量形成差异,认为早稻产量显著低于晚稻的主要原因是显著低的有效穗数,并提出培育分蘖能力强的早稻品种是未来育种的重要发展方向。吕伟生等[11]认为机插早稻取得超高产的关键是在足穗的前提下提高每穗粒数并保证较高的结实率,提出超高产群体需保证有效穗数345 m-2以上。随着超级稻理想株型构建与超高产育种理论的应用,从品种自身出发,选用“穗大粒多”的品种为实现早稻超高产攻关创造了可能。本研究中,超高产类型水稻平均产量为9.3 t·hm-2,分别较更高产和高产增产 12.0%和 20.8%,主要由于显著增加了每穗粒数、总颖花量和千粒重。这与当前单季中稻和双季晚稻超高产形成规律基本是一致的[16-17],是基于品种优化的超高产理论在早稻季的一次重要验证。此外,适当提高千粒重是早稻品种获得超高产的重要指标,这在通常的高产栽培研究中往往是较容易忽视的。
在南方亚热带气候导致的水稻生育期限制条件下,日产量是品种温光利用效率的重要评价指标[19,29]。因此,研究沿江双季稻北缘区早稻品种的日产量特征,其意义更加突出。当前基于高日产量的早稻品种筛选及其群体共性特征探讨已有研究[12,13],XU 等[12]提出日产量为 100 kg·hm-2·d-1是中国中部地区直播短生育期双季早稻品种的理想数值。与直播相比,机插稻的生育期可适当延长,对应允许的适宜日产量必然有所不同或下降。吕伟生等[14]指出高产机插早稻适宜日产量为 75.0—78.3 kg·hm-2·d-1。本研究发现,高产类型水稻品种日产量为 73.0—76.9 kg·hm-2·d-1,与之前研究基本一致,此外本研究进一步提出了超高产类型水稻品种日产量为 80.8—83.7 kg·hm-2·d-1。另外,本研究中超高产类型水稻品种的全生育期为110.7—115.3 d,是本区域“早籼晚粳”茬口衔接的早稻安全生育期[30],因此提出的超高产类型水稻品种日产量标准具有重要的生产指导意义。
3.2 机插超高产早稻的群体形成特征
提高收获指数和群体干物质积累量是增加水稻产量的主要途径。然而,从品种角度而言,通过提高当前半矮杆品种的收获指数来增产是难以实现的[31],本研究中不同品种的收获指数(46.5%—46.9%)无显著差异,也很好地证实了这一点,这与XIONG等[32]在早稻上的研究结果基本一致。因此,增加群体干物质积累量被广泛认为是提高水稻产量形成的主要方向。本研究表明,3种产量类型水稻成熟期的干物质积累量为超高产>再高产>高产,与产量规律一致。超高产类型品种更高的干物质积累主要是显著增加了中后期群体生长速率,进而增加了中后期干物质积累量。然而,与之相反的是,本研究中超高产类型水稻品种移栽—穗分化期干物质生产速率和阶段积累量显著低于其他2种产量水平,这可能是由于该类品种较低的分蘖能力有关,其显著低的有效穗数也很好地证明了这一点。这与杨惠杰等[33]研究的水稻超高产干物质积累规律基本一致。因此,本研究认为,进一步提高中后期,尤其是灌浆期的干物质积累量和群体生长速率,是机插早稻由高产向超高产攻关的主要群体形成生理机制。这与XU等[12]在直播方式上的研究结果基本一致。
唐海明[34]、童平[35]、HUANG[36]、LAZA[37]等在中稻和晚稻上研究证实水稻穗分化—抽穗期以及抽穗—成熟期阶段干物质积累量分别与穗粒数以及千粒重呈显著正相关。本研究表明,水稻穗分化期群体有效光截获量与每穗粒数,以及抽穗期群体有效光截获量与千粒重均呈明显的正相关关系。因此,本研究中不同产量类型水稻每穗粒数和千粒重的差异,主要是由于群体有效光截获量影响水稻对应时期干物质积累而引起的。另外,本研究表明,水稻穗分化期、抽穗期叶面积指数与群体有效光截获量呈极显著正相关关系,这与HOU等[20]研究结果基本一致。因此,本研究中超高产类型品种表现更高的每穗粒数和千粒重,可能是由于其更高的穗分化期和抽穗期叶面积指数,进而增加了对应阶段的群体有效光截获量,从而显著增加了水稻中后期干物质积累。然而,除冠层的有效光截获量以外,作物的光截获利用率对干物质积累量同样具有重要的影响。本研究中,不同产量类型间水稻中后期光截获利用率表现超高产>高产>更高产。HORIE等[38]认为品种的光截获利用率与其源库生产特性有关,库限制性品种一般不利于光截获利用率的提高。超高产品种更高的中后期光截获利用率可能是由于其更高的库容量(穗粒数)和库活力(千粒重)。因此,由于光截获利用率的正向调节效应,超高产类型品种在单位有效光截获量下,或表现更高的增产潜力。当前,超高产类型品种的穗粒数和千粒重表现了更高的有效光截获响应效率,很好地证实了这一猜想。此外,我们注意到,水稻穗分化—成熟期的光截获利用率与总粒重/叶呈极显著正相关,这可能是由于提高了光截获利用率,进一步提高光能向水稻穗数和千粒重的转化效率。综上所述,本研究认为,更高的光截获和利用率,增加水稻穗分化—抽穗期以及抽穗—成熟期干物质生产能力,提高水稻穗粒数和千粒重,促进源库平衡,是实现机插早稻超高产的重要光合生理机制。
3.3 机插超高产早稻品种的氮素管理探讨
由于地处沿江双季稻北缘区的特殊生产环境[3-6],在早稻生产上为抵消营养生长期低温对产量的损失,当地农户习惯采用“重施基肥、早施攻蘖肥”的方式来提高早稻产量[39]。然而,这种依靠增加穗数来提高产量的方式,不仅增加了肥料、种子和机械投入,还带来了一定的环境风险。本研究提出的超高产类型早稻品种较常规品种一定程度上降低了有效穗数的要求,在栽培技术上可适当降低基蘖肥和栽植密度,具有重要的经济和环境效益。但值得注意的是,与常规品种相比,本研究中超高产类型水稻品种需要建立更高的穗分化期和抽穗期叶面积指数,以保证中期高光效群体的构建和后期的灌浆结实。因此,改进现有的施肥模式,减少基蘖肥用量,适当调节穗肥氮素的施用时期和施用量,对进一步提高机插超高产类型水稻品种产量具有重要意义。
4 结论
本研究初步明确机插超高产早稻品种的共性群体特征为日产量高(80.8—83.7 kg·hm-2·d-1),较高的每穗粒数(124—132)和总颖花量(45.2×103—47.9×103m-2),千粒重大(25.8—27.0 g),更高的穗分化期和抽穗期的叶面积指数(5.6—6.0,7.1—7.3),粒叶比高。并以此筛选适合沿江双季稻北缘机插超高产早稻品种4种,分别为株两优2013、陆两优35、浙辐203、株两优829。
猜你喜欢
烟草科技(2022年9期)2022-09-24 08:58:58
农产品市场周刊(2022年14期)2022-08-10 07:09:10
农业科技与信息(2021年23期)2021-12-06 01:14:10
今日农业(2021年17期)2021-11-26 23:38:44
古今农业(2021年1期)2021-07-20 07:52:08
山西农业科学(2021年4期)2021-04-19 08:51:56
中国锰业(2021年3期)2021-03-31 07:32:38
山西农业科学(2018年7期)2018-07-13 06:00:40
天然气与石油(2015年1期)2015-02-28 17:00:54
中国林副特产(2012年4期)2012-02-24 09:59:58
