
不同无性系四川桤木嫁接苗生物量及其分配特征
2021-04-07 07:49:48白文玉冯茂松铁烈华汪亚琳高嘉翔戴晓康
南京林业大学学报(自然科学版) 2021年2期
白文玉,冯茂松,铁烈华,汪亚琳,高嘉翔,赖 娟,戴晓康
(四川农业大学林学院,长江上游森林资源保育与生态安全国家林业和草原局重点实验室,长江上游林业生态工程四川省重点实验室,四川 成都 611130)
四川桤木(Alnusledgeriana)为桦木科(Betulaceae)桤木属(Alnus)落叶乔木,根系具有根瘤,可有效改良土壤,其适应性强、繁殖容易、生长迅速且材质优良,生态和经济价值较高,是集用材、造纸和生态防护等为一体的多功能树种,已经成为我国西南地区的速生用材树种和四川省重要的乡土树种,以及长江防护林工程及国家造林项目的重要树种[1-4]。
林木优良无性系的选育是林业研究的重要内容之一[5]。生物量是植物的基本生物学特征和功能性状之一,反映了植物的物质积累状况和对环境资源利用的能力[6-7]。植物生物量及其在各器官(枝、叶、干、根)的分配特征受植物体自身遗传特性和外界环境条件的共同影响[8-10],反映了其对外界环境的适应策略及能力,是衡量林木优良无性系的重要指标[11-12]。四川桤木不同无性系长期处于各种生境中,受遗传变异和自然选择的影响,其生长情况、对环境的适应能力和生物量分配特征等可能发生了明显变化,这为其良种育苗提供了繁育材料。已有研究主要集中在混交林营造[13]、种源选择[14]、生物固氮[15]以及光合特征[16]等方面,但对四川桤木种子园建设中不同无性系生物量及其分配特征的研究还鲜见报道。为此,本研究依托国家级桤木种子园和良种基地建设平台,调查无性系嫁接幼苗各器官生物量,研究四川桤木不同无性系1年生嫁接幼苗各器官的生物量及其分配特征,探索四川桤木无性系嫁接幼苗对环境的适应策略和能力,为其优良无性系的筛选提供基础数据。
1 材料与方法
1.1 试验地概况
试验地位于四川省平昌县元山镇的桤木良种育苗基地(107°7′57.4″E,31°37′18.6″N),属于中亚热带湿润季风气候,海拔650 m,年均降雨量1 200 mm,早春时具有明显的季节性干旱现象。年均气温为16.8 ℃,四季分明,年均日照时间为1 239.3 h,无霜期297 d。基地坡向为东南方,坡度约10°,具有灌溉条件,光照充足,土壤为紫色壤土,呈弱酸性,土层深度大于40 cm。
1.2 试验材料
2015年12月,在四川省宣汉县、巴中市恩阳区和金堂县分别选择出林相整齐的15~20 年生四川桤木纯林,采用优势木对比法筛选出自然整枝良好、树干通直、无病虫害的四川桤木优树共34株[宣汉(编号XH)16株,恩阳(编号EY)11株,金堂(编号JT)7株],每株采集穗条约200枝。采集穗条时选择树冠中上部生长健壮、叶芽饱满、粗度3~5 mm,长度30~50 cm的1年生枝条,编号分别为宣汉1~16(XH1~XH16)、恩阳1~11(EY1~EY11)、金堂1~7(JT1~JT7)。将采集好的穗条打捆,用湿报纸包裹,登记好编号,并尽快带回实验室,保存于4 ℃冰柜中待用。
2015年11月,将苗圃地划分为3个区组,每个区组包含若干个1.2 m × 6.0 m的小区,选择生长健壮、长势基本一致、无病虫害的当年生四川桤木实生营养袋苗(种子采自平昌县四川桤木母树林),按株行距20 cm × 20 cm进行栽植,作为砧木。2016年1月初,采用随机区组设计方法,将采集的四川桤木优树的穗条嫁接于长势基本一致、无病虫害的砧木上,每小区内约嫁接100株(成活率45%~60%),同株优树上采集的穗条重复3次(嫁接于3个小区)。挂上对应的标号牌,登记好编号,搭建小拱棚,及时浇水、除草。当嫁接幼苗成活并经过炼苗后,去除小拱棚,此后根据土壤水分条件适当浇水,定时除草,常规田间管理。
1.3 试验方法
2016年8月中旬,测定小区内所有嫁接幼苗(平均约50株)的株高和基径,根据测定结果在每个小区内选出3株平均株作为待测植株,将待测植株全株挖起测定各器官生物量。
将待测植株按器官(主干、侧枝、叶、根)进行分离,用精度为0.01 g的电子天平分别称量待测植株主干、侧枝和叶的鲜质量。将根系带回实验室洗净表土,利用Epson Expression 10000XL 110数字化扫描仪(Seiko Epson Corp.,Nagano,Japan)测定根系直径(d),按照根系粗度进行分级。根径分级标准为细根(d<1 mm)、中根(1 mm ≤d≤ 2 mm)、粗根(d>2 mm)。根系扫描后,将植株的主干、侧枝、叶、粗根、中根和细根放入烘箱105 ℃杀青30 min,在烘箱里65 ℃烘干至质量恒定,分别称其干质量。
分别计算植株的地上部分生物量(主干干质量+侧枝干质量+叶干质量)、地下部分生物量(粗根干质量+中根干质量+细根干质量)、总生物量(地上部分生物量+地下部分生物量)、叶生物量比(叶生物量/总生物量)、侧枝生物量比(侧枝生物量/总生物量)、主干生物量比(主干生物量/总生物量)、粗根生物量比(粗根生物量/总生物量)、中根生物量比(中根生物量/总生物量)、细根生物量比(细根生物量/总生物量)、根冠比(地下部分生物量/地上部分生物量)。植株叶、侧枝和主干的相对含水量计算公式:相对含水量=(鲜质量-干质量)/鲜质量×100%。
1.4 数据处理
用Excel 2010和SPSS 25.0统计分析软件(IBM SPSS,Chicago,USA)对数据进行整理和统计分析,采用单因素方差分析(One-way ANOVA)计算无性系嫁接幼苗鲜质量、相对含水量、生物量等指标的平均值和标准偏差,并用Excel 2010作图。采用系统聚类方法对四川桤木无性系嫁接幼苗生物量及其分配进行聚类分析。
2 结果与分析
2.1 无性系嫁接幼苗鲜质量和含水量特征
经测定,四川桤木无性系嫁接幼苗鲜质量和含水量特征见图1、图2。
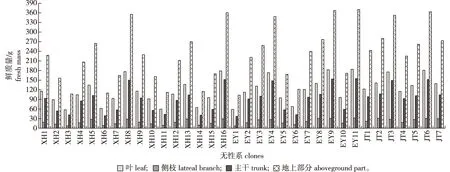
图1 四川桤木不同无性系地上部分鲜质量的差异Fig.1 Differences of the aboveground parts of fresh mass in different Anus ledgeriana clones
无性系嫁接幼苗地上部分各器官鲜质量表现为叶>主干>侧枝,EY11的叶、侧枝、主干以及地上部分鲜质量均最大,XH3叶片鲜质量最小(57.59 g),EY1侧枝、主干和地上部分鲜质量最小。XH6、XH8、XH16、EY1、EY4、EY9、EY11、JT3、JT6地上部分各器官相对含水量表现为叶>主干>侧枝,其他无性系地上部分各器官相对含水量表现为叶>侧枝>主干。无性系嫁接幼苗叶片相对含水量为62.20%~72.79%,侧枝相对含水量为49.23%~69.20%,主干相对含水量为50.38%~54.41%。

图2 四川桤木不同无性系地上部分相对含水量的差异Fig.2 Differences of the aboveground parts of relative water content in different A.ledgeriana clones
2.2 无性系嫁接幼苗生物量特征
各无性系幼苗生物量见图3、图4,无性系XH2、XH7、XH10、XH14、XH15、EY5、EY6、EY10地上部分各器官生物量表现为叶>主干>侧枝,其他无性系地上部分各器官生物量表现为主干>叶>侧枝,其中EY11的叶、侧枝、主干以及地上部分生物量均最大,分别为68.72、15.16、74.73和159.06 g,EY1的叶、侧枝、主干以及地上部分生物量均最小(图3)。四川桤木不同无性系地下部分生物量的差异见图4。

图3 四川桤木不同无性系地上部分生物量的差异Fig.3 Differences of the aboveground parts of biomass in different A.ledgeriana clones
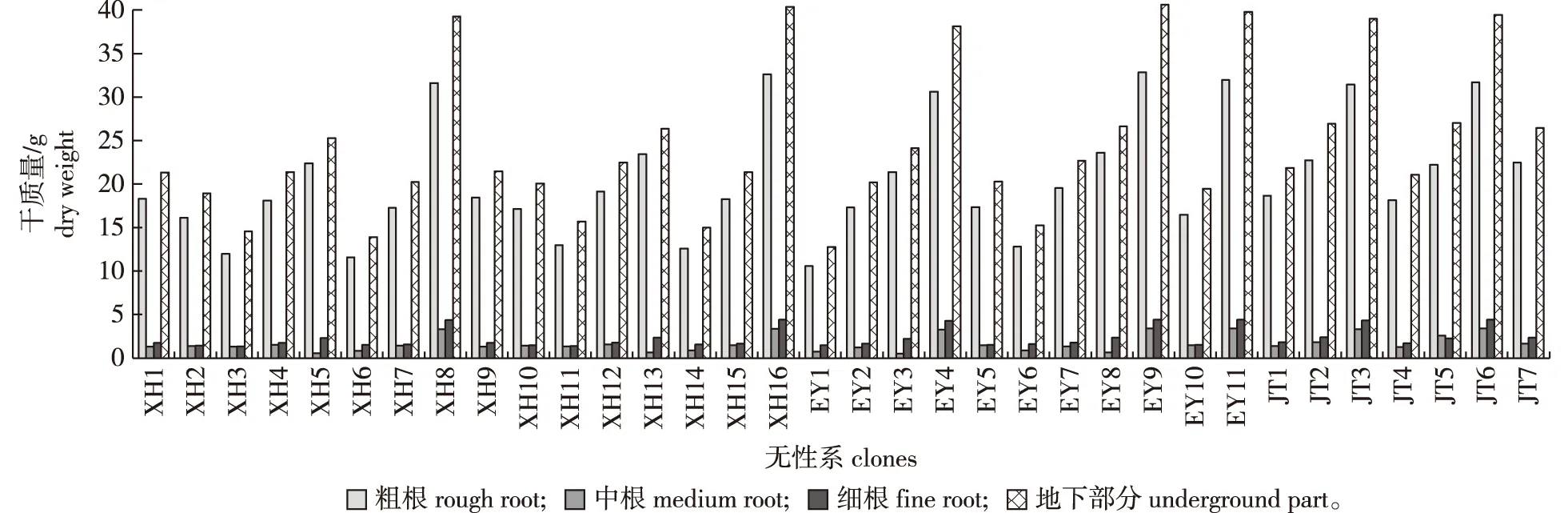
图4 四川桤木不同无性系地下部分生物量的差异Fig.4 Differences of the underground parts of biomass in different A.ledgeriana clones
由图4可知,无性系JT5地下部分各根径生物量表现为粗根>中根>细根,其他无性系地下部分各根径生物量表现为粗根>细根>中根,四川桤木无性系嫁接幼苗粗根生物量为10.55~32.64 g,中根生物量在0.57~3.38 g,细根生物量为1.32~4.40 g。EY9总生物量最大(199.41 g),是无性系XH6总生物量最小值的3.7倍。可见,四川桤木不同无性系嫁接幼苗生物量存在较大差异。
2.3 无性系嫁接幼苗各器官生物量分配特征
经测定发现(图5),无性系JT5生物量分配表现为主干>叶>侧枝>粗根>中根>细根,无性系XH2、XH7、XH10、XH14、XH5、EY5、EY6和EY10生物量分配表现为叶>主干>侧枝>粗根>细根>中根,其他无性系则表现为主干>叶>侧枝>粗根>细根>中根。其中EY10叶的生物量占比最大(36.05%),XH4叶生物量占比最小(31.40%),XH3侧枝生物量占比最小(5.96%),EY2主干生物量占比最大(41.04%),而EY6则具有最大侧枝生物量占比(8.50%)和最小的主干生物量占比(32.27%)。

图5 四川桤木不同无性系各器官生物量分配比Fig.5 Allocation ratio of biomass in different A.ledgeriana clones
经测定发现(图6),无性系XH11根冠比最大为0.37,EY2根冠比最小为0.24。可见,四川桤木不同无性系嫁接幼苗生物量分配存在差异。

图6 四川桤木不同无性系根冠比Fig.6 Root-shoot ratio of different A.ledgeriana clones
2.4 不同无性系嫁接幼苗聚类分析
系统聚类分析表明,在平均欧式距离约13处,可将34个四川桤木无性系嫁接幼苗按照生物量划分为3类(图7)。
由图7可以看出,第1类包含15个无性系(XH1、XH4、XH5、XH9、XH12、XH13、EY2、EY3、EY7、EY8、JT1、JT2、JT4、JT5、JT7),第2类包含7个无性系(XH8、XH16、EY4、EY9、EY11、JT3、JT6),第3类包含12个无性系(XH2、XH3、XH6、XH7、XH10、XH11、XH14、XH15、EY1、EY5、EY6、EY10)。其中,第2类无性系植株总生物量和各器官生物量均最大,第1类无性系生物量次之,第3类无性系生物量最小。

图7 四川桤木不同无性系生物量聚类分析Fig.7 Cluster analysis of A.ledgeriana clones based on biomass
经测算可知(表1),第2类无性系与第1、3类相比较,分配给侧枝和细根的生物量最多;第1类无性系植株与第2、3类比较,侧枝和主干生物量占比较大,而叶片和细根生物量占比最小;第3类无性系植株与第1、2类相比较,叶片生物量占比最大,侧枝和主干生物量占比最小,粗根生物量占比最大但是其生物量明显小于第1、2类。可见,3类四川桤木无性系嫁接幼苗生物量及其分配特征存在明显差异。

表1 四川桤木不同无性系生物量终聚类中心分布Table 1 Final cluster center distribution of biomass in different A.ledgeriana clones
3 讨 论
3.1 嫁接幼苗生物量分配特征
植物生物量是植物物质和能量积累的基本体现[17],生物量的大小表明植物利用和积累空间资源的能力[18],而植物器官水分含量的高低则表明其抗旱能力的强弱[19]。本研究中,四川桤木嫁接幼苗叶片和主干相对含水量均在50%以上,侧枝相对含水量为49%~69%,表明四川桤木嫁接幼苗抗旱能力较强,对环境具有一定的适应能力。
植物同化产物可在代谢源-库之间运输,且同化运输具有优先供应生长点的特点[20]。本研究中,四川桤木嫁接幼苗主干和叶片分配的生物量较高,而侧枝分配的生物量最低。这表明主干可能是1年生四川桤木嫁接幼苗地上部分主要的代谢库,而叶片不仅是主要的代谢源还是重要的代谢库,主干和叶片是同化产物优先供应的生长点。
3.2 嫁接幼苗生物量分配特点及其环境适应能力
生物量分配可以反映植物对环境的适应策略和能力[21-22]。通过对四川桤木无性系嫁接幼苗生物量及其分配聚类分析表明,34个四川桤木无性系嫁接幼苗按照生物量被划分为3类。四川桤木不同无性系嫁接幼苗栽培于相同的环境条件中,并给予相同的管理措施,但是不同无性系嫁接幼苗的长势不同,这可能主要是因为四川桤木不同无性系间遗传性状存在差异,从而导致其生物量分配特征不同。这与李鑫等[23]对不同地理种源云南松幼苗生物量分配及其异速生长的研究结果相似。
与第1、3类无性系相比,第2类无性系嫁接幼苗的总生物量最大,嫁接幼苗分配给侧枝和细根的生物量最大。植物对土壤水分和矿质营养元素的吸收主要是通过细根(根毛区)完成的,细根分配的生物量较多说明根系对养分的吸收更强,根系可以吸收更多的水分、矿质营养元素等供地上部分生长所需,这潜在地增加了植物适应土壤水分和养分变化的能力[24-25]。不仅如此,根系对矿质元素的吸收能力还会影响同化产物的运输能力。因此,通常细根生物量越大,植物体代谢源-库之间物质运输越快,植物适应环境变化(如倒春寒、季节性干旱)的能力越强[26-27]。而本试验地位于少日照地区(年日照时间<1 300 h),侧枝的生物量分配较高说明幼苗冠层截获光合辐射的能力越强[20],这潜在地增加了植物适应少日照环境的能力。可见,第2类无性系嫁接幼苗累计的生物量最大,且生物量分配较均匀,可以很好地适应当地土壤水热条件、光照条件等。
此外,第1类无性系总生物量次于第2类无性系,侧枝和主干生物量占比较大,叶片和细根生物量占比最小,说明虽然第1类无性系同化产物主要运输到侧枝和主干,幼苗接收光能的面积较大,但是由于细根生物量较少,根系吸收养分的能力较差,这表明第1类无性系适应当地少日照的能力较强,但是对土壤水热条件的适应能力不强。这与舒文将等[28]对棱角山矾幼苗研究结果相似。
根冠比可以反映植株地上部分及地下部分生长关系及植物自身对环境的适应能力[29]。一般情况下,植物在受到干旱胁迫[30-31]和盐胁迫[32]时会增加根冠比,以提高其对环境的适应能力。本研究中四川桤木无性系根冠比存在一定的差异,第3类无性系根冠比大于第1类和第2类无性系,这表明第3类四川桤木无性系嫁接幼苗可能受到了干旱胁迫或盐胁迫。进一步分析发现,第3类无性系的叶片生物量占比最大,主干、侧枝和细根生物量占比较低,粗根和中根生物量占比最大但是其生物量明显小于第1、2类。第3类无性系嫁接幼苗根系生物量主要分配给了粗根和中根,而细根生物量及其分配比例均较低。说明第3类无性系幼苗虽然根系分配的生物量较大,但是其根系活性较低,嫁接幼苗只是保留着砧木原有的根系,但是没有及时生长出新根(细根)吸收土壤水分和矿质养分[33],幼苗虽然生长出了完整的叶片,可以进行光合作用,但是光合作用积累的同化产物不多,同化产物只能就近运输(胞内或胞间运输)到光合叶片,满足光合细胞自身所需,而长距离运输到侧枝、主干、细根的能力较差[34],对光照条件和土壤水热条件的适应能力可能最差。
此次研究了四川桤木不同无性系1年生嫁接幼苗的生物量及分配特征。为了进一步了解不同无性系四川桤木嫁接幼苗的环境适应策略和能力,今后应整合不同无性系四川桤木嫁接幼苗生长、光合、生理特征等指标,综合评价其环境适应策略和能力。
猜你喜欢
浙江农业科学(2024年2期)2024-02-27 15:03:24
中国书法(2023年9期)2023-12-26 18:03:59
水土保持研究(2020年2期)2020-03-19 09:03:26
乡村科技(2019年21期)2019-02-21 09:19:29
生态学报(2018年17期)2018-10-18 02:26:48
四川农业大学学报(2018年3期)2018-09-04 08:38:36
农村农业农民·B版(2018年2期)2018-03-22 08:47:22
农村.农业.农民(2018年4期)2018-01-23 02:07:28
中南林业科技大学学报(2016年1期)2016-12-20 08:09:31
广西林业科学(2016年3期)2016-03-16 05:43:36
