
绵羊Musclin基因真核表达载体的构建及其对肌细胞糖代谢和胰岛素作用的影响
2021-03-30 09:20郑腾飞韩高链李孟心李俊玲
畜牧兽医学报 2021年3期
郑腾飞,辛 香,韩高链,李孟心,李俊玲,秦 健,2,杜 荣*
(1.山西农业大学动物医学学院,太谷030801; 2.山西农业大学 实验教学中心,太谷030801)
Musclin是Nishizawa等[1]于2004年在小鼠中首先发现的一种骨骼肌衍生分泌因子。其蛋白分子量为11 ku,包含130个氨基酸残基,其N端存在一个包含30个氨基酸残基的信号肽。Musclin参与糖代谢,对胰岛素刺激的葡萄糖摄取和糖原合成功能有抵抗作用[2-4]。Nishizawa等[1]在肥胖导致的胰岛素抵抗KKAy小鼠的肌肉中观察到MusclinmRNA的高表达。Zhang等[5]和Chen等[6]的研究结果也表明,Musclin与肥胖导致的胰岛素抵抗等代谢性疾病有关。蛋白激酶B(AKT)[7-9]和葡萄糖转运蛋白(GLUT)[10-12]是糖代谢通路中重要的两个因子。在胰岛素信号通路中,Musclin能够降低AKT的活性,从而使GLUT蛋白失活[2]。Musclin的表达与机体运动有关[13]。运动能够改变Musclin的表达,延缓癌症恶病质小鼠的肌肉萎缩,也能通过升高Musclin调控GLUT4的活性,改善高脂饮食喂养大鼠的脂质代谢和胰岛素敏感性[14-15]。然而,Musclin在葡萄糖代谢和胰岛素抵抗中的作用机制目前尚不清楚,是否与糖代谢关键因子胰岛素受体底物(IRS1)[16-17]和糖原合成酶激酶3β(GSK3β)[18]的表达有关尚不清晰。而且,目前有关Musclin的研究大多以小鼠或大鼠为研究对象[19],而针对大型哺乳动物的研究报道很少。Chen和Gallo-Villegas等[20-21]研究发现,人Musclin可通过紊乱葡萄糖代谢,导致胰岛素抵抗发生。针对家畜,虽然有研究人员对兔[22]、牛[23]、猪[24]Musclin基因进行了克隆和初步分析,但诸如糖代谢调节及其机制的深入研究鲜见报道。Li等[25]鉴定了能够影响绵羊Musclin基因转录调控的重要元件。但有关绵羊Musclin基因结构和性质的生物信息学分析及对糖代谢和胰岛素作用的影响尚未阐明。
本试验旨在利用生物信息学软件分析和预测绵羊Musclin蛋白的结构特征,为进一步研究绵羊Musclin的生物功能提供依据。同时以绵羊成肌细胞为研究对象,通过构建和转染绵羊Musclin基因真核表达载体实现过表达,研究绵羊Musclin基因对正常或胰岛素刺激状态下肌细胞糖代谢的影响及相关基因表达的调控作用,为揭示Musclin对绵羊肌肉发育的作用及机制奠定基础,也为通过调控营养代谢改善绵羊肌肉生长性能提供依据。
1 材料与方法
1.1 试剂与耗材
Lipofectamine 2000 购自Invitrogen 公司;D-MEM/F-12、新生牛血清、Pen Strep均购自Gibco公司;胰酶、胰岛素、FBS均购于Sigma公司;马血清购自四季青公司;DL5000 DNA Marker、pMD18-T Vector、BamH I、XhoI、Trizol、T4 DNA连接酶、反转录试剂、电泳用琼脂糖、Real time PCR试剂和八连管均购于TaKaRa公司;凝胶回收试剂盒购于OMEGA公司;EndoFree Plasmid Mini Kit购于北京全式金生物技术科技有限公司;60 mm培养皿、6孔 板、冻存管均购于Corning公司;葡萄糖含量试剂盒和糖原含量试剂盒均购自苏州科铭;感受态细胞购于山西赛奥生物科技有限公司;pcDNA3.1质粒和绵羊成肌细胞由本实验室保存。
1.2 绵羊Musclin生物信息学分析
利用表1中的生物信息学在线网站和软件,对绵羊Musclin蛋白的结构特征等进行分析。
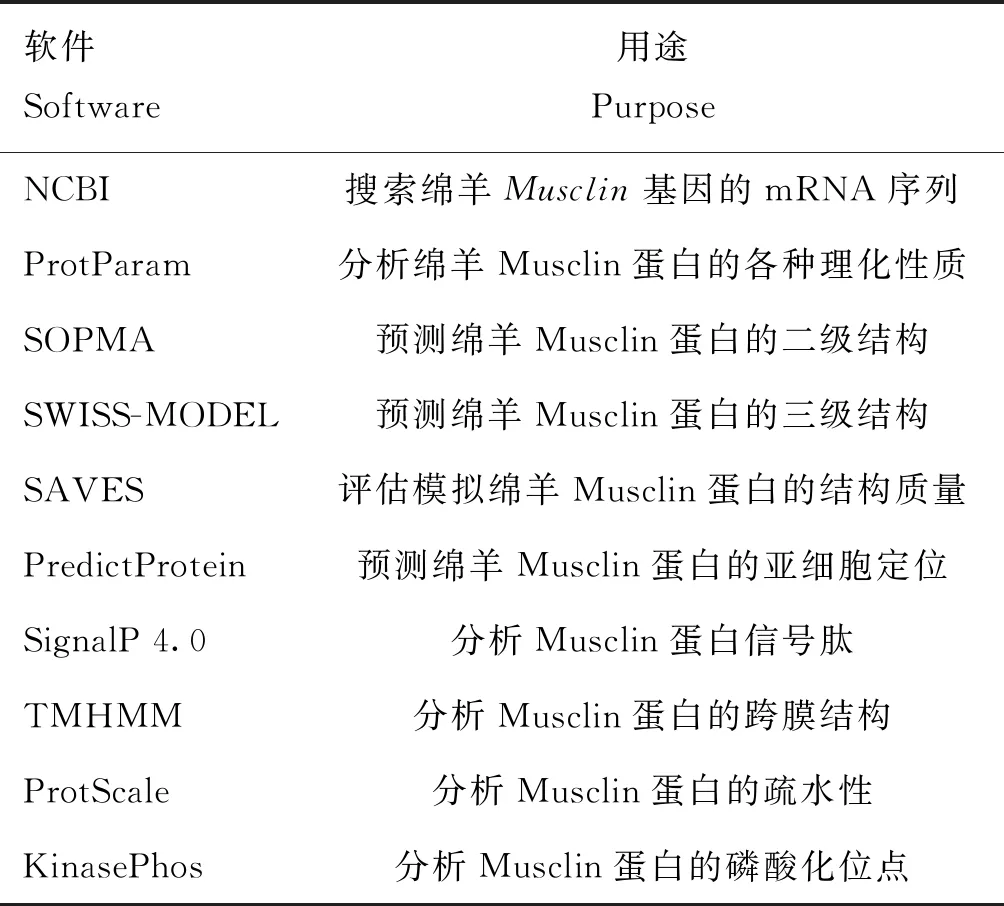
表1 生物信息学分析软件
1.3 绵羊Musclin cDNA的克隆
根据绵羊MusclinCDS区,合成gBlock序列。根据NCBI中绵羊Musclin的基因序列(XM_004003038.4),在NCBI上设计引物。产物总长度为435 bp,其中上游引物序列(5′→3′)F:CAAGATCCGGAATTCGCCACCATGCTGGAC;下游引物序列(5′→3′)R:TCCTAGCCGCTCGAGTTAGCCTCTGGAATT。起始密码子ATG前加kozak序列和酶切位点BamH I,终止密码子端加酶切位点XhoI,两端分别加上保护碱基。然后PCR扩增、胶回收,并连接至pMD18-T载体上。将克隆载体37 ℃过夜摇菌,提取质粒进行双酶切鉴定并测序。
1.4 绵羊pcDNA3.1-Musclin真核表达载体的构建与鉴定
对测序正确的克隆载体以及真核表达载体pcDNA3.1进行BamH I-XhoI双酶切,1%琼脂糖凝胶电泳胶回收后连接构建pcDNA3.1-Musclin真核表达载体。连接体系和反应条件:pcDNA3.1 50 ng,目的基因Musclin50 ng,T4 DNA连接酶0.5 μL,10× T4 DNA连接酶缓冲液1 μL,补ddH2O至10 μL; 混匀后16 ℃过夜连接。将连接产物转化到E.coliDH5α感受态细胞后挑选阳性克隆,37 ℃过夜摇菌,提取质粒,BamH I-XhoI双酶切鉴定并测序。
1.5 绵羊成肌细胞培养和转染
将本实验室保存的绵羊胎儿成肌细胞复苏后,用含有10%新生牛血清和1%青霉素-链霉素的DMEM培养基,在37 ℃、5% CO2的细胞培养箱中培养。质粒转染根据Lipofectamine 2000的说明操作进行:在转染前1 d按照2.2×105个·孔-1的密度将细胞接种到6孔板。第2天,更换新鲜培养基800 μL,然后将2.5 μg质粒与200 μL DMEM培养基或7 μL Lipofectamine 2000各自混匀并在室温下孵育5 min,然后混合并继续孵育20 min,加到更换新鲜培养液的以上贴壁细胞上,在37 ℃、5% CO2条件下继续培养。
1.6 细胞分组和处理方法
细胞共分为4组:空载体组(pcDNA3.1)、Musclin过表达组(pcDNA3.1-Musclin)、空载体+胰岛素(80 nmol·L-1)组(pcDNA3.1 + Insulin)、Musclin过表达+胰岛素(80 nmol·L-1)组(pcDNA3.1-Musclin + Insulin)。每组3个重复。质粒转染6 h后,更换为含2%马血清的分化培养基诱导分化,并根据分组方案添加胰岛素或PBS。在48、72 h后收集培养液和细胞,用于后续检测。
1.7 各处理组葡萄糖和糖原含量检测
收取分化48和72 h时的培养液和细胞到2 mL 离心管。超声波破碎细胞,离心取上清。调节酶标仪至特定波长,测定之后,依照各试剂说明书计算各组的葡萄糖和糖原含量。
1.8 各处理组RNA的提取及荧光定量PCR
用Trizol法(TaKaRa)提取绵羊成肌细胞的总RNA,并使用PrimeScriptTMRT试剂盒(Perfect Real Time,TaKaRa)进行反转录。反转录体系:Mix 2 μL、RNA 4 μL、RNase Free ddH2O 4 μL。然后使用Power SYBR Green Kit(TaKaRa)在BIO-RAD定量PCR仪上进行荧光定量PCR反应。以β-actin基因为内参基因(表2)。

表2 内参和目的基因引物序列
荧光定量扩增体系为10 μL:SYBR Premix Ex TaqⅡ(2×)5 μL、Forward primer 0.4 μL、Reverse primer 0.4 μL、cDNA 1 μL、RNase-free water 3.2 μL;扩增程序:95 ℃ 2 min;95 ℃ 30 s,60 ℃ 34 s,共40个 循环。
1.9 数据分析
通过2-ΔΔCT方法计算基因相对表达量。使用SPSS 19.0软件单因素方差分析差异显著性,所有数据均表示为“Mean±SD”。P<0.05表示差异显著;P<0.01表示差异极显著。并使用GraphPad Prism 5软件制作图表。
2 结 果
2.1 绵羊Musclin的生物信息学分析
2.1.1 绵羊Musclin蛋白的理化性质 利用ProtParam在线网站对绵羊Musclin蛋白的理化性质进行分析,计算出绵羊Musclin基因编码132个氨基酸,分子式为C640H1048N188O189S4,预测分子量为14 528.75 u,原子总数为2 069。理论等电点为10.15,消光系数(280 nm)为0.757。不稳定系数为42.12。脂肪系数为90.08,亲水性平均系数为-0.234。
2.1.2 绵羊Musclin蛋白的二级和三级结构预测及其评估 通过SOPMA在线网站对绵羊Musclin蛋白进行二级结构预测,结果显示,α-螺旋(α-helix) 有52个,占39.39%;β-折叠(β-sheet)有13个,占9.85%;β-转角(β-turn)有8个,占6.06%;无规卷曲(random coil)有59个,占44.7%(图1A)。应用在线网站SWISS-MODEL预测绵羊Musclin结构域区的三级结构。结果显示,三级结构主要由α-螺旋和无规卷曲组成,与二级结构相吻合(图1B)。利用SAVES在线网站评估模拟蛋白结构的质量(图1C)。结果显示,残基在红色区域[A,B,L]的比例为88.0%,在黄色区域[a,b,l,p]的比例为4.0%,在浅黄区域[~a,~b,~l,~p]的比例为4.0%,在白色区域的比例为4.0%。其中红色区域中残基的原子之间空间位阻最小。残基在红色区域的比例接近90%,说明模拟的蛋白质结构质量较好。
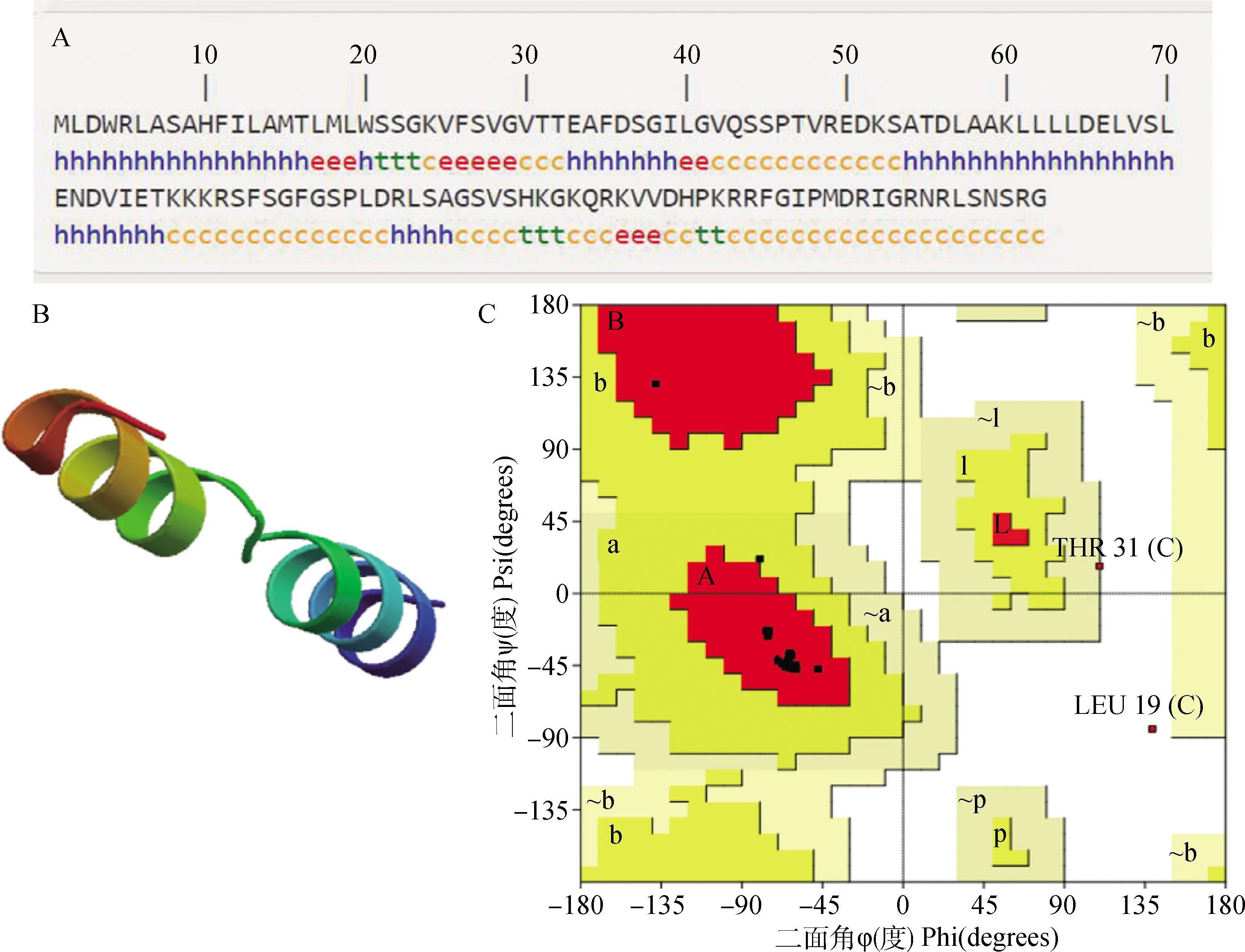
图A中,h为α-螺旋,e为β-折叠,t为β-转角,c为无规卷曲。图C中,黑色小斑点表示残基,红色区域为最佳合理区,黄色区域为较合适区,浅黄色区域为勉强接受区,白色区域为不合理区 In figure A, h is α-helix, e is extended strand, t is β-turn and c is random coil; In figure C, the small black spots are residues, the red regions are the most favoured regions, the yellow regions are the appropriate regions, the light yellow regions are the barely permitted regions and the white regions are the disallowed regions图1 绵羊Musclin蛋白的二级(A)和三级结构(B)及蛋白结构的拉式构象图(C)Fig.1 Secondary structure(A), tertiary structure(B) and ramachandran plot(C) of Musclin protein in sheep
2.1.3 绵羊Musclin蛋白亚细胞定位及信号肽分析 应用在线网站PredictProtein预测绵羊Musclin蛋白的亚细胞定位,绿色部分表示Musclin蛋白定位情况,结果显示,绵羊Musclin蛋白主要定位于细胞外(图2A)。经SignalP分析,绵羊Musclin蛋白存在信号肽,1~27位氨基酸为信号肽位置,在28氨基酸位点处存在1个酶切位点(图2B)。用TMHMM分析绵羊Musclin蛋白跨膜结构发现,绵羊Musclin蛋白不存在跨膜区(图2C)。综合以上结果,说明绵羊Musclin为分泌性蛋白。
2.1.4 绵羊Musclin蛋白疏水性和磷酸化位点分析 使用ProtScale在线网站分析蛋白的疏水性(图3A),结果显示,最大疏水指数为2.622,最大亲水指数为-2.211,但从整体来看氨基酸序列有明显的亲水性区域,表明该蛋白有较强的亲水性,因此预测此蛋白为可溶性蛋白。KinasePhos在线网站预测结果表明,该蛋白的潜在磷酸化位点为3个丝氨酸激酶(82、88、128),2个苏氨酸激酶(47、77),但并没有酪氨酸激酶位点(图3B)。
2.2 绵羊Musclin基因真核表达载体的酶切鉴定和测序鉴定
用BamH I -XhoI双酶切pcDNA3.1-Musclin,得到近500(Musclin)和5 000 bp的两条带,说明连接成功(图4)。将测序结果与登录号为XM_004003038.1的绵羊MusclinCDS序列在DNAMAN软件中进行比对,比对结果正确,表明载体构建成功,可进行后续试验。
2.3 不同处理对绵羊肌细胞Musclin mRNA表达的影响
细胞分化48 h时,与空载体组相比,Musclin过表达组的MusclinmRNA表达量升高了4 056.77倍(P<0.01);与空载体+胰岛素组相比,Musclin过表达+胰岛素组的MusclinmRNA表达量升高了3 202.59倍 (P<0.01,图5)。
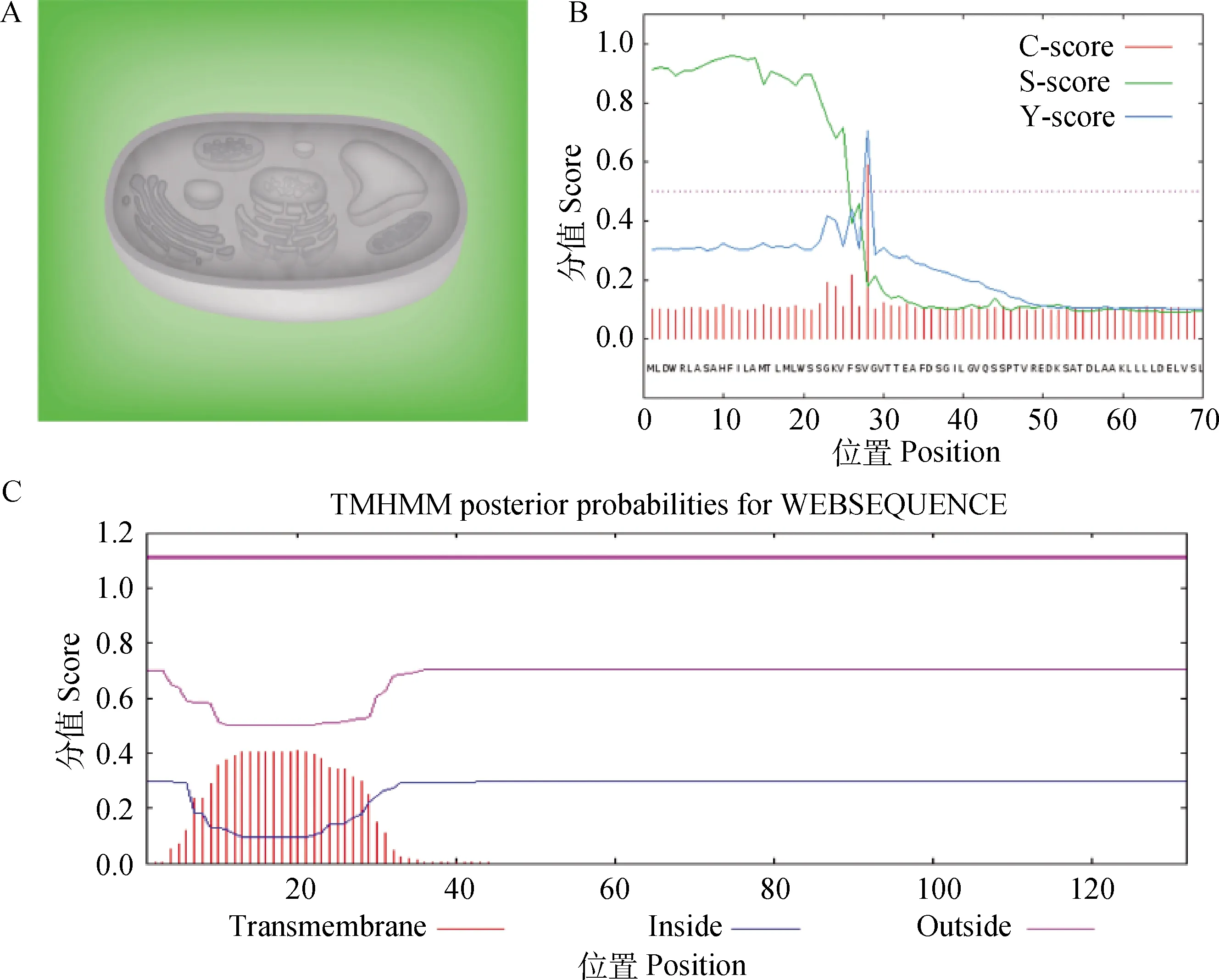
A. 亚细胞定位;B. 信号肽;C. 跨膜区 A. Subcellular localization; B. Signal peptide; C. Transmembrane图2 绵羊Musclin蛋白亚细胞定位及信号肽与跨膜区分析Fig.2 Subcellular localization, signal peptide and transmembrane analysis of the Musclin protein in sheep
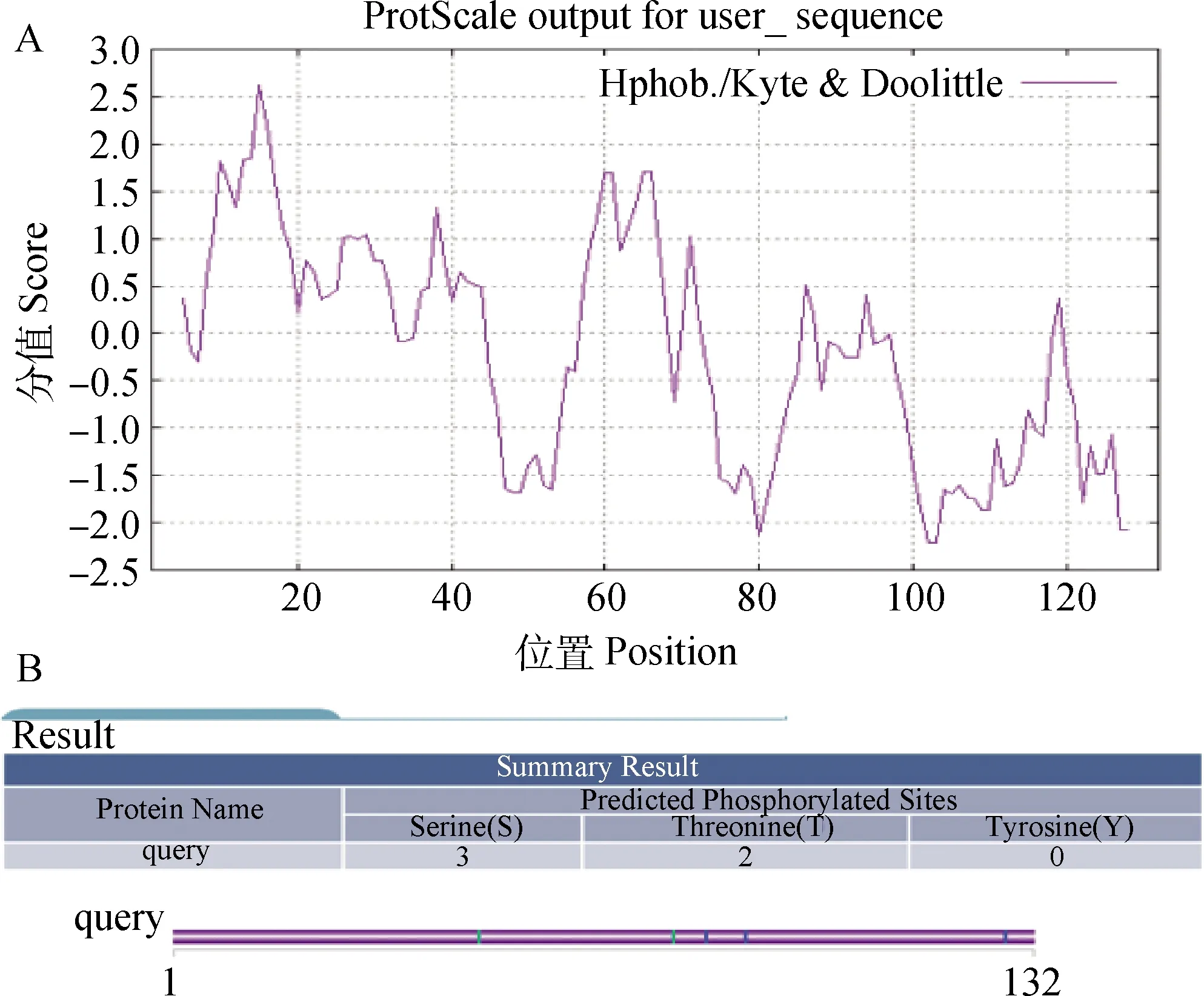
A. 疏水性分析;B. 磷酸化分析 A. Hydrophobicity analysis; B. Phosphorylation analysis图3 绵羊Musclin蛋白疏水性和磷酸化位点分析Fig.3 Hydrophobicity and phosphorylation site analysis of the Musclin protein in sheep
细胞分化72 h时,与空载体组相比,Musclin过表达组的MusclinmRNA表达量升高了6 024.56倍(P<0.01); 与空载体+胰岛素组相比,Musclin过表达+胰岛素组的MusclinmRNA表达量升高了6 273.20倍 (P<0.01);与Musclin过表达组相比,Musclin过表达+胰岛素组的MusclinmRNA表达量升高了3.26倍(P<0.01,图5)。
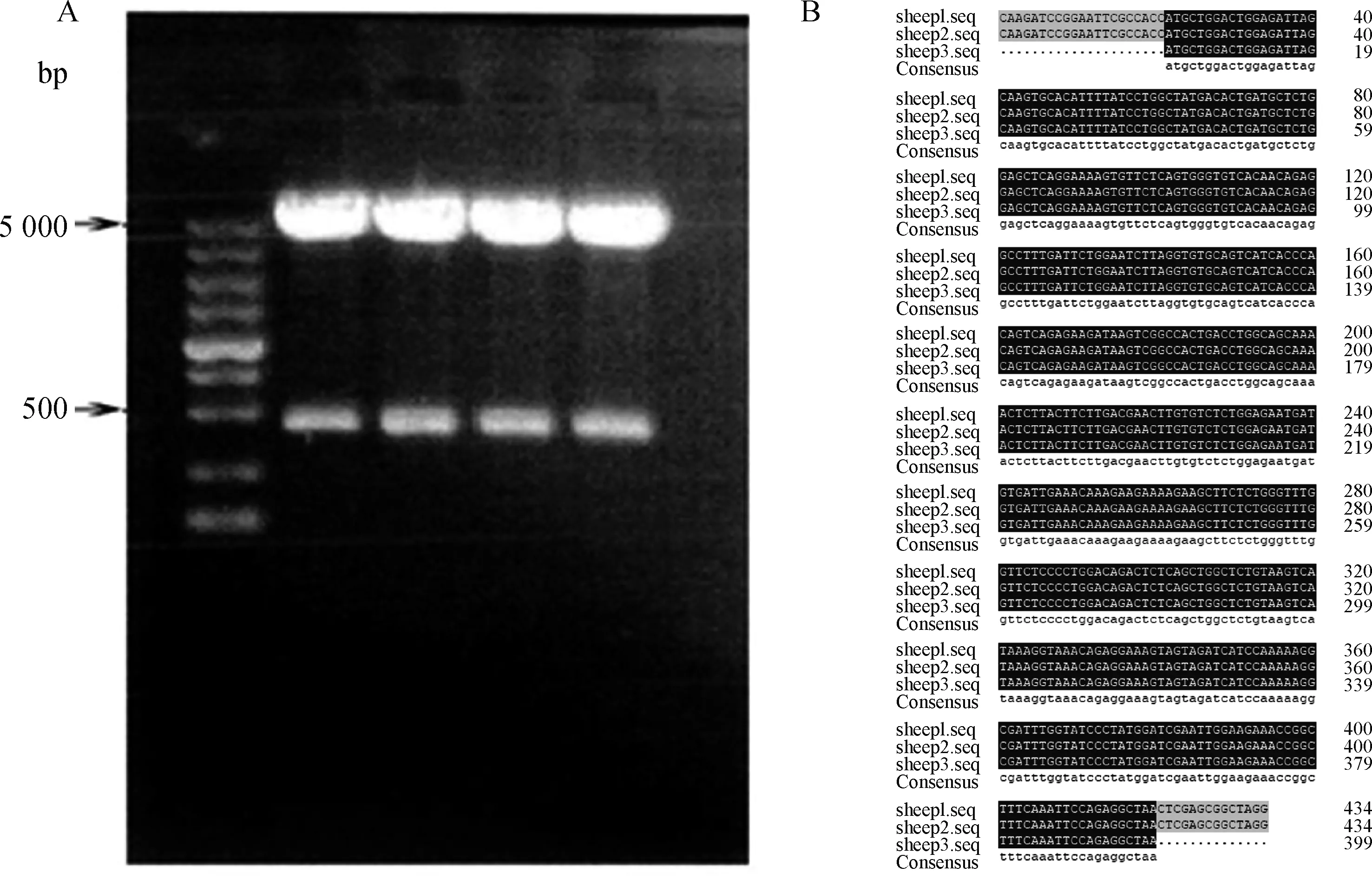
A. 酶切结果; B. DNAMAN比对结果,sheep1为质粒测序,sheep2为gBlocks合成序列,sheep3为NCBI中序列 A.Results of digestion; B. Comparison results by DNAMAN, sheep1 is the sequence of plasmid, sheep2 is the gBlocks synthetic sequence, sheep3 is the sequence in NCBI图4 pcDNA3.1-Musclin重组质粒酶切鉴定和DNAMAN比对结果Fig.4 Results of digestion and DNAMAN comparison for the recombinant plasmid pcDNA3.1-Musclin
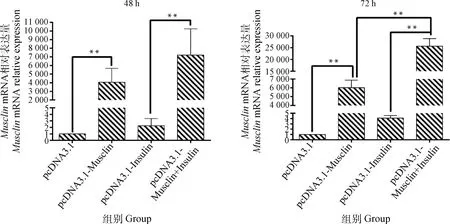
*. P<0.05,**. P<0.01,下同 *. P<0.05, **. P<0.01, the same as below图5 不同转染组绵羊Musclin mRNA表达的结果Fig.5 Expression result of sheep Musclin mRNA in different transfection groups
2.4 绵羊Musclin基因过表达及联合胰岛素对绵羊肌细胞培养液中葡萄糖含量的影响
细胞分化48 h时,与空载体组相比,Musclin过表达组的葡萄糖含量升高了0.14倍(P<0.05);与空载体+胰岛素组相比,Musclin过表达+胰岛素组的葡萄糖含量升高了0.11倍(P<0.05);与空载体组相比,空载体+胰岛素组葡萄糖含量降低了0.08倍(P<0.05,图6A)。
细胞分化72 h时,与空载体组相比,Musclin过表达组的葡萄糖含量升高了0.17倍(P<0.05);与空载体+胰岛素组相比,Musclin过表达+胰岛素组的葡萄糖含量升高了0.19倍(P<0.05,图6A)。
2.5 绵羊Musclin基因过表达及联合胰岛素对绵羊肌细胞中糖原含量的影响
细胞分化48 h时,与空载体+胰岛素组相比,Musclin过表达组+胰岛素组的糖原含量下降了0.15倍(P<0.05);与空载体组相比,空载体+胰岛素组糖原含量升高了0.31倍(P<0.01,图6B)。
细胞分化72 h后,与空载体组相比,Musclin过表达组糖原含量降低了0.12倍(P<0.05);与空载体+胰岛素组相比,Musclin过表达+胰岛素组的糖原含量下降了0.17倍(P<0.01);与空载体组相比,空载体+胰岛素组糖原含量升高了0.33倍(P<0.01); 与Musclin过表达组相比,Musclin过表达组+胰岛素组糖原含量升高了0.25倍(P<0.01,图6B)。
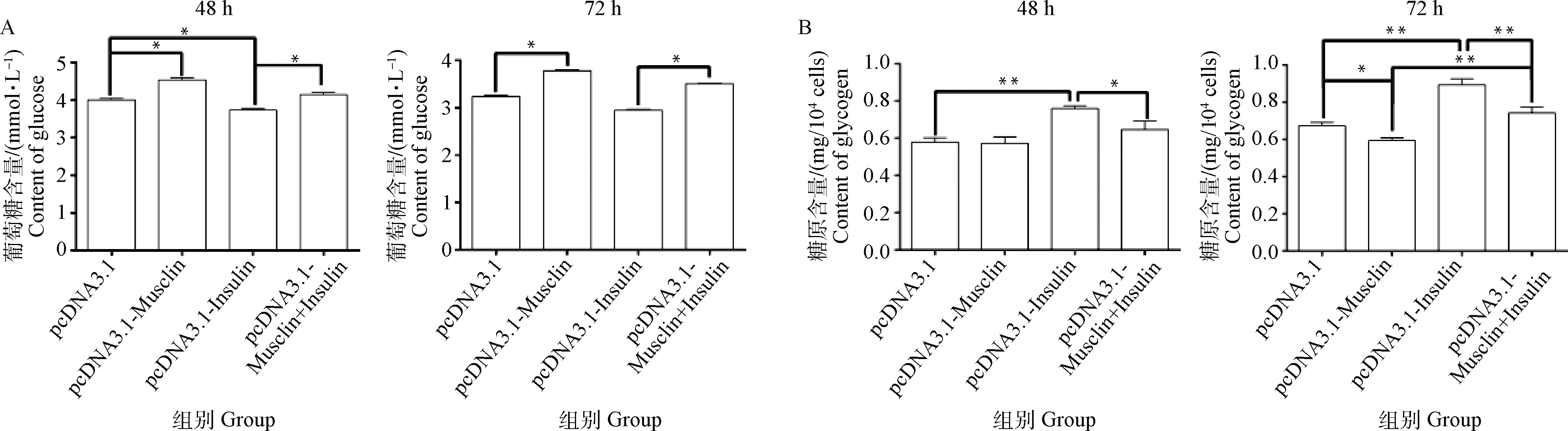
A. 葡萄糖含量;B. 糖原含量 A. Content of glucose; B. Content of glycogen图6 不同转染组培养液葡萄糖和细胞糖原含量Fig.6 Contents of glucose in media and glycogen in cells for different transfection groups
2.6 绵羊Musclin基因过表达及联合胰岛素对绵羊肌细胞糖代谢相关基因mRNA表达的影响
细胞分化48 h后,与空载体组相比,Musclin过表达组GLUT4 mRNA表达量降低了0.22倍(P<0.05);与空载体+胰岛素组相比,Musclin过表达+胰岛素组GLUT4 mRNA表达量显著降低了0.38倍(P<0.05);与空载体组相比,空载体+胰岛素组GLUT4和IRS1mRNA表达量分别升高了0.38和0.23倍(P<0.05,图7)。
细胞分化72 h后,与空载体组相比,Musclin过表达组IRS1表达量降低了0.27倍(P<0.01),AKT2和GLUT4 mRNA表达量降低了0.23和0.32倍(P<0.05),AKT1 mRNA表达量降低了0.31倍(P<0.01),GSK3β mRNA表达量升高了1.00倍(P<0.01);与空载体+胰岛素组相比,Musclin过表达+胰岛素组的IRS1、AKT1、AKT2、GLUT1和GLUT4 mRNA表达量分别降低了0.46、0.39、0.55、0.64和0.31倍(P<0.01);与空载体组相比,空载体+胰岛素组IRS1、AKT1、AKT2和GLUT4 mRNA表达量分别升高了0.57、0.36、0.76和1.52倍(P<0.01),GLUT1 mRNA升高了0.27倍(P<0.05);与Musclin过表达组相比,Musclin过表达+胰岛素组的GLUT1和GSK3β mRNA表达量分别下降0.48倍和0.62倍(P<0.01),GLUT4 mRNA表达量升高了1.56倍(P<0.01,图7)。
3 讨 论
绵羊Musclin蛋白二级和三级结构均以α螺旋和无规则卷曲结构为主。尽管绵羊Musclin的氨基酸序列与其他物种存在差异(结果未显示),但根据绵羊Musclin蛋白信号肽分析和亚细胞定位结果可以推测出绵羊Musclin蛋白与小鼠的类似,也属于分泌蛋白,这与Nishizawa等[1]将Musclin定义为骨骼肌分泌因子的结果相符。
Nishizawa等[1]研究表明,高胰岛素处理的小鼠肌肉中MusclinmRNA水平升高。本课题组前期研究结果表明,较高浓度胰岛素能够促进绵羊Musclin启动子活性的增加[26],说明胰岛素对Musclin的转录表达可以起到上调作用。然而,本试验中细胞分化48或72 h时,胰岛素+空载体组相比于空载体组的MusclinmRNA水平有增加趋势但并没有呈现显著性差异。而细胞分化72 h时,Musclin过表达+胰岛素组相比于Musclin过表达组的MusclinmRNA水平则显著增加。这可能是由于存在某种能够抑制胰岛素对Musclin调控作用的机制。如,FoxO1是Musclin基因的上游转录因子,在小鼠成肌细胞中研究发现,FoxO1-3A元件能调控Musclin基因的转录,抑制胰岛素诱导的MusclinmRNA表达上调作用[27-28]。而当细胞Musclin
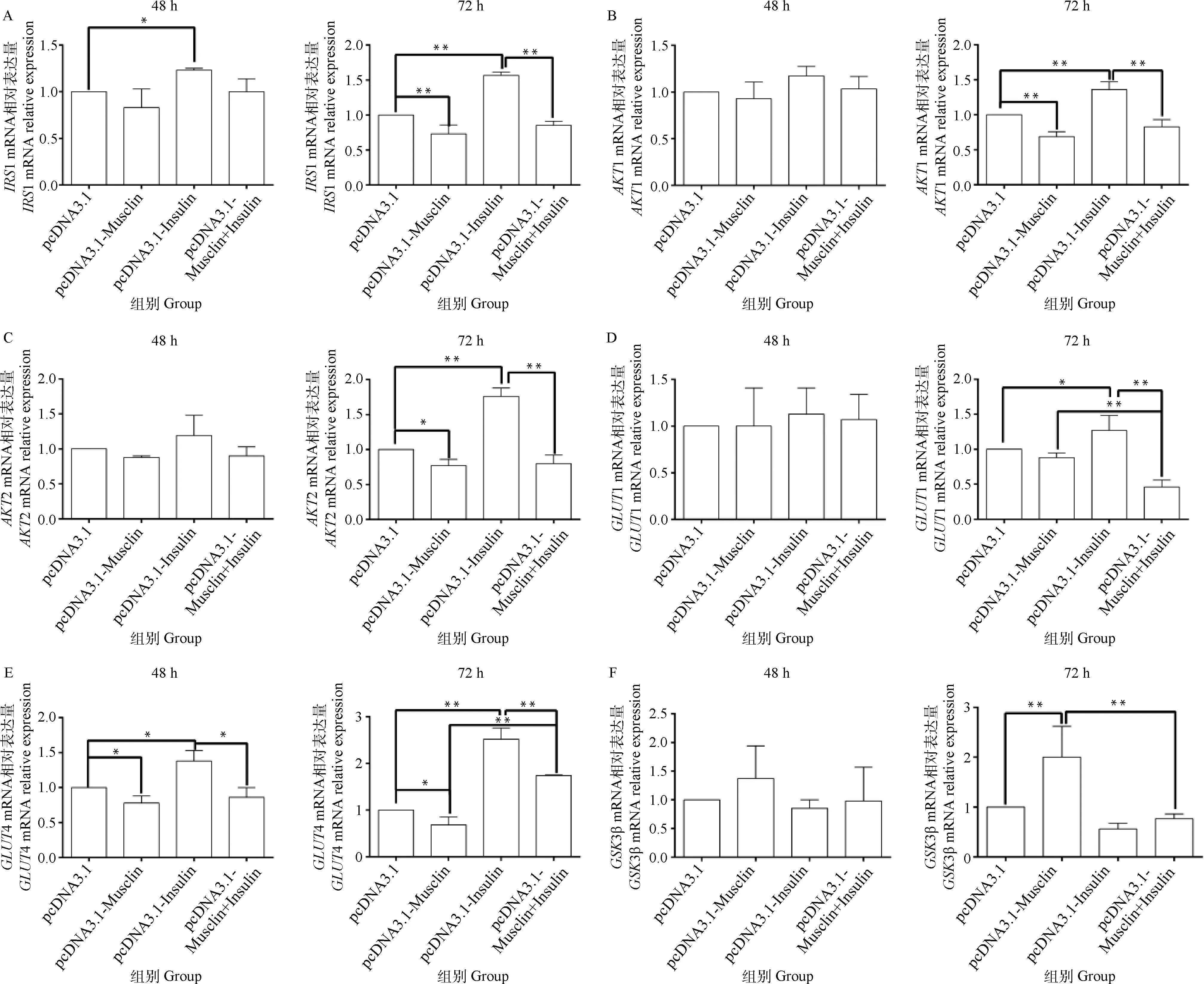
图7 不同转染组相关基因mRNA表达结果Fig.7 Expression results of the related genes mRNA in different transfection groups
处于高水平时则会减弱这种抑制作用,从而表现出胰岛素促进Musclin表达的作用。胰岛素通过上调Musclin表达而控制自身的作用,以相互协调而维持糖稳态[29]。当然,胰岛素和Musclin之间精确的调控关系和机制与复杂的糖代谢有关,尚需要更多的系统研究。
本试验中,无论Musclin过表达组相比空载体组,还是Musclin过表达+胰岛素组相比空载体+胰岛素组,培养液中的葡萄糖含量明显升高,细胞糖原含量则显著降低。推测,绵羊Musclin可能通过抑制肌细胞摄取葡萄糖和糖原合成或者加快糖原分解而影响糖代谢并减弱胰岛素功能。这与研究人员发现大鼠[2,14]和人[21]Musclin可通过紊乱葡萄糖代谢,导致胰岛素抵抗的发生相符。
IRS1受到胰岛素刺激后可以酪氨酸化,激活下游PI3K-AKT信号通路[30-31],从而促进胰岛素刺激的葡萄糖摄取和储存。活化的AKT,一方面会促进GLUT的表达或转移,促进细胞对葡萄糖的摄取[32-36],另一方面使GSK3β N端的Ser残基磷酸化而变成无活性形式,促进糖原的合成[37-38]。为了进一步研究绵羊Musclin影响糖代谢和胰岛素功能的分子机制,本研究对糖代谢相关基因的表达进行了检测,发现绵羊成肌细胞分化48 h时,Musclin过表达对IRS1、AKT1、AKT2、GLUT1、GSK3β mRNA的表达量没有产生明显影响,只有GLUT4 mRNA的表达变化显著;而在分化72 h时,绵羊Musclin过表达则显著抑制了正常或胰岛素处理的成肌细胞中IRS1、AKT1、AKT2、GLUT1、GLUT4基因mRNA表达,上调了GSK3β mRNA的表达。说明绵羊Musclin对糖代谢相关基因表达的调控作用与成肌细胞的分化程度有关。诱导分化48 h时,分化程度不高,绵羊Musclin主要通过GLUT4基因的表达上调而发挥糖代谢调节作用,这与本课题组先前在未分化小鼠C2C12的研究结果一致[39]。而当成肌细胞分化到一定程度时,绵羊Musclin可能通过下调糖代谢通路中IRS1、AKT1、AKT2、GLUT1、GLUT4基因的表达,抑制细胞对葡萄糖的摄取,同时通过上调GSK3β基因的表达,抑制糖原合成,从而影响糖代谢并减弱细胞对胰岛素的生物反应。具体的调控模式尚需要利用各基因过表达或相应蛋白增补、基因沉默或蛋白抑制剂增补等各种措施,在蛋白水平进一步深入研究。
4 结 论
绵羊Musclin过表达可以影响肌细胞糖代谢并减弱胰岛素的作用。绵羊Musclin和胰岛素通过相互协调以维持糖代谢稳态,其作用机制涉及到IRS1、AKT1、AKT2、GLUT1、GLUT4和GSK3β相关基因表达的变化,并且与绵羊成肌细胞的分化状态有关。本试验为进一步揭示Musclin对绵羊肌肉发育的作用机制奠定了基础。
猜你喜欢
广西糖业(2022年5期)2022-11-24
肝博士(2021年1期)2021-03-29
中国甜菜糖业(2020年1期)2020-12-07
意林(儿童绘本)(2019年6期)2019-08-05
小学生必读(低年级版)(2019年12期)2019-04-20
江苏卫生保健(2018年7期)2018-07-31
幼儿画刊(2018年7期)2018-07-24
成都体育学院学报(2017年1期)2017-02-21
中国乳业(2016年4期)2016-11-07
数学大王·低年级(2016年8期)2016-05-14
