
半干旱黄土小流域不同植被类型植物与土壤生态化学计量特征
2021-04-07 03:31杨智姣李宗善张钦弟
生态学报 2021年5期
温 晨,杨智姣,杨 磊,李宗善,卫 伟,张钦弟,*
1 山西师范大学生命科学学院, 临汾 041004 2 中国科学院生态环境研究中心 城市与区域生态国家重点实验室, 北京 100085
生态化学计量学主要关注生态过程中营养元素间的相互作用与平衡,是评价生物地球化学循环和生态系统营养状况的重要方法[1- 2]。碳(C)、氮(N)、磷(P)元素是构成生命体的基本元素[1],在生物体的活动中起着至关重要的作用。N和P是限制植物生长的主要元素,它们影响植物的生产力[3]、光合速率[4]和其他生态系统功能[5]。土壤养分含量也是指示植物生长养分供应的关键指标[6]。采用生态化学计量学的原理和方法研究生态系统中C、N、P元素的变化与平衡,为理解生态系统中生物体与自然环境相互作用关系及养分循环过程提供了有效手段[7]。
陆地生态系统中植物和土壤是紧密联系,相互关联的[8]。例如,土壤中C、N、P含量可决定植被群落结构和生长状态[9],反之植被通过凋落物和根系分泌物[10]的输入来影响土壤养分变化。近年来,大量研究探讨了草原[11]、农田[12]和森林[13]陆地生态系统中C∶N∶P化学计量特征。例如,Jiang等[12]对鄱阳湖农田土壤化学计量特征研究发现,N是决定土壤C∶N和N∶P的主要因素,而土壤C∶P主要受P的影响。杨阔等[13]研究发现青藏高原植物叶片N、P含量具显著正相关关系,且N∶P是判断植物生长元素受限情况的关键因子。Song等[14]研究发现在植被演替过程中,土壤C、N含量总是同步增加,而P含量则无显著变化。这些研究都是针对植物叶片或土壤的生态化学计量特征进行的,而对土壤-植物系统中的生态化学计量特征的研究较少,主要集中于区域尺度上叶片与土壤[15-16]化学计量特征的研究,对于植物其他器官如茎、根的化学计量特征及其与土壤关系研究较为缺乏,需进一步阐明。
黄土高原为有效防治严重的水土流失,实施开展以自然恢复和人工恢复为主的大规模“退耕还林还草”工程[17]。经过二十多年的不断治理,黄土高原植被覆盖率、土壤质量得到大幅度提升,形成了以自然恢复植被和人工恢复植被为主的多种植被类型。为了解黄土高原植被恢复中植物与土壤的变化特征,前人对不同植被恢复类型下土壤理化性质[18]、植被特征[19]等进行系统研究,但关于不同植被恢复类型下植物各器官生态化学计量特征及与土壤生态化学计量特征关系的研究仍较为缺乏。因此,本研究以典型半干旱黄土小流域3种植被恢复方式下(天然荒草、自然恢复、人工恢复)的5种植被类型(长芒草草地、赖草草地、苜蓿草地、柠条灌丛、山杏林)为研究对象,通过对其优势种植物的叶、茎、根及土壤的生态化学计量特征进行研究,试图明晰:不同植被恢复类型下植物各器官C、N、P含量及化学计量特征变化有何差异;土壤C、N、P含量及化学计量特征在各植被恢复类型间有何不同;该黄土小流域植物与土壤C、N、P含量及生态化学计量特征存在怎样的相互关系。旨在对黄土高原生态系统植物与土壤间养分循环进一步了解,为黄土高原植被恢复类型选择及策略优化提供科学依据。
1 材料与方法
1.1 研究区概况及样地选择
研究区位于黄土高原西部定西龙滩流域(104°27′—104°32′ E,35°43′—35°46′ N),是典型黄土丘陵沟壑区,属暖温带半干旱气候,年平均温度为6.8℃,年平均降水量为386 mm,大部分降雨发生在7月到9月[20]。本研究区土壤类型主要为低肥力、含沙量高、结构疏松的黄绵土[21]。流域内主要有以长芒草(Stipabungeana)、赖草(Leymussecalinus)、苜蓿(Medicagosativa)、柠条(Caraganakorshinskii)、山杏(Armeniacasibirica)等为主的多种植被。
1.2 样地选择及野外调查
依据代表性和典型性原则,在研究区内选取长芒草草地、赖草草地、苜蓿草地、柠条灌丛及山杏林5种典型植被类型共31个样地进行研究。其中,长芒草草地为该研究区代表性植被,未经种植、耕种等人为干扰活动,即天然荒草地;赖草草地为农地休耕后自然恢复样地,代表自然恢复植被;苜蓿草地、柠条灌丛及山杏林均为人工引进植被样地,代表人工恢复植被。在样方内进行物种调查,乔木层随机设置4个10 m×10 m的样方,记录其树种、树高、胸径、冠幅等因子;灌木层随机设置4个5 m×5 m样方,记录每个样方内灌木的种类、丛数、基径、株高、盖度等因子;草本层随机设置4个1 m×1 m样方,调查草本的种类、株数、盖度等因子。样地基本特征见表1。

表1 样地基本情况
1.3 样品采集与测定
植物叶、茎、根和土壤样品均于2017年8月采集。在乔灌木样地中随机选择5株长势良好、大小基本一致的优势种采集其叶片、茎样品分别混匀装入密封袋。乔木根样品的采集是以5株采样标准木基干为中心,在冠幅范围内随机钻取一个0—60 cm的土芯,采用冲洗法获得根样品,灌木根样品采用四分之一圆法进行取样,以标准株基部为圆心,半径为0.5 m,在土壤剖面取0—60 cm的土芯,经冲洗获得根样品。在草本样地中随机设置3个0.5 m×0.5 m小样方,在小样方内随机取优势植物10株采集其叶、茎样品,随之移除样方内地上植被及凋落物对根进行取样,分别混匀装入自封袋。上述根的取样均采用内径为8 cm的根钻。土壤取样点与植物取样点相对应,采用土钻法在对应样方内对0—60 cm土层以20 cm为间隔分三层取样。将以上样品经烘干、研磨、过筛等步骤处理以备元素测定。所有样品C、N、P含量分别采用重铬酸钾外加热法、凯氏定氮法、钼锑抗比色法进行测定[22]。
1.4 数据分析
采用单因素方差分析法(one-way ANOVA)对不同植被类型下叶、茎、根以及土壤C、N、P含量及化学计量特征进行分析,在检验方差齐性时,若方差齐,则采用最小显著差异法(LSD)进行多重比较;若方差不齐,则采用Tamhane′s T2法进行多重比较。同时采用双因素方差分析法(two-way ANOVA)对植被类型和不同器官对植物C、N、P含量及化学计量特征的影响进行分析。数据统计分析及作图分别在SPSS 17.0与SigmaPlot 10.0中进行,其中土壤C、N、P含量及化学计量比的取值均为3个土层的均值。利用R 3.5.1软件对植物与土壤养分含量及化学计量特征进行Pearson相关分析。
2 结果与分析
2.1 不同植被类型下植物各器官C、N、P含量及化学计量特征
从表2可得,同一器官内,植被类型对植物C、N、P含量及化学计量比具显著影响(P<0.05);同一植被类型中,不同器官对植物C、N、P含量及化学计量比也具显著影响(P<0.05)。不同植被类型下各器官C、N、P含量均表现为:叶>茎>根,且叶、茎、根的C、N、P含量及化学计量特征在5种典型植被间具一定差异(图1)。各器官C含量在不同植被间表现为山杏最高,长芒草最低,N含量则为柠条显著高于其他植被(P<0.05),而P含量在各植被间无显著差异(P>0.05)。不同植被类型下叶、茎、根的C∶N表现为赖草显著高于其他4种植被(P<0.05),叶、茎、根的C∶P在各植被间表现为柠条显著最低(P<0.05),但长芒草、赖草、苜蓿及山杏间无显著差异(P>0.05),N∶P则表现为柠条和苜蓿显著高于其他植被(P<0.05)。从总的恢复方式来看(表3),人工恢复植被各器官C、N含量及N∶P均显著高于天然荒草地(P<0.05)。P含量与C∶P在不同植被恢复类型间则无显著差异(P>0.05)。自然恢复植被各器官C∶N显著高于人工恢复植被与天然荒草地(P<0.05)。

表2 植被类型和器官对C、N、P含量及化学计量比的影响

图1 不同植被类型下植物各器官C、N、P含量及化学计量比Fig.1 Concentrations of C, N, P and stoichiometric ratio in plant organs under different vegetation types图中数据为平均值±标准差;不同大写字母代表同一器官不同植被类型间差异显著(P<0.05)
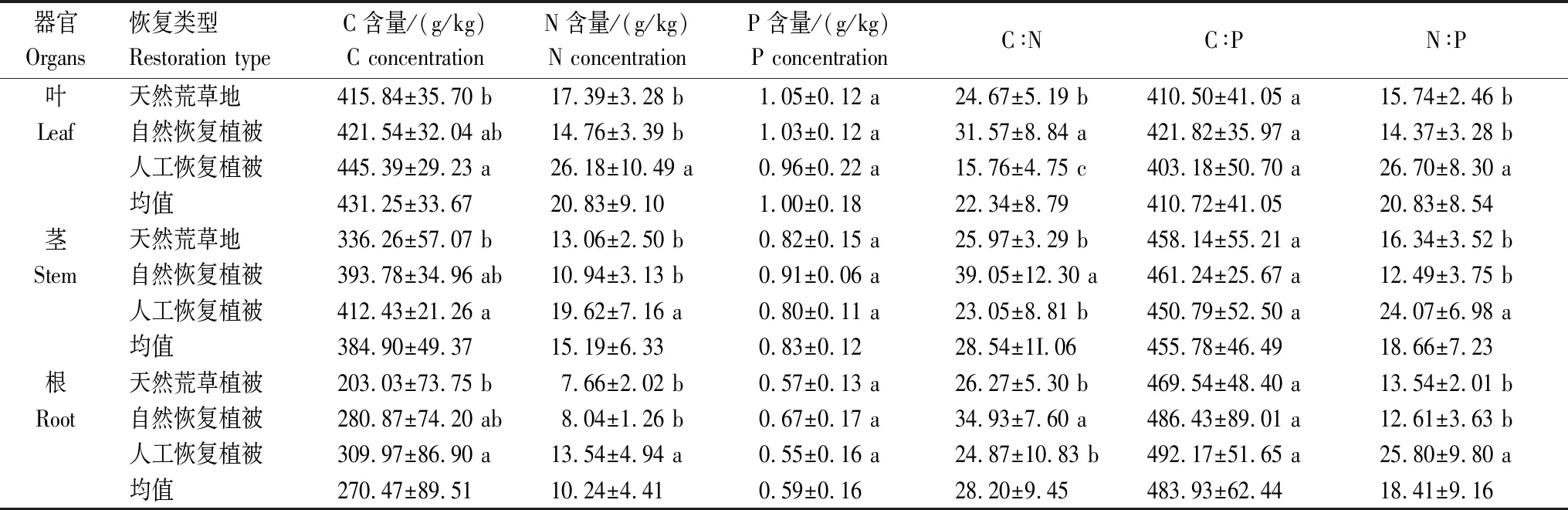
表3 3种恢复方式下植物各器官C、N、P含量及化学计量比
2.2 不同植被类型下土壤C、N、P含量及化学计量特征
土壤C、N、P含量及化学计量特征在5种典型植被间存在差异(图2)。土壤C、N含量在不同植被类型间表现为:柠条显著高于其他4种植被(P<0.05),赖草显著低于其他4种植被(P<0.05)。土壤P含量在5种典型植被间无显著差异(P>0.05)。土壤C∶N在不同植被类型间表现为:山杏>柠条>苜蓿>赖草>长芒草,但彼此间无显著差异(P>0.05)。土壤C∶P、N∶P在各植被间表现为柠条显著高于赖草(P<0.05),但柠条、赖草与长芒草、苜蓿及山杏均无显著差异(P>0.05)。从3种恢复方式整体来看(表4),自然恢复植被土壤C、N、P含量及C∶P、N∶P均显著低于人工恢复植被(P<0.05),土壤C∶N则在不同恢复方式间无显著差异(P>0.05)。
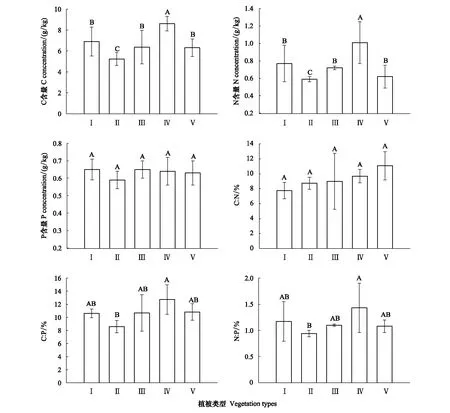
图2 不同植被类型下土壤C、N、P含量及化学计量比Fig.2 Soil C, N, P concentrations and stoichiometric ratio under different vegetation typesⅠ:长芒草 Stipa bungeana;Ⅱ:赖草 Leymus secalinus;Ⅲ: 苜蓿 Medicago sativa;Ⅳ:柠条 Caragana korshinskii;Ⅴ:山杏 Armeniaca sibirica. 不同大写字母代表不同植被类型间差异显著(P<0.05)
2.3 不同植被类型下植物与土壤C、N、P含量及化学计量特征的相关关系
对不同植被恢复类型下植物叶片与土壤C、N、P含量做Pearson相关分析发现,叶片C、N、P含量与土壤C、N、P含量的相关性在各植被类型下存在差异。如表5所示,长芒草叶的C含量与土壤C含量呈显著负相关(P<0.05),赖草叶的C含量与土壤C、N含量均呈显著负相关(P<0.05)。在人工恢复植被中,苜蓿叶的N含量与土壤C含量呈显著正相关(P<0.05),山杏叶的N含量与土壤P含量呈显著负相关(P<0.05),柠条叶片C、N、P含量与土壤C、N、P含量均无显著相关性(P>0.05),就人工植被总体而言,叶片N含量与土壤C、N含量均呈显著正相关(P<0.05),叶片P含量与土壤N含量呈显著负相关(P<0.05)。

表4 3种恢复方式下土壤C、N、P含量及化学计量比
为探究该研究区植物各器官与土壤生态化学计量特征的整体相关性,对两者C、N、P含量及化学计量特征做进一步相关分析。如图3,4所示,植物器官内,C、N、P含量及C∶N、C∶P、N∶P分别在叶、茎、根间呈显著正相关(P<0.05)。在植物与土壤间,土壤C、N、P含量与植物各器官N含量呈显著正相关(P<0.05),土壤C∶P与植物各器官C∶P呈显著负相关(P<0.05),土壤N∶P与植物各器官C∶N呈显著负相关(P<0.05),与植物各器官N∶P呈显著正相关(P<0.05)。此外,在土壤内C、N、P含量彼此间具显著正相关(P<0.05)。

图3 植物与土壤C、N、P含量相关关系Fig.3 Correlation between plant and soil C, N and P concentrationsLC,LN,LP:叶C含量, 叶N含量, 叶P含量; SC,SN,SP:茎C含量,茎N含量,茎P含量; RC,RN,RP:根C含量,根N含量,根P含量;TC,TN,TP:土壤C含量,土壤N含量,土壤P含量(* P < 0. 05, ** P < 0. 01, *** P < 0. 001)

图4 植物与土壤化学计量比相关关系Fig.4 Correlation between plant and soil C, N and P stoichiometric ratioLC.N,LC.P,LN.P:叶C∶N,叶C∶P,叶N∶P; RC.N,RC.P,RN.P:根C∶N,根C∶P,根N∶P; SC.N,SC.P,SN.P:茎C∶N,茎C∶P,茎N∶P; TC.N,TC.P,TN.P:土壤C∶N,土壤C∶P,土壤N∶P(* P < 0. 05,** P < 0. 01, *** P < 0. 001)
3 讨论3.1 不同植被类型对植物化学计量特征变化的影响
本研究对5种典型植被C、N、P含量进行比较发现,C、N、P含量在不同植被类型间存在一定差异(图1),这与Yang等[23]对温带沙漠典型植被、Zeng等[24]对黄土高原不同植被群落、李慧等[25]对晋西不同树种的研究结果相吻合。该研究区5种植被叶片C、N、P平均含量分别为431.25 g/kg、20.83 g/kg和1.0 g/kg略低于郑淑霞等[26]对黄土高原植被养分含量研究结果,这一方面与该研究区土壤较为干旱有关,另一方面与黄土高原地区植被稀疏,土壤结构疏散,植物从土壤中吸收N、P元素能力减弱有关。本研究中发现,人工植被柠条各器官N含量显著高于其他植被类型(P<0.05),这是由于柠条作为快速生长的豆科植物可通过根瘤菌固定大气中的氮[27]。在不同器官间,叶片的养分含量为最高(图1),这是叶片作为光合作用的场所既能积累大量有机质又能合成蛋白质、核酸等多种化合物的结果[28]。不同植被类型下叶、茎、根的C、N、P含量分配存在差异且C、N、P含量及化学计量特征在各器官间具显著相关性(P<0.05,图3,4),可反映不同植被可通过调节自身独特的养分分配策略来适应环境,但在生长过程中植物体叶、茎、根间的养分变化又具有协同一致性。
植物叶片C∶N、C∶P值是反映植物生长的重要指标,C∶N、C∶P值通常与植物N、P元素的养分利用效率成正比,与植物生长速率成反比[29]。本研究中人工恢复植被、自然恢复植被及天然荒草地植被的C∶N、C∶P均高于郑淑霞等[26]对黄土高原植物的研究结果,可反映该流域植被对N、P元素的利用效率较高,生长速率缓慢,其中人工恢复植被中柠条各器官C∶N、C∶P均为最低,表明柠条在生长过程中生长速率相对较快。大量研究表明植物叶片N、P含量及N∶P值可用作判断植物营养受限情况[30]。Koerselman等[6]研究表明,当N∶P<14时,植物群落生长主要受N元素限制;当14
3.2 不同植被类型对土壤生态化学计量特征的影响
本研究发现土壤C含量在5种典型植被间表现为柠条>长芒草>山杏>苜蓿>赖草(图2),这与季波等[33]对宁南不同退耕模式土壤有机碳研究结果不同,这可能与黄土高原土壤母质及气候条件有关,植物为适应其生存环境而调整自身养分分配策略进而影响土壤养分的平衡与分配。本研究发现人工恢复植被土壤C、N、P含量相对较高,自然恢复植被土壤C、N、P含量较低,说明该研究区在植被恢复过程中,人工恢复植被对土壤质量改善作用高于自然恢复植被。前人研究表明不同植物生活型,不同管理模式等对土壤养分的积累均有影响[34]。本研究发现人工植被中苜蓿植物体C、N、P含量较高,但土壤C、N、P含量较低,这与王振等[35]对黄土高原苜蓿土壤C、N、P变化研究结果不一致,这可能是由于苜蓿作为该研究区优质牧草一年被收割2—3回,不能很好的完成养分循环及再吸收造成的。土壤C∶N、C∶P是指示土壤有机质矿化速率及元素有效性的关键指标[36]。本研究中不同植被恢复类型下土壤C∶N、C∶P的平均值略低于全国土壤C∶N、C∶P水平[37],说明该研究区土壤有机质矿化效率及土壤P元素的有效性较高,养分含量较为贫乏,这是与该研究区前期严重的人为扰动及水土流失导致土壤养分条件较差有关。不同植被类型下土壤C∶N无显著差异,与欧延升等[38]对人工草地土壤C∶N值变化研究结果一致,同时此结果验证了前人研究表明土壤C∶N在生态系统中总保持相对稳定的结果[39]。
本研究对植物与土壤C、N、P含量及化学计量特征进行Pearson相关分析,发现C、N、P含量及化学计量特征在植物与土壤间均具有相关关系。例如,土壤C∶P与植物C∶P呈负相关,土壤N∶P与植物N∶P呈显著正相关(P<0.05,图3,4),表明该研究区植物生态化学计量特征受土壤化学计量特征制约,反映在半干旱黄土小流域生态系统中,植物与土壤养分变化是紧密相关相互影响的,这与An等[40]对荒漠草原植物与土壤养分相关关系研究结果一致。但本研究对不同植被类型下植物叶片与土壤C、N、P含量相关关系的研究发现,不同植被类型下叶片与土壤C、N、P含量相关关系有所差异(表3),这与罗艳等[15]对塔里木4种植被叶片与土壤生态化学计量特征研究结果一致,说明植物化学计量特征变化不仅受土壤化学计量特征影响,还与自身遗传特性有关。
4 结论
本文探究了半干旱黄土小流域3种植被恢复方式下(天然荒草、自然恢复、人工恢复)5种植被类型(长芒草草地、赖草草地、苜蓿草地、柠条灌丛、山杏林)的叶、茎、根和土壤的生态化学计量特征。结果表明,人工恢复植被叶、茎、根的C∶N、C∶P总体小于自然恢复植被,人工恢复植被生长速率相对较快。人工恢复植被苜蓿、柠条、山杏N∶P较高,生长主要受P元素限制,而自然恢复植被、天然荒草地N∶P较低,植被生长更倾向于受N、P共同限制,建议黄土高原在后期植被建设中可根据植被生长所需适量添加氮、磷肥。人工恢复植被土壤C、N、P含量及化学计量比均大于自然恢复植被,表明人工恢复植被对土壤养分改善更优于自然恢复植被,且在人工恢复植被中柠条对土壤C、N、P元素的积累效果更好,更适应于在该研究区引种来提高土壤质量。整体上,5种典型植被叶、茎、根与土壤C、N、P含量及化学计量比总体较低,植物养分吸收能力较弱,土壤有机质矿化效率及磷的有效性总体较高,养分含量较为贫乏。在研究区内,植物生态化学计量特征的变化不仅受植被类型及不同器官的影响,同时与土壤生态化学计量特征的变化紧密相关彼此制约,因此在未来植被恢复过程中应结合恢复区实际情况,因地制宜,选择合适物种,在提高植物与土壤养分含量的同时促进生态系统养分循环能力。
参考文献(References):
[1] Elser J J, Sterner R W, Gorokhova E, Fagan W F, Markow T A, Cotner J B, Harrison J F, Hobbie S E, Odell G M, Weider L W. Biological stoichiometry from genes to ecosystems. Ecology Letters, 2000, 3(6): 540- 550.
[2] 贺金生, 韩兴国. 生态化学计量学: 探索从个体到生态系统的统一化理论. 植物生态学报, 2010, 34(1): 2- 6.
[3] LeBauer D S, Treseder K K. Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology, 2008, 89(2): 371- 379.
[4] Reich P B, Oleksyn J, Wright I J. Leaf phosphorus influences the photosynthesis-nitrogen relation: a cross-biome analysis of 314 species. Oecologia, 2009, 160(2): 207- 212.
[5] Yu Q, Chen Q S, Elser J J, He N P, Wu H H, Zhang G M, Wu J G, Bai Y F, Han X G. Linking stoichiometric homoeostasis with ecosystem structure, functioning and stability. Ecology Letters, 2010, 13(11): 1390- 1399.
[6] Koerselman W, Meuleman A F M. The vegetation N∶ P ratio: a new tool to detect the nature of nutrient limitation. Journal of Applied Ecology, 1996, 33(6): 1441- 1450.
[7] Hu Y F, Shu X Y, He J, Zhang Y L, Xiao H H, Tang X Y, Gu Y F, Lan T, Xia J G, Ling J, Chen G D, Wang C Q, Deng L J, Yuan S. Storage of C, N, and P affected by afforestation withSalixcupularisin an alpine semiarid desert ecosystem. Land Degradation & Development, 2018, 29(1): 188- 198.
[8] Lambers H, Mougel C, Jaillard B, Hinsinger P. Plant-microbe-soil interactions in the rhizosphere: an evolutionary perspective. Plant and Soil, 2009, 321(1/2): 83- 115.
[9] Bui E N, Henderson B L. C∶N∶P stoichiometry in Australian soils with respect to vegetation and environmental factors. Plant and Soil, 2013, 373(1/2): 553- 568.
[10] Zechmeister-Boltenstern S, Keiblinger K M, Mooshammer M, Peuelas J, Richter A, Sardans J, Wanek W. The application of ecological stoichiometry to plant-microbial-soil organic matter transformations. Ecological Monographs, 2015, 85(2): 133- 155.
[11] 李明雨, 黄文广, 杨君珑, 李小伟. 宁夏草原植物叶片氮磷化学计量特征及其驱动因素. 草业学报, 2019, 28(2): 23- 32.
[12] Jiang Y F, Rao L, Sun K, Han Y, Guo X. Spatio-temporal distribution of soil nitrogen in Poyang lake ecological economic zone (South-China). Science of the Total Environment, 2018, 626: 235- 243.
[13] 杨阔, 黄建辉, 董丹, 马文红, 贺金生. 青藏高原草地植物群落冠层叶片氮磷化学计量学分析. 植物生态学报, 2010, 34(1): 17- 22.
[14] Song M, Peng W X, Du H, Xu Q G. Responses of soil and microbial C∶N∶P stoichiometry to vegetation succession in a karst region of southwest China. Forests, 2019, 10(9): 755.
[15] 罗艳, 贡璐, 朱美玲, 安申群. 塔里木河上游荒漠区4种灌木植物叶片与土壤生态化学计量特征. 生态学报, 2017, 37(24): 8326- 8335.
[16] 孙力, 贡璐, 朱美玲, 解丽娜, 李红林, 罗艳. 塔里木盆地北缘荒漠典型植物叶片化学计量特征及其与土壤环境因子的关系. 生态学杂志, 2017, 36(5): 1208- 1214.
[17] An S S, Darboux F, Cheng M. Revegetation as an efficient means of increasing soil aggregate stability on the Loess Plateau (China). Geoderma, 2013, 209- 210: 75- 85.
[18] Zhang W, Qiao W J, Gao D X, Dai Y Y, Deng J, Yang G H, Han X H, Ren G X. Relationship between soil nutrient properties and biological activities along a restoration chronosequence ofPinustabulaeformisplantation forests in the Ziwuling Mountains, China. CATENA, 2018, 161: 85- 95.
[19] Zheng F L. Effect of vegetation changes on soil erosion on the loess plateau. Pedosphere, 2006, 16(4): 420- 427.
[20] 于洋, 卫伟, 陈利顶, 冯天骄, 杨磊, 张涵丹. 黄土丘陵区坡面整地和植被耦合下的土壤水分特征. 生态学报, 2016, 36(11): 3441- 3449.
[21] Han F P, Ren L L, Zhang X C. Effect of biochar on the soil nutrients about different grasslands in the Loess Plateau. Catena, 2016, 137: 554- 562.
[22] 史瑞和, 鲍士旦, 秦怀英. 土壤农化分析. 北京: 中国农业出版社, 1998: 25- 97.
[23] Yang H T, Wang Z R, Li X J, Gao Y H. Vegetation restoration drives the dynamics and distribution of nitrogen and phosphorous pools in a temperate desert soil-plant system. Journal of Environmental Management, 2019, 245: 200- 209.
[24] Zeng Q C, Li X, Dong Y H, An S S, Darboux F. Soil and plant components ecological stoichiometry in four steppe communities in the Loess Plateau of China. CATENA, 2016, 147: 481- 488.
[25] 李慧, 王百田, 刘涛. 晋西黄土区不同森林树种及其林地土壤养分含量的变化. 林业科学研究, 2016, 29(4): 587- 595.
[26] 郑淑霞, 上官周平. 黄土高原地区植物叶片养分组成的空间分布格局. 自然科学进展, 2006, 16(8): 965- 973.
[27] Yang Y, Liu B R, An S S. Ecological stoichiometry in leaves, roots, litters and soil among different plant communities in a desertified region of Northern China. CATENA, 2018, 166: 328- 338.
[28] 孙美美, 关晋宏, 岳军伟, 李国庆, 杜盛. 黄土高原西部针叶林植物器官与土壤碳氮磷化学计量特征. 水土保持学报, 2017, 31(3): 202- 208.
[29] 曾昭霞, 王克林, 刘孝利, 曾馥平, 宋同清, 彭晚霞, 张浩, 杜虎. 桂西北喀斯特森林植物-凋落物-土壤生态化学计量特征. 植物生态学报, 2015, 39(7): 682- 693.
[30] Zhang W, Liu W C, Xu M P, Deng J, Han X H, Yang G H, Feng Y Z, Ren G X. Response of forest growth to C∶N∶P stoichiometry in plants and soils duringRobiniapseudoacaciaafforestation on the Loess Plateau, China. Geoderma, 2019, 337: 280- 289.
[31] Han W X, Fang J Y, Guo D L, Zhang Y. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytologist, 2005, 168(2): 377- 385.
[32] Güsewell S. N∶ P ratios in terrestrial plants: variation and functional significance. New Phytologist, 2004, 164(2): 243- 266.
[33] 季波, 李娜, 马璠, 蔡进军, 董立国, 许浩, 韩新生. 宁南典型退耕模式对土壤有机碳固存的影响. 浙江农业学报, 2017, 29(3): 483- 488.
[34] 曹娟, 闫文德, 项文化, 谌小勇, 雷丕锋. 湖南会同3个林龄杉木人工林土壤碳、氮、磷化学计量特征. 林业科学, 2015, 51(7): 1- 8.
[35] 王振, 王子煜, 韩清芳, 李文静, 韩丽娜, 丁瑞霞, 贾志宽, 杨宝平. 黄土高原苜蓿草地土壤碳、氮变化特征研究. 草地学报, 2013, 21(6): 1073- 1079.
[36] Li Y Q, Zhao X Y, Zhang F X, Awada T, Wang S K, Zhao H L, Zhang T H, Li Y L. Accumulation of soil organic carbon during natural restoration of desertified grassland in China′s Horqin Sandy Land. Journal of Arid Land, 2015, 7(3): 328- 340.
[37] 黄昌勇. 土壤学. 北京: 中国农业出版社, 2000: 8- 8.
[38] 欧延升, 汪霞, 李佳, 贾海霞, 赵云飞, 黄政, 洪苗苗. 不同恢复年限人工草地土壤碳氮磷含量及其生态化学计量特征. 应用与环境生物学报, 2019, 25(1): 38- 45.
[39] Cleveland C C, Liptzin D. C∶N∶P stoichiometry in soil: is there a “Redfield ratio” for the microbial biomass? Biogeochemistry, 2007, 85(3): 235- 252.
[40] An H, Tang Z S, Keesstra S, Shangguan Z P. Impact of desertification on soil and plant nutrient stoichiometry in a desert grassland. Scientific Reports, 2019, 9(1): 9422.
猜你喜欢
科学技术创新(2022年30期)2022-10-21
河北地质(2022年2期)2022-08-22
现代农村科技(2021年5期)2021-12-07
天然气与石油(2021年5期)2021-11-06
乡村科技(2021年16期)2021-10-21
中国煤炭工业(2020年1期)2020-12-28
科学大众(2020年17期)2020-10-27
当代陕西(2019年7期)2019-04-25
消费导刊(2017年24期)2018-01-31
中国公路(2017年11期)2017-07-31
