
黄淮平原农业景观不同生境鸟类多样性特征及年际动态
2021-04-07 03:19洪咏怡卢训令赵海鹏
生态学报 2021年5期
洪咏怡,卢训令,3,*,赵海鹏
1 河南大学黄河中下游数字地理技术教育部重点实验室, 开封 475004 2 河南大学环境与规划学院, 开封 475004 3 河南大学河南省大气污染综合防治与生态安全重点实验室, 开封 475004 4 河南大学生命科学学院, 开封 475004
生物多样性是人类赖以生存的最重要的物质来源,决定着区域生态系统功能与服务的形成和维持[1- 6],鸟类是生物多样性的重要组成部分,并在生态系统功能和服务方面有着重要作用[7- 11],同时对环境变化又非常敏感。当生态环境发生改变时,鸟类对其做出的反应较为迅速并易于观测,所以鸟类是生态环境变化的重要指示物种[12]。鸟类多样性变化是其栖息地质量的重要指标,鸟类多样性及其种群数量变化可以直接反映其栖息地服务功能的实际情况[13]。
在全球人口不断增加,对粮食的需求日益增长的情况下,农业用地持续扩张,自然、半自然景观下降,农业景观中非农生境日益减少,异质性降低,并进一步加剧了生物多样性的持续快速丧失[14- 16],包括鸟类多样性[17- 18]。目前国内有关鸟类的研究多集中于自然保护区、湿地、城市等区域,对农业景观中的鸟类研究相对较少[19- 20],尤其缺乏长时间连续观测研究和数据。对农业景观中不同生境间的鸟类多样性组成特征进行探究,有助于区域鸟类生物多样性形成机制的揭示,并可为区域鸟类多样性保护及区域生态环境持续发展提供理论参考和建议。区域生物多样性(γ多样性)可分解为α和β多样性,通过对生物多样性的拆分,有助于从局地和景观等不同尺度上揭示生物多样性的组成机制,因此引起了研究者更多的重视,并取得了大量的成果[21- 24]。另外,世界上超过50%的鸟类以昆虫为食,近75%的鸟类至少偶尔吃无脊椎动物[25],因此食虫鸟类在农业景观中提供了重要的虫害控制服务,而这关系着农业生产的持续发展与粮食安全[10- 11,26]。本研究以黄淮平原农业景观作为研究区,选取湖泊、河流、农田、村庄等4种主要生境类别,对其进行连续4年的鸟类多样性调查,分析以下问题:不同生境间的鸟类多样性差异如何?各生境在年际间又呈现怎样的变化趋势?并结合鸟类食性对研究区鸟类所提供的虫害控制服务进行探究。同时借助物种多样性拆分分析,探讨区域鸟类多样性的形成机制,旨在为区域鸟类多样性保护提供理论基础和科学建议。
1 研究区概况
选取河南省开封市杞县(34°13′—34°46′N,114°35′—114°56′E)和周口市淮阳县(33°24′—33°55′N,114°38′—115°5′E)作为研究区。该研究区位于黄淮平原,地形起伏较小,为典型农业区,属温带大陆性季风气候(图1)。

图1 研究区示意图Fig.1 Sketch map of the research area
2 研究方法
2.1 数据获取
根据研究区土地利用现状选取典型地段布置样线,共设置20条,杞县和淮阳县各10条,每条样线长约2.5 km,包含湖泊、河流、农田、村庄4种生境(图1)。2016年至2019年4年间每年于鸟类繁殖期(5—6月)分两次进行调查,选择晴朗无大风的天气,于每日5:00—9:30,16:00—18:30两个鸟类活动高峰时间段进行。调查时至少两人一组,向前行进速度大约为2—3 km/h,记录沿前进方向样线两侧宽度大约50 m内观测到的鸟类,只记录迎面鸟,从背后飞入的不作记录。在调查中以肉眼观测为主,同时辅以望远镜、照相机进行辨别及记录,以备后期对有疑义的物种进一步确认[19]。鸟类区系分布、食性、迁徙等资料主要依据《世界鸟类分类与分布名录》和《中国鸟类志(上下)》等资料[27- 28]。
2.2 数据处理与分析
2.2.1α与β多样性指数
本研究选取常规的α多样性指数分别有Shannon-Wiener多样性指数、Pielou均匀度指数、Margalef丰富度指数以及Simpson多样性指数[29]。β多样性测算选取Cody指数计算物种年际替代速率,选取Sørensen和Jaccard等相似性指数,以计算生境间鸟类群落组成的相似程度[30]。
2.2.2物种的丰富度加性的分配
物种丰富度加性分配能够在不同尺度上计算物种丰富度对各生境的贡献以及不同样线间、不同生境间物种多样性的差异,并说明物种多样性组成特征,且度量单位一致,生态学意义明显,更易于为区域生物多样性保护提供科学依据[23,31- 32]。
物种丰富度加性分配公式(S):
S=α+β1+β2
在计算各生境物种的加性分配时,α表示该生境内各样线的平均物种丰富度,β1为该生境不同样线间的物种多样性差异,S为该生境总物种丰富度;进行区域物种总丰富度计算时,α和β1的含义同前,β2表示生境间物种丰富度的差异,S为区域总物种丰富度。
本文的数据整理和α多样性、β多样性计算均在Microsoft Excel 2007中进行。
3 结果分析
3.1 物种组成总体特征
研究区内连续4年的鸟类多样性调查,共观察记录到22922只个体,分属14目,38科,53属,87种。调查中共发现国家二级保护动物两种:红脚隼(Falcoamurensis)22只,红隼(Falcotinnunculus)7只,均是在农田或河流生境中观察到。4年数据进行对比发现:从2016年至2019年,记录到的鸟类个体数在年际间呈先减少后增加趋势,其中2019年个体数最多,为8036只;2017年最少,仅4117只。麻雀(Passermontanus)物种多度最高,共计7815只;其次是家燕(Hirundorustica),共观察到4504只。从区系分布来看,各生境中均以广布种占绝对优势,还有一部分为东洋种,古北种数量最少;从居留型来看,留鸟是本研究区鸟类群落的最主要组成部分,占总个体数的70.72%,尤其在村庄与农田生境中更为突出,在河流与湖泊生境中也有一定量的旅鸟出现;从生态类群来看,各生境中均以鸣禽为主导,游禽与涉禽仅在河流与湖泊等生境中被观察到(图2)。

图2 黄淮平原农业景观中鸟类物种组成基本特征Fig.2 Characteristics of bird species composition in agricultural landscape of Huanghuai Plain
3.2 食性组成特征
研究区鸟类食性组成显示(表1),在研究区调查记录到的87种鸟类中,有食虫鸟类57种,约占总物种数的77%,植食性和杂食性鸟类各11种和9种,分别占12.64%和10.34%;从个体数目数据看,杂食性鸟类形成了区内农业景观中鸟类的主体,占比超过了52%,其次是食虫鸟类,约占40%,植食性鸟类个体较少,仅占7.30%。从不同生境类别中鸟类个体数来看(图3),农田和村庄中均以杂食性鸟类占优势,占比超过61%;而在湖泊和村庄生境中,则以食虫鸟类占优,分别占56%和47%;而植食性鸟类在各生境中数量均较少。
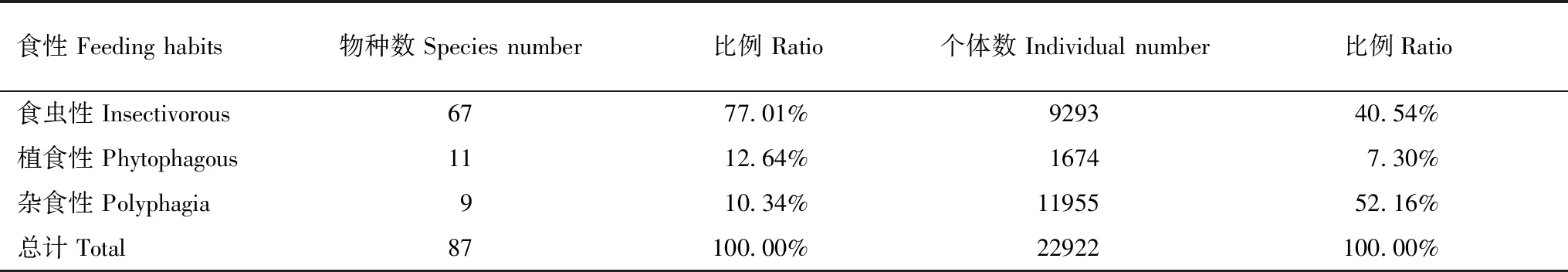
表1 黄淮平原农业景观中鸟类食性组成特征
鸟类食性组成的年际变化显示(图4),在物种丰富度方面,从2016年至2019年,食虫鸟类物种比例逐年上升,而其他两类总体上有轻微的下降;从个体数目来看,食虫鸟类比例亦呈轻微上升态势,植食性鸟类波动不大,杂食性鸟类个体数所占比例下降趋势明显。
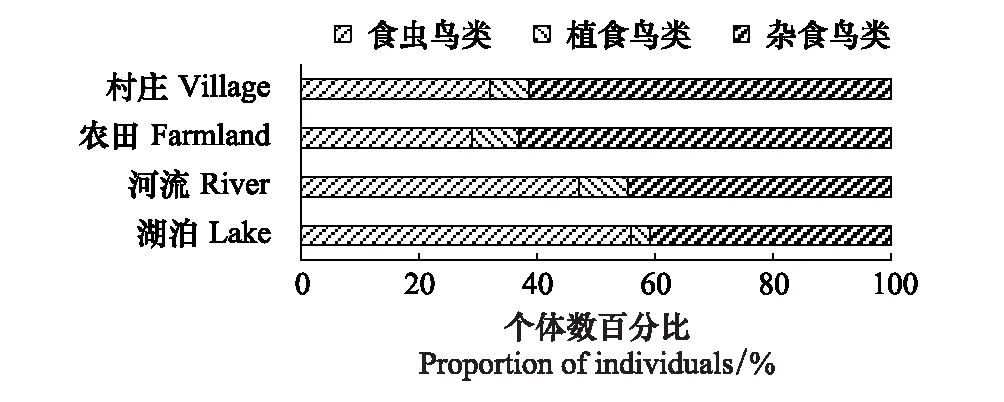
图3 黄淮平原农业景观不同生境鸟类食性组成特征 Fig.3 Characteristics of bird feeding habits in different habitats of agricultural landscape in Huanghuai Plain

图4 黄淮平原农业景观鸟类食性组成年际变化Fig.4 Interannual variation of bird feeding composition in agricultural landscape of huanghuai plain
3.3 α多样性及其年际变化特征
对黄淮平原农业景观不同生境中鸟类多样性特征分析发现(图5),湖泊生境中的Margalef丰富度指数在2017年明显下降,而后逐年上升,2016年的数值为4年中的最高值;河流和农田生境中呈先上升后下降的趋势,最高值均出现在2018年,最低值出现在2016年;村庄中的丰富度指数变化趋势表现为先下降后上升,随后又有小幅度的下降。河流和农田中的Shannon指数和Simpson指数历年变化趋势比较接近。河流生境中的两种多样性指数均呈先增后减的趋势,其中Shannon指数最高值出现在2018年,Simpson多样性最高值则出现在2017年;农田中的Shannon指数先增加而后减少,在2019年又有极小幅度的增加,Simpson指数在2017年最高,呈先增后减趋势。湖泊和村庄生境的变化趋势相反,在湖泊中呈现先下降后上升的趋势,最低值出现在2018年;在村庄中则先上升后下降,2018年出现最高值。河流、农田、村庄生境中的Pielou均匀度指数变化趋势一致,均为先上升后下降,且最高值出现在2017年。

图5 黄淮平原农业景观鸟类群落α多样性特征及其年际变化Fig.5 α diversity characteristics and interannual variation of bird communities in agricultural landscape of huanghuai plain图中误差线为标准误
3.4 生境间群落相似性及年际变化特征
3.4.1群落相似性分析
通过群落相似性分析发现,各生境鸟类群落间的Sørensen和Jaccard相似性系数变化趋势基本一致(表2)。农田和村庄的相似性最高,其次是河流和农田、湖泊和农田以及湖泊和村庄。河流和湖泊、河流和村庄的相似性最低。研究区所有群落间的Jaccard相似性指数均超过50%,根据Jaccard相似性原理,所有群落间均为中等相似(0.50 表2 黄淮平原生境间鸟类群落相似性 3.4.2群落相似性年际变化特征 β多样性指沿不同环境梯度变化,不同群落之间物种组成的差异性。β多样性越高,表明群落之间共有种越少[33]。以时间为梯度,分析黄淮平原各生境鸟类群落的年际变化,结果(图6)表明农田和村庄生境中,相邻年份之间的Cody多样性指数(βc)随时间推移呈上升趋势,意味着物种替代速率加快,年际间的物种丰富度波动幅度上升。湖泊生境中的Cody指数在2017—2018年有所下降,而后上升;在河流生境中,Cody指数仅在2018—2019有略微下降。 图6 黄淮平原农业景观不同生境中鸟类群落2016—2019年Cody多样性指数的年际变化Fig.6 Interannual change of Cody diversity index in different habitats of agricultural landscape in Huanghuai Plain in 2016—2019 图中误差线表示标准误 研究区物种丰富度加性分配结果(图7)显示:从各生境来看,样线尺度上α多样性对物种丰富度的贡献均较高,全在50%以上,从高到低依次为湖泊(31种,64.58%)、村庄(28.8种,62.50%)、农田(28.6种,58.37%)、河流(37.1种,52.29%);从全区来看,α多样性贡献了总物种丰富度的23.06%。在各生境中,样线间的物种多样性差异(β1)的贡献率均比α多样性低,说明各生境内不同样线之间物种差异较低;在区域水平上,β1多样性贡献了全区总物种丰富度的38.86%,余下的38.08%的物种丰富度来自于生境间的物种多样性(β2)的贡献。 图7 研究区各生境物种丰富度加性分配结果Fig.7 Additive distribution results of species richness in each habitat of the study area 图中误差线表示标准误;图上*前的数据为物种丰富度,()中数据为该水平物种丰富度在总丰富度中的占比 从总体来看,河流和湖泊生境的物种多样性更明显,推测水体的存在对提高物种多样性有显著作用,因为这将为鸟类提供一种全新或更多样化资源,尤其是为部分生境专性鸟类所利用,如涉禽和游禽。农田和村庄的α多样性相对而言较低,这可能与农业景观中的异质性有关。在农业景观中环境异质性对生物多样性的提高和保护往往有着积极作用[34]。促进农业景观中不同类型植被尤其是农作物和非农作物的配置、减少大规模种植几种作物为主的景观,有利于提高农田中的物种多样性[35- 36]。 农业景观中的鸟类提供了重要的生态系统功能和服务,尤其是食虫鸟类所提供的天敌控制服务意义重大。在本调查中发现的鸟类中,超过77%的物种和40%的个体均为为食虫鸟类,这些鸟类将为区域内的虫害控制提供重要服务,即使是杂食性的鸟类,在繁殖期大多也需要取食昆虫来获得更多的能量进行繁殖。对于以昆虫为主食的鸟类而言,在繁殖期则需要取食更多的昆虫甚至无脊椎动物。调查发现研究区2017年鸟类个体总数为4年中最少,推测鸟类在此期间可能没有获得更多的食物资源以度过繁殖期。食肉的猛禽在食物链中属于顶级消费者,影响着世界各地各种生态系统中各种啮齿动物和其他小型哺乳动物的种群动态[37]。许多猛禽物种栖息在农业生态系统中,对啮齿动物、小型哺乳动物种群的数量有一定的控制作用。此外,猛禽提供的生态系统服务还包括控制病虫害以及疾病的传播、减少啮齿动物等取食农作物、清除动植物残体以及促进营养循环等等。猛禽的数量和分布受生境变化的影响并不强烈,而主要受到食物丰富度的影响[38]。调查共发现红隼、红脚隼共计29只,其中2016年在农田生境中发现20只,2016年之后猛禽遇见概率大幅下降,推测此处可供猛禽捕食的资源有所减少,又或者是猛禽原有的生态位受到其他生物挤压。 另一方面,鸟类还可能会对农作物产生一定的危害,但鸟类造成的经济损失并没有得到很好的量化,并且认为农田中存在着鸟类就意味着农作物损失是来源于鸟类的观点往往是有失偏颇的[39- 40]。与农田间的啮齿动物以及昆虫相比,鸟类造成的农作物损失可能相当低[41]。 Margalef丰富度指数除农田呈逐年稳步增长态势之外,其余生境均有更复杂的波动;从4年总体水平来看,河流在拥有相对较高的丰富度的同时也包含了最多的个体数;湖泊包含的个体数最少,但丰富度仅次于河流。河流、湖泊的Shannon指数相对而言历年波动较大,而农田、村庄的变化则相对平缓。河流和湖泊的Simpson指数变化趋势几乎相反,而农田和村庄则较为相近。除去湖泊,其余三种生境的Pielou均匀度指数历年变化趋势一致。从食性组成来看,近年来区域内农业景观中食虫鸟类的种类和个体数比例均呈一定的上升趋势,杂食性鸟类则有所下降。 造成鸟类多样性年际波动的原因复杂多样。从栖息地本身来看,可能是食物资源种类和数量增加或减少、受到的人为干扰程度改变,或是植被物种丰富度等受到了影响[42]。宏观来看,也可能与全球气候波动等有关。本研究受数据时长所限,仅能综合各方面因素做些许猜想,当前的年际波动是否属于正常范围?以及年际间是否存在一个正常波动的阈值?超出正常波动的阈值将会产生何种影响抑或是预警?都是值得日后深入思考的问题。 生物多样性的持续快速丧失是当前人类面临最严重的环境问题[43- 44],生物多样性保护也成为生态学研究中的核心内容之一,为加强生物多样性保护,人类提出了大量的生物多样性保护计划,但生物多样性仍在持续下降中,各保护措施的有效性也有一定的争论[45- 46]。农业景观中的生物多样性发挥着重要的生态系统功能[8- 9,47- 48]。区域生物多样性形成机制及其影响因素的研究是有效保护措施提出的前提,前人对物种多样性的研究多关注的是不同尺度上的编目多样性,而对β多样性的研究相对较弱,但通过对生物多样性的拆分,会有助于对生物物种群落结构、动态与机制的理解,并进而有助于更有针对性保护措施的提出与施行[23]。研究区农业景观中鸟类多样拆分发现,尽管各生境类别中α多样性具有重要作用,但β多样性在不同尺度上对总物种丰富度具有重要作用,从区域尺度看,生境异质性的提高对鸟类多样性的增加也意义重大,未来区域农业景观中鸟类多样性的保护需要加强两方面的工作:一是重视区域生境多样性的提高,因为任一新生境的出现均可能促进某种鸟类的生存与扩散[19,49];另外还要关注景观尺度上异质性的作用,景观异质性将通过β多样性的变化作用于区域生物多样性,而已有大量研究证实了这种现象[19,23,50- 52]。 良好的生态环境能够为鸟类繁殖提供稳定场所,维持区域生物多样性。物种多样性拆分结果表明α多样性和β多样性对区域鸟类多样性的形成均意义重大,应当多尺度联合采取保护措施,提高生境异质性的同时也要注重小尺度环境质量的改善。 许多鸟类生态系统的服务和功能都是通过消耗资源来产生的[53]。掌握鸟类的食性不仅是鸟类保护工作的重要一环,也是保护被取食动植物的前提。对于破坏农作物的鸟类,了解它们的觅食行为可以用来制定更有效的减轻损失的策略[54]。河流和湖泊对水鸟而言,是为必要的停歇地、觅食地和栖息地。因此,应当重点关注河流和湖泊生境质量,完善污水和垃圾处理,改善现有水域环境质量,并提高水域周边植被覆盖率。农田和村庄中的林下荫蔽环境是伴人鸟类的栖身之所,农田边缘的动植物、人类废弃的食物等都能够为鸟类提供充足的食物资源,通过多样化作物种植与非农生境的留存将有助于鸟类多样性的保护,可探讨土地共享模式的有效性[55- 56]。但总体上农业景观中鸟类多样性的保护依然任重而道远[20]。 致谢:感谢河南黄河湿地国家级自然保护区马朝红工程师,河南本地鸟类爱好者徐奎、姚鹏、韩红波、陈志磊、宋剑南、雷中广、黄锋、韩志明和赵鑫等,河南大学李振、张露西、孙冬旭、王子豪、崔璨、张慧、高书文、韩子威、李思涵、李晶晶、李理想、于中斌、胡家宁、魏俊俊、史华杰和罗丽莎等同学在野外调查和数据录入中提供帮助。 参考文献(References): [1] Tilman D, Isbell F, Cowles J M. Biodiversity and ecosystem functioning. Annual Review of Ecology, Evolution, and Systematics, 2014, 45: 471- 493. [2] Wood S A, Karp D S, DeClerck F, Kremen C, Naeem S, Palm C A. Functional traits in agriculture: agrobiodiversity and ecosystem services. Trends in Ecology & Evolution, 2015, 30(9): 531- 539. [3] Isbell F, Cowles J, Dee L E, Loreau M, Reich P B, Gonzalez A, Hector A, Schmid B. Quantifying effects of biodiversity on ecosystem functioning across times and places. Ecology Letters, 2018, 21(6): 763- 778. [4] Veen G F, Van Der Putten W H, Bezemer T M. Biodiversity-ecosystem functioning relationships in a long-term non-weeded field experiment. Ecology, 2018, 99(8): 1836- 1846. [5] Van Der Plas F. Biodiversity and ecosystem functioning in naturally assembled communities. Biological Reviews, 2019, 94(4): 1220- 1245. [6] Mace G M, Norris K, Fitter A H. Biodiversity and ecosystem services: a multilayered relationship. Trends in Ecology & Evolution, 2012, 27(1): 19- 26. [7] Bommarco R, Kleijn D, Potts S G. Ecological intensification: harnessing ecosystem services for food security. Trends in Ecology & Evolution, 2013, 28(4): 230- 238. [10] Karp D S, Mendenhall C D, Sandi R F, Chaumont N, Ehrlich P R, Hadly E A, Daily G C. Forest bolsters bird abundance, pest control and coffee yield. Ecology Letters, 2013, 16(11): 1339- 1347. [11] Maas B, Karp D S, Bumrungsri S, Darras K, Gonthier D, Huang J C C, Lindell C A, Maine J J, Mestre L, Michel N L, Morrison E B, Perfecto I, Philpott S M,Ç H, Silva R M, Taylor P J, Tscharntke T, Van Bael S A, Whelan C J, Williams-Guillén K. Bird and bat predation services in tropical forests and agroforestry landscapes. Biological Reviews, 2016, 91(4): 1081- 1101. [12] Ikin K, Yong D L, Lindenmayer D B. Effectiveness of woodland birds as taxonomic surrogates in conservation planning for biodiversity on farms. Biological Conservation, 2016, 204: 411- 416. [13] 于长水, 常青, 王涛, 莫日根, 魏敬铤, 潘艳秋. 乌梁素海生态健康与鸟类服务功能调查. 北方环境, 2013, 29(3): 10- 13. [14] FAO. The State of the World′s Biodiversity for Food and Agriculture. Rome: FAO, 2019: 572- 572. [15] Kubiszewski I, Costanza R, Anderson S, Sutton P. The future value of ecosystem services: Global scenarios and national implications. Ecosystem Services, 2017, 26: 289- 301. [16] Newbold T, Hudson L N, Hill S L L, Contu S, Lysenko I, Senior R A, Börger L, Bennett D J, Choimes A, Collen B, Day J, De Palma A, Díaz S, Echeverria-Londoo S, Edgar M J, Feldman A, Garon M, Harrison M L K, Alhusseini T, Ingram D J, Itescu Y, Kattge J, Kemp V, Kirkpatrick L, Kleyer M, Correia D L P, Martin C D, Meiri S, Novosolov M, Pan Y, Phillips H R P, Purves D W, Robinson A, Simpson J, Tuck S L, Weiher E, White H J, Ewers R M, Mace G M, Scharlemann J P W, Purvis A. Global effects of land use on local terrestrial biodiversity. Nature, 2015, 520(7545): 45- 50. [17] Stanton R L, Morrissey C A, Clark R G. Analysis of trends and agricultural drivers of farmland bird declines in North America: a review. Agriculture, Ecosystems & Environment, 2018, 254: 244- 254. [18] Robinson R A, Sutherland W J. Post-war changes in arable farming and biodiversity in Great Britain. Journal of Applied Ecology, 2002, 39(1): 157- 176. [19] 卢训令, 赵海鹏, 孙金标, 杨光. 黄淮平原农业景观鸟类多样性及生境差异. 生态学报, 2019, 39(9): 3133- 3143. [20] Li L, Hu R C, Huang J K, Bürgi M, Zhu Z Y, Zhong J, Lü Z. A farmland biodiversity strategy is needed for China. Nature Ecology & Evolution, 2020, 4(6): 772- 774. [21] Legendre P, De Cáceres M. Beta diversity as the variance of community data: dissimilarity coefficients and partitioning. Ecology Letters, 2013, 16(8): 951- 963. [22] López-Martínez J O, Hernández-Stefanoni J L, Dupuy J M, Meave J A. Partitioning the variation of woody plant β-diversity in a landscape of secondary tropical dry forests across spatial scales. Journal of Vegetation Science, 2013, 24(1): 33- 45. [23] 卢训令, 汤茜, 梁国付, 丁圣彦. 黄河下游平原非农植物多样性拆分研究. 生态学报, 2016, 36(14): 4395- 4405. [24] Baselga A. Partitioning abundance‐based multiple‐site dissimilarity into components: balanced variation in abundance and abundance gradients. Methods in Ecology and Evolution, 2017, 8(7): 799- 808. [25] Wenny D G, Devault T L, Johnson M D, Kelly D, Sekercioglu C H, Tomback D F, Whelan C J. The need to quantify ecosystem services provided by birds. The Auk, 2011, 128(1): 1- 14. [26] Grass I, Lehmann K, Thies C, Tscharntke T. Insectivorous birds disrupt biological control of cereal aphids. Ecology, 2017, 98(6): 1583- 1590. [27] 赵正阶. 中国鸟类志(上下). 长春: 吉林科学技术出版社, 2001. [28] 郑光美. 世界鸟类分类与分布名录. 北京: 科学出版社, 2002. [29] 刘玉明, 马克平. 生物群落多样性的测度方法: Ⅰα多样性的测度方法(下). 生物多样性, 1994, 2(4): 231- 239. [30] 梅象信, 徐正会, 张继玲, 赵宇翔. 昆明西山森林公园东坡蚂蚁物种多样性研究. 林业科学研究, 2006, 19(2): 170- 176. [31] Crist T O, Veech J A. Additive partitioning of rarefaction curves and species-area relationships: unifying α‐, β‐ and γ‐diversity with sample size and habitat area. Ecology Letters, 2006, 9(8): 923- 932. [32] 陈圣宾, 欧阳志云, 徐卫华, 肖燚. Beta多样性研究进展. 生物多样性, 2010, 18(4): 323- 335. [33] 王世雄, 王孝安, 郭华. 黄土高原植物群落演替过程中的β多样性变化. 生态学杂志, 2013, 32(5): 1135- 1140. [34] Lee M B, Goodale E. Crop heterogeneity and non-crop vegetation can enhance avian diversity in a tropical agricultural landscape in southern China. Agriculture, Ecosystems & Environment, 2018, 265: 254- 263. [35] Pithon J A, Beaujouan V, Daniel H, Pain G, Vallet J. Are vineyards important habitats for birds at local or landscape scales? Basic and Applied Ecology, 2016, 17(3): 240- 251. [36] Assandri G, Bogliani G, Pedrini P, Brambilla M. Diversity in the monotony? Habitat traits and management practices shape avian communities in intensive vineyards. Agriculture, Ecosystems & Environment, 2016, 223: 250- 260. [37] Korpimäki E, Krebs C J. Predation and population cycles of small mammals: a reassessment of the predation hypothesis. BioScience, 1996, 46(10): 754- 764. [38] 冯理, 闵龙, 韩联宪, 赵健林, 刘学先, 松建华. 云南纳帕海自然保护区越冬猛禽考察初报. 四川动物, 2008, 27(3): 445- 448. [39] Greene C D, Nielsen C K, Woolf A, Delahunt K S, Nawrot J R. Wild turkeys cause little damage to row crops in Illinois. Transactions of the Illinois State Academy of Science, 2010, 103(3/4): 145- 152. [40] Gebhardt K, Anderson A M, Kirkpatrick K N, Shwiff S A. A review and synthesis of bird and rodent damage estimates to select California crops. Crop Protection, 2011, 30(9): 1109- 1116. [41] Basili G D, Temple S A. Dickcissels and crop damage in Venezuela: defining the problem with ecological models. Ecological Applications, 1999, 9(2): 732- 739. [42] 胡君梅, 丁志锋, 王玲, 杨道德, 胡慧建. 广州城市绿地鸟类物种多样性的时空变化. 野生动物学报, 2017, 38(1): 44- 51. [43] Ceballos G, Ehrlich P R, Barnosky A D, García A, Pringle R M, Palmer T M. Accelerated modern human-induced species losses: entering the sixth mass extinction. Science Advances, 2015, 1(5): e1400253. [44] Payne J L, Bush A M, Heim N A, Knope M L, McCauley D J. Ecological selectivity of the emerging mass extinction in the oceans. Science, 2016, 353(6305): 1284- 1286. [45] Butchart S H M, Walpole M, Collen B, Van Strien A, Scharlemann J P W, Almond R E A, Baillie J E M, Bomhard B, Brown C, Bruno J, Carpenter K E, Carr G M, Chanson J, Chenery A M, Csirke J, Davidson N C, Dentener F, Foster M, Galli A, Galloway J N, Genovesi P, Gregory R D, Hockings M, Kapos V, Lamarque J F, Leverington F, Loh J, McGeoch M A, McRae L, Minasyan A, Morcillo M H, Oldfield T E E, Pauly D, Quader S, Revenga C, Sauer J R, Skolnik B, Spear D, Stanwell-Smith D, Stuart S N, Symes A, Tierney M, Tyrrell T D, Vié J C, Watson R. Global biodiversity: indicators of recent declines. Science, 2010, 328(5982): 1164- 1168. [46] Cardinale B. Overlooked local biodiversity loss. Science, 2014, 344(6188): 1098- 1098. [47] Isbell F, Adler P R, Eisenhauer N, Fornara D, Kimmel K, Kremen C, Letourneau D K, Liebman M, Polley H W, Quijas S, Scherer-Lorenzen M. Benefits of increasing plant diversity in sustainable agroecosystems. Journal of Ecology, 2017, 105(4): 871- 879. [49] Jeliazkov A, Mimet A, Chargé R, Jiguet F, Devictor V, Chiron F. Impacts of agricultural intensification on bird communities: New insights from a multi-level and multi-facet approach of biodiversity. Agriculture, Ecosystems & Environment, 2016, 216: 9- 22. [50] Tscharntke T, Klein A M, Kruess A, Steffan-Dewenter I, Thies C. Landscape perspectives on agricultural intensification and biodiversity-ecosystem service management. Ecology Letters, 2005, 8(8): 857- 874. [51] Tscharntke T, Tylianakis J M, Rand T A, Didham R K, Fahrig L, Batáry P, Bengtsson J, Clough Y, Crist T O, Dormann C F, Ewers R M, Frund J, Holt R D, Holzschuh A, Klein A M, Kleijn D, Kremen C, Landis D A, Laurance W, Lindenmayer D, Scherber C, Sodhi N, Steffan-Dewenter I, Thies C, Van Der Putten W H, Westphal C. Landscape moderation of biodiversity patterns and processes-eight hypotheses. Biological Reviews, 2012, 87(3): 661- 685. [52] Angelella G M, Holland J D, Kaplan I. Landscape composition is more important than local management for crop virus-insect vector interactions. Agriculture, Ecosystems & Environment, 2016, 233: 253- 261. [53] Whelan C J, Wenny D G, Marquis R J. Ecosystem services provided by birds. Annals of the New York Academy of Sciences, 2008, 1134(1): 25- 60. [54] Herrmann E, Anderson M D. Foraging behaviour of damage-causing birds in table grape vineyards in the Orange River valley, South Africa. South African Journal of Enology and Viticulture, 2007, 28(2): 150- 154. [55] Baudron F, Schultner J, Duriaux J Y, Gergel S E, Sunderland T. Agriculturally productive yet biodiverse: human benefits and conservation values along a forest-agriculture gradient in southern Ethiopia. Landscape Ecology, 2019, 34(2): 341- 356. [56] Green R E, Cornell S J, Scharlemann J P W, Balmford A. Farming and the fate of wild nature. Science, 2005, 307(5709): 550- 555.

3.5 物种加性分配
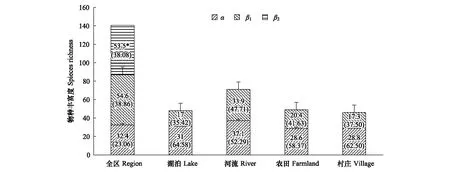
4 讨论
4.1 不同生境鸟类多样性及其生态系统服务效应
4.2 不同生境鸟类多样性年际动态
4.3 农业景观鸟类多样性构成和保护
5 建议

猜你喜欢
大气科学学报(2022年2期)2022-05-12
华北水利水电大学学报(自然科学版)(2021年4期)2021-10-08
空间科学学报(2021年2期)2021-07-21
落叶果树(2021年6期)2021-02-12
现代装饰(2020年10期)2020-10-28
中国海洋大学学报(自然科学版)(2019年9期)2019-01-04
现代园艺(2018年1期)2018-03-15
小学阅读指南·低年级版(2017年2期)2017-03-23
体育科技(2016年2期)2016-02-28
浙江大学学报(农业与生命科学版)(2015年4期)2015-12-15
