
长链非编码RNA在肝纤维化中的作用研究进展
2021-03-31 03:53:22张春艳颜羽昕梁洁孟根斯立木马月宏
解放军医学杂志 2021年2期
张春艳,颜羽昕,梁洁,孟根斯立木,马月宏
内蒙古医科大学基础医学院,呼和浩特 010059
肝纤维化是肝脏结缔组织异常增生的慢性病理过程,可导致细胞外基质(extracellular matrix,ECM)大量累积,包括α-平滑肌肌动蛋白(α-smooth muscle actin,α-SMA)和Ⅰ型胶原α(type Ⅰ collagen α1 chain,Col1α1),并释放促炎因子和促纤维化因子。多种慢性肝病如病毒性肝炎、酒精中毒、药物滥用、代谢综合征、遗传代谢性疾病及自身免疫性肝炎等均会导致肝纤维化[1-2]。目前肝纤维化的调节机制尚不明确,激活肝星状细胞(hepatic fibrosis,HSC)是肝纤维化发展的重要步骤。正常的HSC处于静止状态,由于某种病因损伤后HSC被激活,并被分化为成肌纤维母细胞样的细胞,从而促进纤维化的形成与发展。随着对长链非编码RNA(long non-coding RNA,lncRNA)生理作用及其在肝纤维化发病中作用机制的深入研究,通过调节肝纤维化患者体内lncRNA活性治疗肝纤维化有望成为现实。本文就lncRNA在肝纤维化中的作用研究进展进行综述。
1 LncRNA概述
1.1定义 LncRNA是长度>200 nt的竞争性内源RNA(competing endogenous RNA,ceRNA),也是RNA聚合酶Ⅱ转录的副产物,广泛存在于动物、植物、酵母甚至病毒中,无编码蛋白质的功能。LncRNA与蛋白质编码基因相似,相比miRNA,其在细胞内的转录比例更高,可在多个水平调控基因的表达,在胚胎发育、细胞分化、疾病及肿瘤的发生过程中具有重要作用[3]。
1.2分类及作用 L n c R N A 可分为5类:同义lncRNA、反义lncRNA、双向lncRNA、内含子lncRNA及基因间lncRNA[4]。LncRNA具有4种作用:(1)支架作用。与两种或两种以上的蛋白质相结合而发挥调控作用;(2)诱饵作用。诱导并结合一系列调控因子并阻碍其与相应的功能位点结合而发挥调控作用;(3)引导作用。招募特定的蛋白质并与之结合形成复合物而发挥调控作用;(4)信号作用。LncRNA可在空间和时间上反映转录因子或信号通路对基因的调节[4]。
2 促肝纤维化lncRNA
2.1肺腺癌相关转录本1(metastasis-associated lung adenocarcinoma transcript 1,MALAT1) MALAT1位于人类染色体11q13.1(小鼠染色体19qA),长约8 kb,在肿瘤细胞的增殖、迁移、侵袭中起到关键作用[5]。Yu等[6]发现,在四氯化碳(CCl4)诱导的小鼠肝纤维化模型中,MALAT1的表达在活化的HSC中明显上调,且与miR-101b的表达呈负相关。MALAT1和RAS相关C3型肉毒杆菌底物1(ras-related C3 botulinum toxin substrate 1,Rac1)是miR101b的靶标,前者可作为ceRNA增强Rac1的表达,促进HSC的增殖和活化。Dai等[7]发现,从亚砷酸盐处理的人肝细胞中提取的MALAT1可通过与miRNA-26b结合促进HSC细胞系LX-2的活化。
2.2浆细胞瘤变型异位子1(plasmacytoma variant translocation gene 1,PVT1) PVT1位于染色体8q24.21,在细胞核和细胞质中均有分布。Yu等[8]的研究发现,PV T1通过竞争性结合miR-152抑制PTCH1(patched 1),从而激活刺猬信号通路(Hedgehog signaling pathway)和上皮-间质转化(epithelial-mesenchymal transition,EMT)过程。自噬相关基因14(autophagy related gene 14,ATG14)是miR-152的直接靶标,通过PVT1-miR-152-ATG14信号通路诱导自噬有助于缺氧条件下HSC的激活,可促进肝纤维化的发展。
2.3HOX转录反义基因间RNA(HOX transcript antisense intergenic RNA,Hotair) Hotair位于12号染色体,具体位于HOXC基因座内HOXC11和HOXC12之间,包含6232个碱基对。Hotair在乳腺癌、胰腺癌、结肠癌及非小细胞肺癌等肿瘤中呈高表达[9]。Fu等[10]的研究发现,Hotair可通过与miR-148b竞争性结合而上调DNA甲基转移酶1(DNA methyltransferase 1,DNMT1),进而诱导HSC的活化。此外,Hotair还可通过抑制母体表达基因3(maternally ex-pressed gene 3,MEG3)的表达而促进HSC的活化。Yu等[11]的研究发现,在CCl4诱导的肝纤维化小鼠中Hotair表达增加。Hotair通过下调miR-29b的表达减弱其介导的表观遗传机制,从而导致DNA甲基转移酶3b(DNA methyltransferase 3b,DNMT3b)、第10号染色体缺失的磷酸酶及张力蛋白同源基因(phosphatase and tensin homolog deleted on chromosome ten,PTEN)的甲基化增强,部分促进HSC活化。
2.4肝纤维化相关lncRNA1(liver-enriched fibrosisassociated lncRNA1,LFAR1) LFAR1位于小鼠基因组的一个区域,该区域与人类染色体4q25同源,并与人CYP2U1和HADH基因相邻。Zhang等[12]的研究发现,LFAR1可促进Smad2/3与转化生长因子-β受体1(transforming growth factor beta receptor 1,TGFβR1)的结合及其在细胞质中的磷酸化,还可直接与Smad2/3结合促进Smad2、Smad3、Notch2、Notch3的转录。因此,LFAR1可通过激活转化生长因子-β(transforming growth factor-β,TGF-β)和Notch通路,促进肝纤维化。
2.5核富含转录本1(nuclear-enriched abundant transcript 1,NEAT1) NEAT1是长约3.7 kb的lncRNA,主要富集于细胞核中,是参与构成细胞核亚结构的重要RNA分子[13]。Yu等[14]发现,NEAT1和Kruppel样因子6(Kruppel-like factor 6,KLF6)是miR-122的靶标。NEAT1可通过竞争性结合miR-122来调节肝纤维化中KLF6的表达。同时,NEAT1/miR-122/KLF6在HSC激活过程中起关键作用。Kong等[15]发现,胰岛素样生长结合蛋白相关蛋白1(insulin-like growth factor binding proteinrelated protein 1,IGFBPrP1)可增高小鼠肝脏组织中NEAT1、自噬相关蛋白9a(autophagy associated proteins 9a,Atg9a)的表达及自噬水平,降低miR-29b的表达水平。其中Atg9a参与了IGFBPrP1诱导的HSC自噬和激活。因此,NEAT1/miR-29b/Atg9a调控轴参与了IGFBPrP1诱导的小鼠HSC的自噬和 激活。
2.6被TGF-β活化的长链非编码RNA(long noncoding RNA activated by TGF-β,lncRNA ATB) LncRNA ATB位于14号染色体上,是长度超过8 kb的lncRNA。在丙型肝炎病毒(hepatitis C virus,HCV)相关肝纤维化患者的肝脏组织、血清及HSC中lncRNA ATB表达均上调[16-17]。LncRNA ATB通过竞争性结合miRNA-425-5p而促进Smad2的表达以及HSC的增殖和活化[16]。Fu等[17]发现,在HCV相关肝纤维化患者的肝脏组织、血清及HSC中miR-200a表达下降,β-连环蛋白(β-catenin)表达上调。β-catenin是miR-200a的靶标,lncRNA ATB作为ceRNA可与miR-200a结合,促进β-catenin的表 达[17]。因此,lncRNA ATB/miR-200a/β-catenin调节轴可能促进了肝纤维化的发展。
2.7长链非编码RNA H19(long non-coding RNA H19,lncRNA H19) LncRNA H19位于人类同源区域染色体11p15.5,为长约2.3 kb的lncRNA。Song等[18]发现,上皮细胞黏附分子(epithelial cells adhere to molecules,EpCAM)是电子盒同源盒1(electronic box homologous box 1,ZEB1)的下游靶基因,lncRNA H19和EpCAM的向上调节与ZEB1的向下调节呈正相关。LncRNA H19通过激活ZEB1/EpCAM信号通路而促进小鼠的肝纤维化。Zhu等[19]的研究发现,lncRNA H19作为ceRNA可与miR-148a相结合,并随后维持泛素特异性蛋白酶4(ubiquitin-specific protease 4,USP4)的水平,USP4是miR-148a的一个已确定的靶标。该研究结果揭示了一个新的lncRNA H19/miR-148a/USP4轴,该轴通过HSC的激活和肝细胞中TGF-β途径来促进肝纤维化。Wang等[20]发现,lncRNA H19通过上调酒精脱氢酶Ⅲ(alcohol dehydrogenase Ⅲ,ADH3)介导的维甲酸信号从而诱导HSC的激活;Huang等[21]发现,在转染IGFBPrP1的JS-1细胞中,细胞自噬活跃程度与lncRNA H19表达水平呈正相关。因此,lncRNA H19可通过磷脂酰肌醇-3激酶(phosphatidylinosital 3-kinase,PI3K)/蛋白激酶B(protein kinase B,Akt)/哺乳动物雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)信号通路来促进IGFBPrP1诱发HSC自噬。
2.8小核RNA宿主基因7(small nucleic RNA host gene 7,SNHG7) SNHG7位于9号染色体,长度为2157 nt,在人类肝纤维化组织和活化的小鼠原代HSC中表达上调。Yu等[22]发现,lncRNA SNHG7可竞争性结合miR-378-3p,致使细胞内游离的紊乱片段极性蛋白2(disorder fragment polarity protein 2,DVL2)含量增加,β-catenin在细胞内大量积累,促进了HSC的激活,诱导了肝纤维化的发展。
2.9INK4位点的反义非编码RNA(antisense noncoding RNA in the INK4 locus,ANRIL) ANRIL位于7号染色体,首先发现于遗传性黑色素瘤和神经系统肿瘤患者。Yang等[23]发现,在活化的HSC和肝纤维化组织中ANRIL的表达显著降低,DNA甲基转移酶3A(DNA methyltransferase 3A,DNMT3A)的表达显著增加。下调DNMT3A可增加活化的HSC中ANRIL的表达,而过表达ANRIL可抑制HSC的激活和腺苷酸活化蛋白激酶(AMP-activated protein kinase,AMPK)信号通路。因此,lncRNA ANRIL缺失可以激活AMPK途径,从而促进肝纤维化和HSC的活化。
2.10软骨发生刺激因子1(stimulator of chondrogenesis 1,SCRG1) SCRG1是一种转录长度为3118 bp的lncRNA,在人肝硬化组织中其表达上调13.62倍。Wu等[24]发现,SCRG1在人体组织和转化生长因子-β1(transforming growth factor-1,TGF-β1)诱导的活化LX-2细胞中的表达,可随着肝纤维化的发展而增加。SCRG1与锌指蛋白36(tristetraprolin,TTP)有结合位点,并可特异性结合TTP蛋白。过表达SCRG1会导致TTP mRNA不稳定,蛋白质表达减少。SCRG1靶向下调TTP可导致肿瘤坏死因子α (tumor necrosis factor alpha,TNF-α)和基质金属蛋白酶2(matrix metalloproteinase,MMP2)的激活,促进肝纤维化的发展。
2.11X染色体失活特异转录物(X-inactive specific transcript,XIST) XIST位于X染色体无活性中心区域,长约16 500 nt,作为染色体Xq13.2的转录产物,能够影响X染色体相关基因的激活。XIST与miR-29b结合可促进高迁移率族蛋白盒-1(highmobility group box-1,HMGB1)的表达,从而增强乙醇诱导的LX-2细胞自噬和激活[25-26]。因此,lncRNA XIST通过miR-29b/HMGB1途径增强乙醇诱导的LX-2细胞的自噬和激活,从而促进肝纤维化的 发展。
2.12小Cajal体特异性RNA 10(small Cajal bodyspecific RNA 10,SCARNA10) SCARNA10位于12号染色体,是一种核内滞留的lncRNA,也是一种潜在的生物标志物。SCARNA10过表达可加重CCl4诱导的小鼠肝纤维化,且SCARNA10在小鼠肝纤维化组织中表达显著上调[27]。SCARNA10在体内、体外均可通过诱导肝细胞凋亡和HSC活化而促进肝纤维化的发展。Zhang等[28]发现,SCARNA10可与多梳抑制复合物2(polycomb repressive complex 2,PRC2)结合而增加Smad2、Smad3、TGF-β1和KLF6的表达,从而促进肝纤维化的发展。
2.13长链基因间非编码RNA01093(long intergenic noncoding RNA 01093,linc01093) Linc01093位于4号染色体。目前主要通过芯片、高通量测序技术筛选肿瘤组织中差异表达的linc01093。Tang等[29]发现,在CCl4诱导的肝组织和TGF-β1刺激的肝细胞中linc01093表达下调。Linc01093的表达下调促进了肝细胞的凋亡,抑制了肝细胞的存活。在TGF-β1的刺激下,下调的linc01093通过促进沉默信息调节因子1 (silent information regulator 1,SIRT1)的降解和泛素化来促进肝细胞凋亡。
2.14Alu介导的p21转录调节因子(alu-mediated p21 transcriptional regulator,APTR) APTR位于染色体7q21,长度为2303 bp,对细胞周期和细胞增殖具有抑制作用。Yu等[30]发现,APTR在肝纤维化组织中表达上调,其沉默可诱导p21的转录,从而抑制U87细胞的生长。APTR与p21在小鼠肝纤维化组织中的表达呈负相关,可通过负调控p21加速细胞周期,对HSC具有促增殖的作用。
2.15核糖核酸HOXA转录本(ribonucleic acid HOXA transcript at the distal tip,HOTTIP) HOTTIP位于染色体7p152,在多种人类肿瘤(包括胃癌、肝细胞癌和结直肠癌)中起关键作用[31-32]。Li等[33]发现,HOTTIP对miR-148a具有负调控作用,作为miR-148a、TGF-βR1和TGF-βR2的新靶标,可增强TGF-β/Smad信号传导。HOTTIP可通过下调miR-148a,增强TGF-βR1和TGF-βR2的表达,从而促进肝纤维化的发生和发展。Zheng等[34]发现,HOTTIP作为miR-150的ceRNA,可增加血清反应因子(serum response factor,SRF)的表达,诱导小鼠HSC的活化。
2.16牛磺酸调节基因1(taurine up-regulated gene 1, TUg1) TUg1是大小为7.1 kb的lncRNA,首次发现于牛磺酸处理的小鼠视网膜细胞上调表达基因中。Han等[35]发现,TUg1在CCl4及胆管结扎(BDL)诱导的肝纤维化小鼠模型的肝脏组织以及肝硬化患者中表达上调。TUg1可促进α-SMA、Col1α1、MMP2/MMP9/MMP10和基质金属肽酶组织抑制因子1(tissue inhibitor of metalloproteinases 1,TIMP1)的表达。miR-29b是TUg1的靶标,TUg1可负向调节miR-29b的表达。因此,lncRNA TUg1作为ceRNA可下调miR-29b的表达而促进HSC的活化,从而促进肝纤维化的发展。
LncRNA促进肝纤维化的作用汇总见表1。
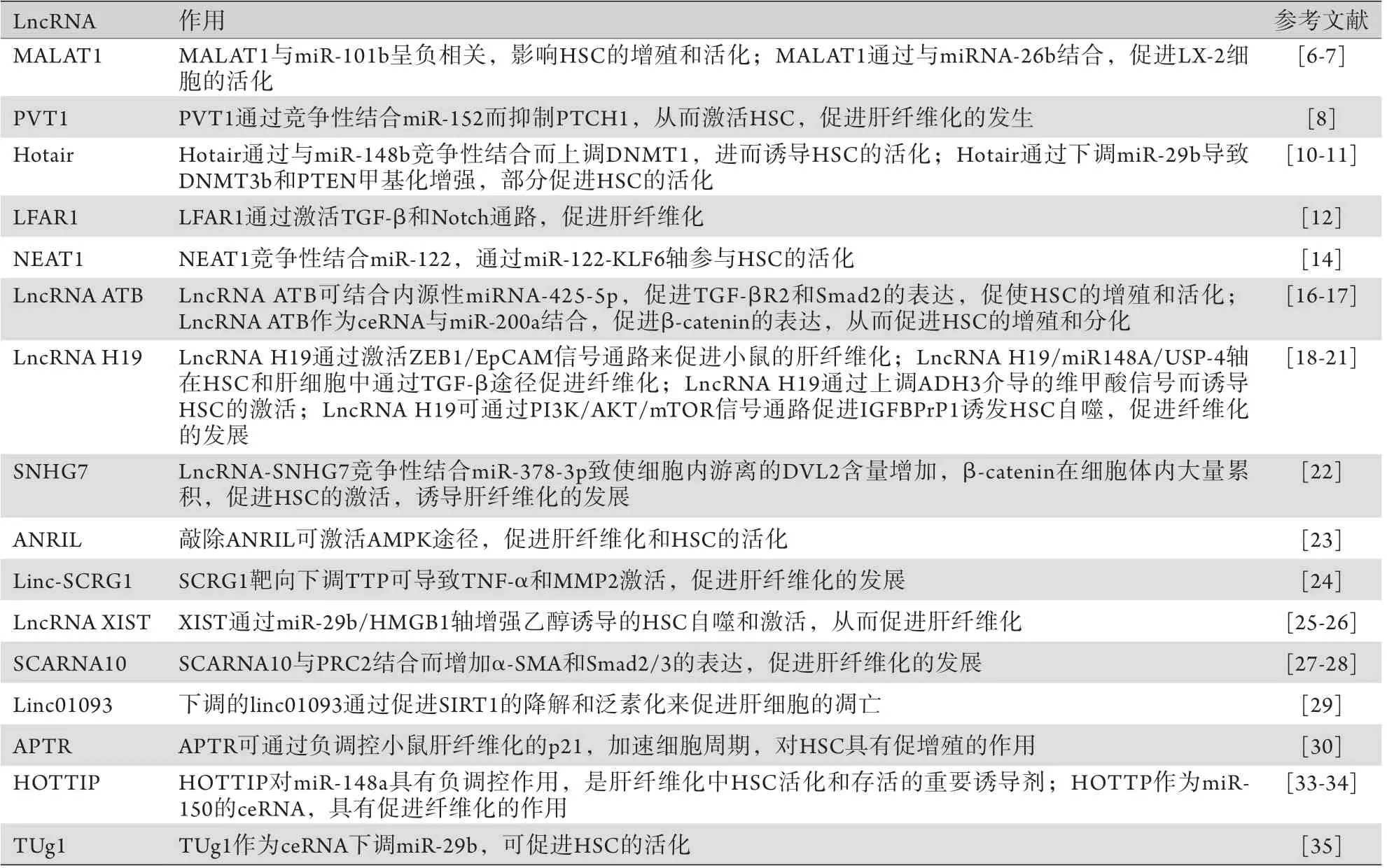
表1 LncRNA促进肝纤维化的作用Tab.1 The promotive effects of lncRNA on liver fibrosis
3 抑制肝纤维化lncRNA
3.1母系表达基因8(maternally expressed gene 8,MEG8) MEG8位于染色体14q32.3上的一簇印迹基因中,主要分布在LX-2细胞和AML12细胞的细胞核内。MGE8可促进肺癌和胰腺癌细胞EMT的表观遗传学进程,抑制血管平滑肌细胞(vascular smooth muscle cell,VSMC)和滋养细胞的增殖和迁移[36-37]。研究发现,MEG8在活化的HSC、损伤的肝细胞和肝纤维化组织中过表达[38]。MEG8可通过抑制Notch通路而抑制肝细胞的HSC活化和EMT过程[38]。MEG8作为Notch信号的新型调节剂,为肝纤维化的分子机制研究提供了新的理论依据。
3.2预测基因5091(predicted gene 5091,Gm5091) Gm5091是一种长度为1179 bp的基因间lncRNA,位于Chr17。乙醇处理可使Gm5091在HSC中的表达下调,并对乙醇诱导的HSC激活和炎症产生负调控作用。Zhou等[39]发现,Gm5091序列包含miR-27b、miR-23b、miR-24的结合位点,且通过RNA下拉测定法证实miR-27b/23b/24均可与Gm5091结合。全长Gm5091可降低miR-27b/23b/24的水平,但截短的Gm5091不能删除结合位点。Gm5091/miR-27b/23b/24可以减轻酒精性肝纤维化(alcoholic hepatic fibrosis,AHF)。
3.3长链基因间非编码RNA-p21(long intergenic noncoding RNA-p21,lincRNA-p21) LincRNA-p21位于编码关键细胞周期调节因子Cdkn1a(也称为p21)的基因上游约15 kb,包含两个外显子,长约3.1 kb。LincRNA-p21可作为TGF-β的效应器与miR-30相互作用,从而促进小鼠的肝纤维化[40];还可作为ceRNA与miR-181b竞争性结合,减弱miR-181b对PTEN表达的抑制作用,最终抑制HSC的活化[41]。Yu等[42]发现,lincRNA-p21作为ceRNA可与miR-17-5p相结合,上调糖原合成激酶3β(glycogen synthesis kinase 3β,GSK-3β)的表达,抑制Wnt/β-catenin信号通路。因此,通过lincRNA-p21/miR-17-5p/β-catenin信号通路轴,lincRNA-p21能够抑制HSC的活化。
3.4长链非编码RNA-Hser(long non-coding RNAHser,lnc-Hser) Lnc-Hser主要位于初级肝细胞和AML12细胞的细胞核中。Zhang等[43]发现,敲除lnc-Hser可通过诱导EMT和肝细胞凋亡在体内和体外加重肝纤维化。同时,lnc-Hser可通过补体C5a受体1(complement C5a receptor 1,C5AR1)-河马(Hippo)-YES相关蛋白(YES-associated protein,YAP)途径抑制HSC的凋亡,并通过Notch途径抑制肝细胞内EMT的累积。总之,lnc-Hser是受损肝细胞的新型生物标志物,也是抗纤维化治疗的潜在靶标。
3.5生长停滞特异性转录本5(growth arrest-specific transcript 5,GAS5) GAS5最初是因在生长抑制的鼠纤维原细胞中呈高表达而被发现的,位于人染色体1q251的小开放阅读框,全长630个核苷酸。 Yu等[44]和Dong等[45]发现,CCl4诱导的大鼠肝纤维化模型中,miR-23a可通过PTEN/PI3K/Akt/mTOR/Snail信号通路促进纤维化的发展。GAS5可与miR-23a竞争性结合,降低miR-23a的表达,从而抑制肝纤维化。GAS5/miR-23a/PTEN/PI3K/Akt/mTOR/Snail通路在肝纤维化中的作用为肝纤维化的治疗提供了潜在的分子靶点。
3.6母系表达基因3(maternal expressed gene 3,MEG3) MEG3位于人类染色体14q32.3的DLK1-MEG3位点,长度为35 kb,具有抑癌作用。Yu等[46]的研究发现,MEG3是一种肿瘤抑制基因,在EMT过程中发挥重要作用。MEG3在体内和体外肝纤维化过程中表达均降低,其表达恢复后,肝纤维化得到抑制,α-SMA和Col1α1表达降低。值得注意的是,MEG3过表达抑制了通过EMT激活的HSC,这与上皮标志物的增加和间质标志物的减少有关。进一步研究发现,刺猬信号通路介导的EMT过程在肝纤维化的发生发展中具有重要作用,平滑蛋白(smoothened,SMO)是Hh途径的一员[46]。此外,MEG3被确认为是miR-212的靶标。miR-212部分参与了MEG3对EMT过程的影响。总之,MEG3通过SMO蛋白和miR-212抑制了刺猬信号通路介导的肝纤维化EMT过程。
LncRNA抑制肝纤维化的作用汇总见表2。
4 总结与展望
LncRNA是近年肝病领域研究的热点,在肝癌(liver cancer,HCC)、肝纤维化、非酒精性脂肪肝(non-alcoholic fatty liver disease,NAFLD)中的研究取得了突破性进展。但多数lncRNA在肝纤维化中的作用机制尚未完全阐明。缺失上调的lncRNA或过度表达下调的lncRNA是进行lncRNA基础治疗的主要机制。未来研究应致力于寻找准确地将lncRNA导入体内的方法并深入研究其机制;准确鉴定lncRNA的靶标,使导入体内的lncRNA作用于特定的与肝纤维化发展相关的靶标。同时,希望临床研究人员进一步提高对lncRNA在肝纤维化调控中作用的认识,从而有效地筛选lncRNA用于肝纤维化的诊断和治疗,为临床试验提供理论依据。

表2 LncRNA抑制肝纤维化的作用Tab.2 The inhibitory effects of lncRNA on liver fibrosis
猜你喜欢
昆明医科大学学报(2022年2期)2022-03-29 00:52:18
军民两用技术与产品(2021年10期)2021-03-16 06:05:10
世界农药(2019年3期)2019-09-10 07:04:10
科学之谜(2019年3期)2019-03-28 10:29:44
科学之谜(2018年8期)2018-09-29 11:06:46
光学精密工程(2016年4期)2016-11-07 09:04:48
恋爱婚姻家庭·养生版(2016年9期)2016-09-07 11:25:01
癌变·畸变·突变(2016年3期)2016-02-27 06:15:36
肿瘤影像学(2015年3期)2015-12-09 02:38:45
哈尔滨医药(2015年4期)2015-12-01 03:57:54
