
福建三种常绿阔叶林碳氮磷生态化学计量特征
2021-04-07 03:19黄雍容黄石德谭芳林游惠明
生态学报 2021年5期
黄雍容,高 伟,*,黄石德,林 捷,谭芳林,游惠明,杨 丽
1 福建省林业科学研究院,福州 350012 2 福建省森林培育与林产品加工利用重点实验室,福州 350012 3 福建省三明市尤溪县林业局,三明 365100
碳(C)、氮(N)、磷(P)是陆地生态系统植物生长所必需的大量元素,参与调节植物生长的各种生理过程[1],其中,C是植物体主要组成元素,N、P则是植物生长的主要限制养分[2]。研究表明植物中C、N、P组成及分配联系紧密,并与环境因子共同决定了植物的生长发育及营养水平[3]。生态化学计量学结合了生态学和化学计量学,是当下研究生态系统中养分元素平衡的热门学科[4]。目前,生态化学计量学被广泛应用于森林生态学(从个体、种群、群落、生态系统直至区域)研究之中,尤其在揭示植物养分循环规律及限制性养分元素、种群结构、生态系统动态平衡、全球气候变化的响应等方面应用较多[5- 10]。近年来,大量研究开始关注驱动植物生态化学计量格局的主要因子,并将影响植物叶片养分含量的光照、温度、水分、土壤养分等环境因子归纳为气候因子、地理条件和土壤性质等三大因素[11- 13]。
在区域尺度上,与气候因子和土壤性质相比,地理条件对植物营养状态的影响更为复杂[14- 16]。近年来研究表明,大尺度上,随纬度升高和温度降低植物叶片的C、N、P含量显著增加而N∶P却减少[17- 19]。姜沛沛等[20]在陕西省的研究表明,植物叶片N、P随温度和降水的增加而增加,随纬度增加而降低,凋落物N含量随年均温和年降水量的增加而增加,P含量和N∶P随年均温和年降水量的增加而增加。王晶苑等[8]研究认为植物叶片的N∶P与气温呈负相关,而叶片C、N、P化学计量与降水量无显著相关性。李路等[21]研究发现降水量和温度是影响土壤C、N、P含量及空间分布的主要因子,表层土壤C∶N随温度和年降水量的增大而增大,湿热地区生产力高,土壤C、N含量较高,淋溶导致表层土壤P流失。可见年均温和降水量会影响植物叶片的C、N、P含量,但影响程度因地理位置和研究对象不同差异较大。常绿阔叶林是亚热带湿润地区特有的地带性森林植被类型,其树种组成多样、群落结构复杂、物种多样性高,在亚热带森林生态系统中具有极其重要的地位和作用。福建省地处东南沿海,纵跨南亚热带和中亚热带两个区域,分布了多个国家级自然保护区,保护了大面积的天然常绿阔叶林,为开展森林生态化学计量研究,阐明亚热带森林生态系统的养分供应状况及其耦合关系,揭示影响森林结构与功能恢复的限制性因子等提供了天然的实验场所。
本研究沿中亚热带至南亚热带依次选择福建省3个国家级保护区内典型常绿阔叶林为研究对象,分别为武夷山保护区的甜槠(Castanopsiseyrei)林,天宝岩保护区的刨花润楠(Machiluspauhoi)林和虎伯寮保护区的薄叶润楠(Machilusleptophylla)林,3片常绿阔叶林分别位于福建省3大山脉(武夷山脉、戴云山脉和博平岭山脉),地理环境造成的局部小气候差异,可能影响不同地区植物种类及生长速率,进而影响生态系统的养分循环。基于此,本研究对福建省3个典型常绿阔叶林内植物、凋落物和土壤的C、N和P含量及其生态化学计量进行研究,拟解决以下问题:(1)福建常绿阔叶林“植物-凋落物-土壤”C、N、P化学计量特征及其限制营养元素;(2)“植物-凋落物-土壤”C、N、P养分的耦合关系;(3)植物、凋落物C、N、P化学计量特征与主要非生物因子的相关关系。
1 材料与方法
1.1 试验地概况
本研究自2016年开始,沿福建中亚热带至南亚热带依次在武夷山、天宝岩和虎伯寮保护区内典型林分中设置样地(图1),面积均为100 m×100 m。对样地生物多样性进行全面调查,并对样地内乔木挂牌编号,各样地自然概况见表1。在样地调查的基础上,分别选择典型植被区设置3个20 m×20 m样方,于2017年10 —11月选择晴朗天气进行植物和土壤样品采集。
1.2 样品的采集和测定
每个样地内选取不同层次(乔木层、灌木层和草本层)重要值前50%的物种进行采样,其中乔木层选取平均木各3—6株,采集东西南北4个方位林冠中部成熟叶片,混匀后采用四分法取样;灌木层和草本层为整株采样。共采集200个植物样品,分属于18科23属32种,其中乔木14种,灌木17种,草本7种。每个20 m×20 m的样方分成4个10 m×10 m的小样方,每个小样方内按梅花点法设置5个1 m×1 m小样方收集地表新鲜凋落物,混匀后分别装入样品袋,每个调查地各12个凋落物样品,共36个。上述样品带回实验室内65℃烘干至恒重粉碎过100目筛备用,测定植株碳氮磷含量。
沿样方对角线每5 m用土钻取0—10 cm土壤样品,每条对角线取5点混成1个样品,共采集土壤样品9个。将样品带回室内风干后去除根系、石砾等杂质,研磨过100目筛,测定土壤碳氮磷含量。采用元素分析仪(EURO Vector EA 3000, Italy)测定全碳和全氮含量。采用比色法测定全磷含量,方法为称取0.30 g样品,植物加入5 mL HNO3+1 mL H2O2,土壤加入10 mL HNO3,于微波消解仪中充分消解,过滤定容后置于全自动间断化学分析仪(Smartchem 200, France)中测定。

图1 样点分布图Fig.1 Distribution of sampling sites
1.3 气象数据
于中国国家气象科学数据共享服务平台(data.cma.cn)下载与固定样地相近的武夷山气象站(海拔222.1 m),大田气象站(海拔400.1 m)和南靖气象站(海拔22.8 m)的累年月平均气温(分别为18.3,19.1和21.2℃)和累年月平均降水量(1926,1552和1798 mm),根据气象站与样地的海拔差异,推算采样区的年均温。
1.4 数据处理及分析

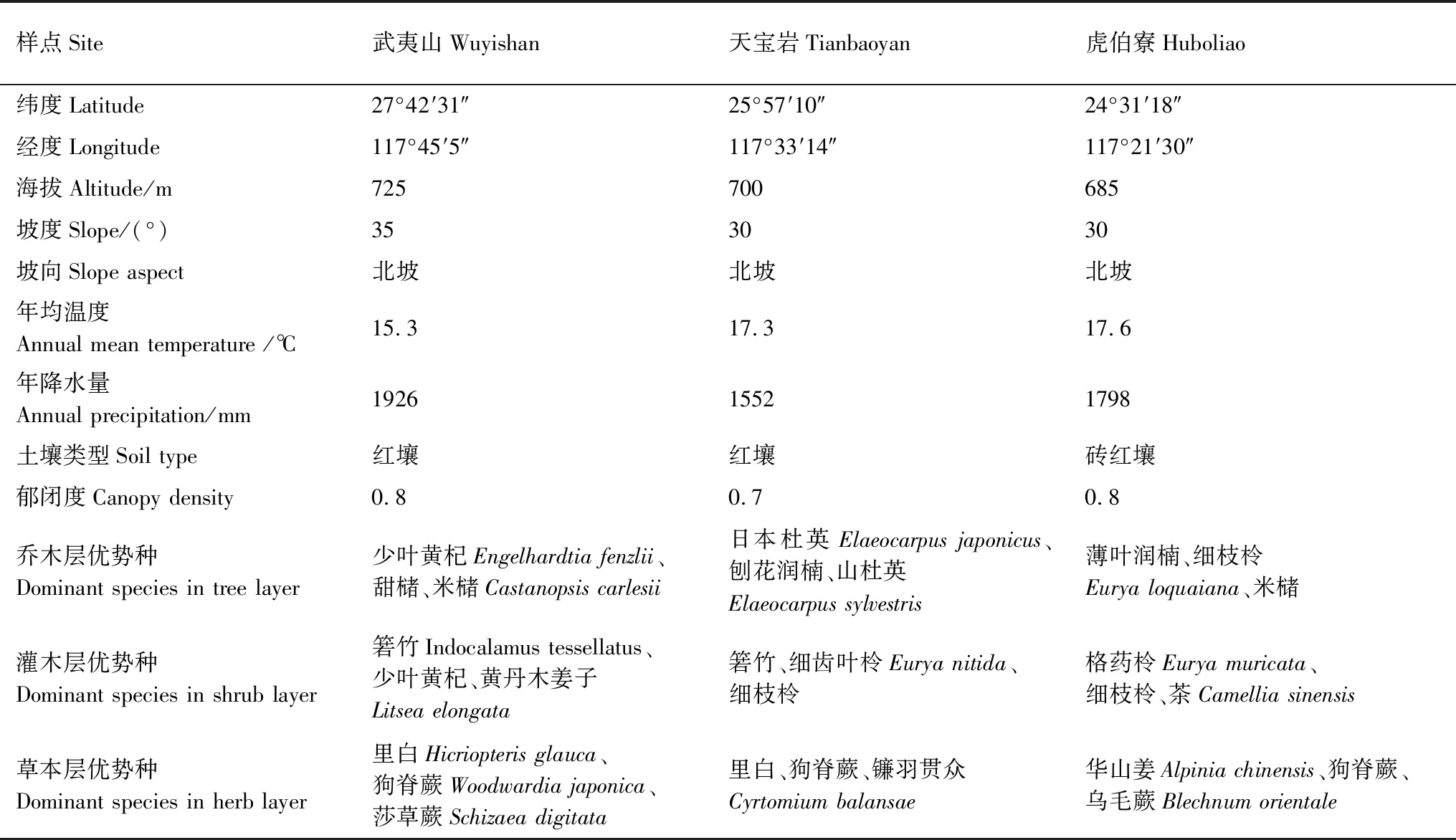
表1 试验地概况
2 结果与分析
2.1 植物、凋落物和土壤C、N和P含量
由表2可知,3个样点乔木层和灌木层C含量均显著高于其他组分。武夷山和天宝岩乔木层N含量显著高于其他组分,虎伯寮各组分中草本层N含量最高,但各植物层之间N含量无显著差异。P含量比较上,各样点间差异较大,武夷山、天宝岩和虎伯寮P含量最高的组分分别为灌木层、乔木层和草本层。除乔木层N含量外,虎伯寮植物和凋落物层C、N和P含量均显著高于天宝岩和武夷山。武夷山表层土壤C和N含量显著高于虎伯寮和天宝岩,3个样点表层土壤P含量表现出显著差异,排序为天宝岩>武夷山>虎伯寮。

表2 不同样点常绿阔叶林各组分碳、氮和磷含量特征
2.2 植物、凋落物和土壤C、N和P化学计量特征
由表3可知,武夷山植物C∶N显著高于天宝岩和虎伯寮,灌木层和草本层C∶N变化均随纬度的升高而增大,表现为武夷山>天宝岩>虎伯寮,各个样点凋落物层C∶N无显著差异,虎伯寮表层土壤C∶N显著高于天宝岩和武夷山。草本层、乔木层以及凋落物层C∶P表现为武夷山>天宝岩>虎伯寮,天宝岩灌木层C∶P显著高于武夷山和天宝岩,其表层土壤则显著低于其他样点。虎伯寮草本层、乔木层以及凋落物层N∶P显著低于武夷山和天宝岩,天宝岩灌木层N∶P显著高于武夷山和虎伯寮,其土壤N∶P显著低于另外2个样点。

表3 不同样点常绿阔叶林各组分C、N和P含量化学计量特征
2.3 植物、凋落物、土壤C、N、P含量及化学计量特征的相关性
由表4可知,叶片C、N、P含量彼此间呈极显著正相关;叶片N与凋落物P、叶片C、P与凋落物C、N、P含量彼此间呈极显著正相关。叶片N、P与土壤养分含量呈极显著负相关;叶C与土壤P呈极显著负相关。凋落物N与土壤N含量呈显著负相关(P<0.05),与土壤P含量呈极显著负相关。土壤N与土壤C呈极显著正相关。
叶的C∶N与叶C∶P、凋落物C∶P和N∶P呈极显著正相关,与叶N∶P、表层土壤C∶N呈极显著负相关;叶C∶P与叶N∶P、凋落物C∶P和N∶P呈极显著正相关,与土壤C∶N呈极显著负相关;叶N∶P与凋落物C∶P和N∶P均呈极显著正相关,与土壤C∶N、C∶P和N∶P呈极显著负相关关系。凋落物C∶N与C∶P呈极显著正相关,与土壤C∶N呈显著负相关;凋落物C∶P与N∶P呈极显著正相关,与土壤C∶N呈极显著负相关,与土壤C∶P呈显著负相关;凋落物N∶P与土壤C∶N和C∶P呈极显著负相关。土壤养分化学计量比彼此间呈极显著正相关。

表4 植物-凋落物-土壤C、N和P含量及化学计量比相关系数
除叶N含量与N∶P呈极显著正相关外、与土壤C∶P关系不显著外,叶N、P含量与叶C∶N、C∶P和N∶P呈极显著负相关;叶N、C、P含量与凋落物C∶P和N∶P均呈极显著负相关、与土壤C∶N和C∶P均呈极显著正相关。凋落物N和C含量分别与叶N∶P和C∶P、P含量与叶C∶N、C∶P和N∶P均呈极显著负相关;除凋落物的N含量与凋落物N∶P关系不显著外,凋落物N、P含量与凋落物C∶N、C∶P和N∶P呈极显著负相关,凋落物C含量与C∶N和N∶P分别呈极显著正相关和负相关;除凋落物N和P含量与土壤N∶P关系不显著外,其N、C和P含量与土壤C∶N、C∶P和N∶P均呈显著正相关。除土壤N含量与土壤C∶N呈极显著负相关外,土壤N和C含量与叶片和土壤C∶N、C∶P、与凋落物C∶N、C∶P和N∶P均呈显著正相关;土壤P含量与叶片和凋落物C∶P、N∶P呈极显著正相关、与土壤C∶N、C∶P和N∶P均呈显著负相关。
2.4 植物、凋落物与主要非生物因子的冗余分析
由图2可见,植物和凋落物化学计量特征在RDA第Ⅰ轴和第Ⅱ轴解释量分别为83.33%和13.96%,累积解释量达97.29%,能准确地反映叶片、凋落物化学计量特征与非生物因子的关系。
沿RDA第Ⅰ轴,大气温度与叶C、N、P以及凋落物P含量之间夹角小于90°,表明温度与叶C、N、 P以及凋落物P含量呈正相关,其中温度的投影较长,表明温度对其具有较大的正面影响。相反,温度与叶和凋落物的C∶N、C∶P和N∶P之间的夹角大于90°,表明温度与叶和凋落物的化学计量比呈负相关,且温度的投影较长,表明温度对其具有较大的负面影响。故温度对叶和凋落物的养分含量呈正相关的影响,而对其化学计量比呈负相关的影响。沿RDA第Ⅱ轴,降水量与叶和凋落物的C含量、C∶N之间夹角小于90°,表明降水量与叶和凋落物的C含量、C∶N之间呈正相关;其中降水量的投影较长,表明叶和凋落物的C含量、C∶N受降水量正面影响显著;相反,降水量与叶片N含量、N∶P的夹角远大于90°,表明降水量与两指标之间呈负相关。
RDA分析表明,环境因子对叶片和凋落物化学计量影响程度重要性排序为大气温度>土壤C∶N>土壤N>土壤P>降水量,5个因子解释量分别为25.3%、6.3%、1.3%、2.5%和5.1%,均达极显著水平(P<0.01),是影响植物叶片、凋落物化学计量特征的主要因子。
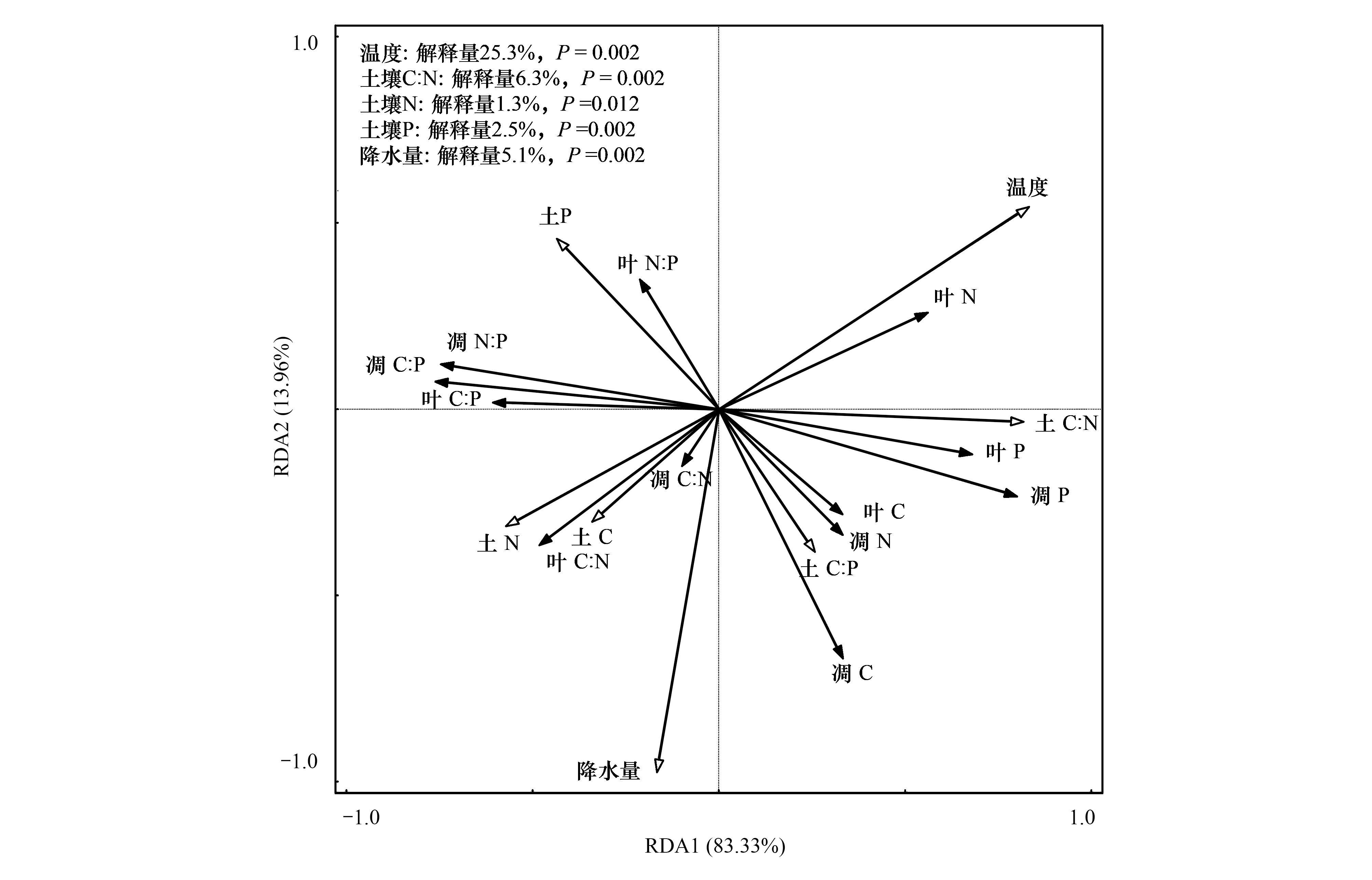
图2 植物、凋落物C、N、P及化学计量比与主要环境因子的RDA分析Fig.2 Redundancy analysis of plant and litter C, N, P content and stoichiometric ratios with main environmental factors
3 讨论
3.1 福建常绿阔叶林碳氮磷化学计量特征
植物生长由大量元素C、N、P互作调节决定,美国海洋学家Redfield提出了“Redfield 比值”[22],近年来生态学家们对陆地生态系统的研究中,发现陆地生态系统各层次的C、N、P也存在既定的比例关系。与中国东部南北样带森林生态系统优势植物叶片C(374.1—646.5 g/kg)、N(8.4—30.5 g/kg)和P(0.6—6.2 g/kg)含量相比[23],本研究中,福建常绿阔叶林群落中植物叶C(459.3 g/kg)、N(16.7 g/kg)、P(0.7 g/kg)平均含量均在其范围内,其中P含量接近最低值。Han等[5]对中国753种陆生植物N和P化学计量进行研究,结果表明753种植物N和P几何平均含量分别为18.6和1.21 g/kg,本研究结果中叶片N平均含量接近该研究结果,P含量则远低于该研究。与王晶苑等[8]对亚热带常绿阔叶林优势植物CNP化学计量比研究结果(C、N、P含量分别为472.8,19.8和1.54 g/kg)相比,本研究乔木层叶C含量与其无显著差异,N含量较低,P含量显著低于该研究。可见,福建常绿阔叶林的植物处于高C低P的元素格局。
植物叶片N∶P比值用来表征植物受N、P养分的限制格局[24,25]。Koersdman等[26]通过施肥试验提出,N∶P比大于16时,该生态系统受P的限制,当N∶P比小于14时,则认为是受N的限制,若N∶P比在14—16范围内,被认为是受到两种元素共同限制或两种元素都不缺少[27]。本研究中叶片C∶N(30.6)和C∶P(791)均显著高于全球尺度的值(C∶N=23.8和C∶P=300.9)[28]和王晶苑[8]对亚热带常绿阔叶林的研究结果(C∶N=23.89,C∶P=457.98);C∶N与中国东部南北样地森林生态系统102种优势种叶片的C∶N(29.1)接近,C∶P则显著高于任书杰[23]的研究结果313.9,这主要跟P含量较低有关。本研究中植物叶的平均N∶P为26.4,大于16,并且远大于全球尺度的研究结果(13.8)[28]、中国区域性的研究结果(18.0)[5]、中国东部南北样带的研究结果(11.5)[23]以及王晶苑等[8]对亚热带常绿阔叶林的研究结果(19.5),可见福建常绿阔叶林生长主要受P元素限制。
森林土壤中的养分主要来源于凋落物的分解,凋落物的分解速率决定养分的释放量[29]。本研究中凋落物的C、N、P平均含量分别为457.4,14.3和0.5 g/kg,与王晶苑等[8]研究结果(C、N、P含量分别为514.12、14.22和0.43 g/kg)相比较,本研究结果C含量较低,N和P含量与其相接近。凋落物的分解依赖于微生物[30],当凋落物中的N、P含量充足时,微生物分解力越强,越有利于分解[31]。有研究表明,凋落物中养分的固持和释放存在既定的临界值,当C∶N低于40[32],C∶P低于600[33],凋落物中N或P含量超过微生物生长所需部分才会释放[29]。本研究3个样点的凋落物C∶N分别为31.8,31.5和32.7,低于C∶N释放临界值,而C∶P分别为746.7,1222.7和1348.4,远大于C∶P释放临界值,可见福建常绿阔叶林凋落物中N元素含量相对充足利于释放,而P元素相对缺乏,容易更多的被微生物固持,从而导致土壤中磷养分的不足。
土壤养分是植物生长所需养分的直接来源,其含量受凋落物分解养分归还的影响[34]。土壤C∶N,C∶P和N∶P是土壤有机质组成和养分有效性的重要指标[35]。本研究3个样点表层土壤C∶N,C∶P和N∶P平均值分别14.8,286.9和22.1,远大于Tian等[36]的研究结果(11.9,61和5.2),体现了土壤养分含量空间上较大的变异性[37]。与王晶苑[8]的研究结果(13.1,113.1和8.6)相比,本研究土壤C∶N比与其接近,C∶P和N∶P则大于其结果,再次说明土壤中P元素的缺乏,是影响福建常绿阔叶林养分循环的重要因素。
3.2 植物、凋落物和土壤碳氮磷化学计量特征的耦合关系
大量研究表明,植物、凋落物和土壤C、N、P含量及其化学计量联系紧密,形成了森林生态系统养分循环的内在调控机制[38-40]。本研究中,亚热带森林植物生长受P元素限制,叶片N、P元素含量与土壤对应元素含量呈显著负相关,与Garnier[41]的研究结果不一致,他认为若植物生长受某元素限制,其叶片相应元素含量会与土壤对该养分的供应能力呈正相关[20]。本研究对象为自然保护区常绿阔叶林,植物组成多样,群落结构复杂,不适于用具体物种的研究结果来解释。叶片与凋落物养分含量间呈良好的正相关性,与姜沛沛等[20]研究结果一致,但凋落物与土壤养分呈负相关性,说明该林分土壤养分的消耗已经超过了凋落物的养分供给能力,由此导致了生态系统养分循环缓慢。此外,叶片与土壤之间N、P含量呈负相关性,说明土壤N、P元素供应严重不足,叶片中的N、P经凋落物分解回归土壤,实际上凋落物中的N、P元素无法及时补充到土壤中,需进一步对凋落物分解过程中N、P元素的分配及去向进行深入研究。凋落物的分解与林分中植物种类、土壤有效养分含量高低[42]、土壤酶活性、土壤微生物种类及含量等环境因子有关[29],什么原因导致的凋落物养分回归障碍有待进一步研究。
3.3 福建常绿阔叶林养分分布格局的非生物驱动因子
冗余分析表明,大气温度和土壤C∶N对植物叶片和凋落物养分含量的影响较大,解释量分别为25.3%和6.3%。土壤C∶N与叶和凋落物P含量之间呈显著正相关。姜沛沛等[20]将植物叶片N含量随温度的升高而升高的现象归因于树种的广生态位和较大的纬度变化幅度,表明在较小区域尺度上,植物叶片化学计量特征与环境因子的关系存在较大的变异性。本研究结果表明,叶片N、P含量与大气温度呈显著正相关,而土壤N含量与大气温度呈显著负相关,与姜沛沛等[20]、马玉珠等[43]的研究结果相似,说明温度升高,有利于植物对土壤中N、P养分的吸收。另外,本研究集中于福建省区域范围内,研究对象为常绿阔叶林,物种多样性高、优势树种生态位广,也可能是导致该结果的另一原因。另一研究结果叶和凋落物的C∶N、C∶P和N∶P与大气温度呈显著负相关,而Ge等[44]对中国主要阔叶树凋落叶碳氮磷化学计量特征与气候因子的关系研究结果表明凋落物C∶P和N∶P随纬度(温度和降水量降低)的升高而降低,该文中所涉及的树种大部分为北方树种,Ge等[44]认为森林的植物组成是影响凋落物碳氮磷化学计量比的关键因子,本研究对象为中亚热带常绿阔叶林的植物组成丰富、生活型复杂,可能是导致群落水平叶片和凋落物碳氮磷化学计量比与气温呈负相关的原因。叶和凋落物C∶N、C∶P和N∶P与土壤C∶N呈显著负相关,当土壤中N含量越高,叶和凋落物养分利用效率越低。沈芳芳等[42]研究表明氮沉降会显著提高土壤中有效氮的含量且存在累积效应,而氮沉降加剧了亚热带森林生态系统P的限制,当土壤中N含量超过一定阈值,会抑制凋落物分解。RDA分析还表明,降水量与叶和凋落物的C∶N具有显著正相关,较高的降水量,一方面激发微生物活性加速了凋落物分解,另一方面土壤淋溶加重,进而影响叶片和凋落物的养分含量及化学计量比[21]。
4 结论
福建常绿阔叶林C、N和P含量在各层次变化趋势,表现为植物>凋落物层>土壤层,同一层次的养分含量在各林分间有较大差异。对3个样点各层次C、N和P化学计量比分析结果表明,P元素是常绿阔叶林植物生长最主要的限制因子,土壤中P元素的缺乏,是影响福建常绿阔叶林养分循环的重要因素。土壤养分消耗超过凋落物养分供给能力,是导致该森林生态系统养分循环缓慢的重要原因。大气温度和土壤C∶N是影响亚热带森林生态系统养分循环的关键环境因子。
致谢:感谢武夷山国家公园管理局金昌善、天宝岩国家级自然保护区刘进山、虎伯寮国家级自然保护区吴卫江及福建省林业科学研究院尤龙辉工程师等对本研究的支持。
参考文献(References):
[1] Reich P B, Tjoelker M G, Machado J L, Oleksyn J. Universal scaling of respiratory metabolism, size and nitrogen in plants. Nature, 2006, 439(7075): 457- 461.
[2] 吴晓燕. 龙福建长汀马尾松及其林下植物叶片化学计量与水分利用效率特征[D]. 福州: 福建师范大学, 2014.
[3] Güsewell S. N∶P ratios in terrestrial plants: variation and functional significance. New Phytologist, 2004, 164(2): 243- 266.
[4] Elser J J, Dobberfuhl D R, MacKay N A, Schampel J H. Organism size, life history, and N∶P stoichiometry: toward a unified view of cellular and ecosystem processes. Bioscience, 1996, 46(9): 674- 684.
[5] Han W X, Fang J Y, Guo D L, Zhang Y. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytologist, 2005, 168(2): 377- 385.
[6] 任书杰, 于贵瑞, 陶波, 王绍强. 中国东部南北样带654种植物叶片氮和磷的化学计量学特征研究. 环境科学, 2007, 28(12): 2665- 2673.
[7] 孙阳, 王晶媛, 张慧, 王建楠, 吴统贵, 张鹏. 海岸梯度上黑松针叶化学计量特征的变化规律. 生态学杂志, 2019, 38(6): 1662- 1668.
[8] 王晶苑, 王绍强, 李纫兰, 闫俊华, 沙丽清, 韩士杰. 中国四种森林类型主要优势植物的C∶N∶P化学计量学特征. 植物生态学报, 2011, 35(6): 587- 595.
[9] 刘兴诏, 周国逸, 张德强, 刘世忠, 褚国伟, 闫俊华. 南亚热带森林不同演替阶段植物与土壤中N、P的化学计量特征. 植物生态学报, 2010, 34(1): 64- 71.
[10] 黄小波, 刘万德, 苏建荣, 李帅锋, 郎学东. 云南普洱季风常绿阔叶林152种木本植物叶片C、N、P化学计量特征. 生态学杂志, 2016, 35(3): 567- 575.
[11] 冯秋红. 南北样带温带区栎属树种功能性状及其与环境因子的关系[D]. 北京: 中国林业科学研究院, 2008.
[12] Kang H Z, Zhuang H L, Wu L L, Liu Q L, Shen G R, Berg B, Man R Z, Liu C J. Variation in leaf nitrogen and phosphorus stoichiometry inPiceaabiesacross Europe: an analysis based on local observations. Forest Ecology and Management, 2011, 261(2): 195- 202.
[13] Chen Y H, Han W X, Tang L Y, Tang Z Y, Fang J Y. Leaf nitrogen and phosphorus concentrations of woody plants differ in responses to climate, soil and plant growth form. Ecography, 2013, 36(2): 178- 184.
[14] Wu T G, Dong Y, Yu M K, Wang G G, Zeng D H. Leaf nitrogen and phosphorus stoichiometry ofQuercusspecies across China. Forest Ecology and Management, 2012, 284: 116- 123.
[15] Wu T G, Wang G G, Wu Q T, Cheng X R, Yu M K, Wang W, Yu X B. Patterns of leaf nitrogen and phosphorus stoichiometry amongQuercusacutissimaprovenances across China. Ecological Complexity, 2014, 17: 32- 39.
[16] Wu T G, Yu M K, Wang G G, Dong Y, Cheng X R. Leaf nitrogen and phosphorus stoichiometry across forty-two woody species in southeast China. Biochemical Systematics and Ecology, 2012, 44: 255- 263.
[17] 任书杰, 于贵瑞, 陶波, 官丽莉, 方华军, 姜春明. 兴安落叶松(LarixgmeliniiRupr.)叶片养分的空间分布格局. 生态学报, 2009, 29(4): 1899- 1906.
[18] Reich P B, Oleksyn J. Global patterns of plant leaf N and P in relation to temperature and latitude. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(30): 11001- 11006.
[19] Hedin O L. Global organization of terrestrial plant-nutrient interactions. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(30): 10849- 10850.
[20] 姜沛沛, 曹扬, 陈云明, 赵一娉. 陕西省3种主要树种叶片、凋落物和土壤N、P化学计量特征. 生态学报, 2017, 37(2): 443- 454.
[21] 李路, 常亚鹏, 许仲林. 天山雪岭云杉林土壤CNP化学计量特征随水热梯度的变化. 生态学报, 2018, 38(22): 8139- 8148.
[22] Redfield A C. The biological control of chemical factors in the environment. American Scientist, 1958, 46(3): 205- 221.
[23] 任书杰, 于贵瑞, 姜春明, 方华军, 孙晓敏. 中国东部南北样带森林生态系统102个优势种叶片碳氮磷化学计量学统计特征. 应用生态学报, 2012, 23(3): 581- 586.
[24] He J S, Fang J Y, Wang Z H, Guo D L, Flynn D F B, Geng Z. Stoichiometry and large-scale patterns of leaf carbon and nitrogen in the grassland biomes of China. Oecologia, 2006, 149(1): 115- 122.
[25] He J S, Wang L, Flynn D F B, Wang X P, Ma W H, Fang J Y. Leaf nitrogen: phosphorus stoichiometry across chinese grassland biomes. Oecologia, 2008, 155(2): 301- 310.
[26] Koerselman W, Meuleman A F M. The vegetation N∶P ratio: a new tool to detect the nature of nutrient limitation. Journal of Applied Ecology, 1996, 33(6): 1441- 1450.
[27] Wassen M J, Venterink H G M O, De Swart E O A M. Nutrient concentrations in mire vegetation as a measure of nutrient limitation in mire ecosystems. Journal of Vegetation Science, 1995, 6(1): 5- 16.
[28] Elser J J, Fagan W F, Denno R F, Dobberfuhl D R, Folarin A, Huberty A, Interlandi S, Kilham S S, McCauley E, Schulz K L, Siemann E H, Sterner R W. Nutritional constraints in terrestrial and freshwater food webs. Nature, 2000, 408(6812): 578- 580.
[29] 李明军, 喻理飞, 杜明凤, 黄宗胜, 石建华. 不同林龄杉木人工林植物-凋落叶-土壤C、N、P化学计量特征及互作关系. 生态学报, 2018, 38(21): 7772- 7781.
[30] 郭剑芬, 杨玉盛, 陈光水, 林鹏, 谢锦升. 森林凋落物分解研究进展. 林业科学, 2006, 42(4): 93- 100.
[31] 陆晓辉, 丁贵杰, 陆德辉. 人工调控措施下马尾松凋落叶化学质量变化及与分解速率的关系. 生态学报, 2017, 37(7): 2325- 2333.
[32] Parton W, Silver W L, Burke I C, Grassens L, Harmon M E, Currie W S, King J Y, Adair E C, Brandt L A, Hart S C, Fasth B. Global-Scale similarities in nitrogen release patterns during long-term decomposition. Science, 2007, 315(5810): 361- 364.
[33] 赵谷风, 蔡延(马奔), 罗媛媛, 李铭红, 于明坚. 青冈常绿阔叶林凋落物分解过程中营养元素动态. 生态学报, 2005, 26(10): 3286- 3295.
[34] 李志安, 邹碧, 丁永祯, 曹裕松. 森林凋落物分解重要影响因子及其研究进展. 生态学杂志, 2004, 23(6): 77- 83.
[35] 朱秋莲. 黄土丘陵区不同植被带立地条件对植物—枯落物—土壤生态化学计量特征的影响[D]. 咸阳: 西北农林科技大学, 2013.
[36] Tian H Q, Chen G S, Zhang C, Melillo J M, Hall C A S. Pattern and variation of C∶N∶P ratios in China′s soils: a synthesis of observational data. Biogeochemistry, 2010, 98(1/3): 139- 151.
[37] 蒋芳. 中亚热带落叶阔叶林和常绿阔叶林土壤有机碳、全氮空间异质性比较[D]. 长沙: 中南林业科技大学, 2016.
[38] Fan H B, Wu J P, Liu W F, Yuan Y H, Hu L, Cai Q K. Linkages of plant and soil C∶N∶P stoichiometry and their relationships to forest growth in subtropical plantations. Plant and Soil, 2015, 392(1/2): 127- 138.
[39] Cao Y, Chen Y M. Coupling of plant and soil C∶N∶P stoichiometry in black locust (Robiniapseudoacacia) plantations on the Loess Plateau, China. Trees, 2017, 31(5): 1559- 1570.
[40] Hobbie S E, Gough L. Foliar and soil nutrients in tundra on glacial landscapes of contrasting ages in northern Alaska. Oecologia, 2002, 131(3): 453- 462.
[41] Garnier E. Interspecific variation in plasticity of grasses in response to nitrogen supply//Garnier G P, ed. Population Biology of Grasses. Cambridge: Cambridge University Press, 1998: 155- 181.
[42] 沈芳芳, 吴建平, 樊后保, 郭晓敏, 雷学明, 沃奇东. 杉木人工林凋落物生态化学计量与土壤有效养分对长期模拟氮沉降的响应. 生态学报, 2018, 38(20): 7477- 7487.
[43] 马玉珠, 钟全林, 靳冰洁, 卢宏典, 郭炳桥, 郑媛, 李曼, 程栋梁. 中国植物细根碳、氮、磷化学计量学的空间变化及其影响因子. 植物生态学报, 2015, 39(2): 159- 166.
[44] Ge J L, Xie Z Q. Leaf litter carbon, nitrogen, and phosphorus stoichiometric patterns as related to climatic factors and leaf habits across chinese broad-leaved tree species. Plant Ecology, 2017, 218(9): 1063- 1076.
猜你喜欢
中国土壤与肥料(2022年4期)2022-06-14
天津医科大学学报(2021年1期)2021-12-05
落叶果树(2021年6期)2021-02-12
文苑(2020年12期)2020-04-13
绿色中国(2019年18期)2020-01-04
海峡姐妹(2019年1期)2019-03-23
农家科技下旬刊(2017年8期)2017-11-13
中国医学影像技术(2017年11期)2017-01-16
科技创新导报(2016年21期)2016-12-17
考试周刊(2016年63期)2016-08-15
