
开放水体脱氮过程及其影响因素研究进展
2021-03-30 09:36杨正健魏辰宇刘德富纪道斌王从锋
水利学报 2021年2期
杨正健,魏辰宇,刘德富,纪道斌,马 骏,王从锋
(1.三峡大学 三峡水库生态系统湖北省野外科学观测研究站,湖北 宜昌 443002;2.湖北工业大学 河湖生态修复与藻类利用湖北省重点实验室,湖北 武汉 430068)
1 研究背景
据统计,2017年中国废水(不含农村面源污染)总氮排放量为216.46万t,总磷排放量为11.84万t,说明中国氮、磷污染问题仍然非常严峻[1]。过量的氮、磷导致的湖库水体富营养化及藻类水华问题,已对居民饮水安全和淡水生态安全构成严重威胁[2]。在海洋中,近年来多次报道的“海洋死亡地带(Dead zone/Hypoxic area)”问题已对区域海洋生态系统造成严重破坏[3],其主要原因也是河流入海氮、磷污染过量[4]。因此,如何有效拦截和去除流域内的氮、磷污染已成为维护全球生态系统平衡的关键,也是当前国际研究的热点和难点。有关研究表明,入海氮、磷总量实质只占陆域排放总量的20%~30%,说明陆域本身是除氮脱磷的主要场所,其河网潜流带和湖库沉积物内的生化反应被认为起到了主要作用[5]。
中国已建各类水库近10万座[6],几乎遍布所有大小流域。因水库改变了河流水文规律,其物质循环较自然河流有很大差别[7],由此对流域氮、磷污染的削减作用目前尚未有明确结论。本文将系统分析近30年来开放水体脱氮机制及其影响因素的国内外研究成果,讨论水库在流域氮削减中的作用,以期为发掘水库的脱氮除污功能、深入认识水库的生态环境影响提供新的研究思路。
2 开放水体典型脱氮机制
开放水体是有别于管道和池罐等相对封闭的水体而言的,包括河流、水库、湖泊、海洋等自然水体,是地球水循环及其伴生过程发生的主要区域。脱氮(Nitrogen removal)是指流域内的有机或无机态氮最终转化为气态氮并进入大气的过程[8]。因常温下只有氮气(N2)属于惰性且无害气体,故生成氮气的脱氮过程被认为是最有效的氮负荷削减过程,成为目前国际上关注的重点[9]。本文所阐述的“开放水体脱氮”,主要是指发生在河流、水库、湖泊、海洋等水体中的沉积物、悬浮物的脱氮。在这些水体内,目前公认的典型脱氮机制主要包括:①厌氧反硝化(Denitrification)、②厌氧氨氧化(Anaerobic ammonium oxidation)、③好氧反硝化(Aerobic denitrification)、④厌氧甲烷氧化(Anaerobic methane oxidation)等4个过程,如图1所示,其具体反应说明如下。
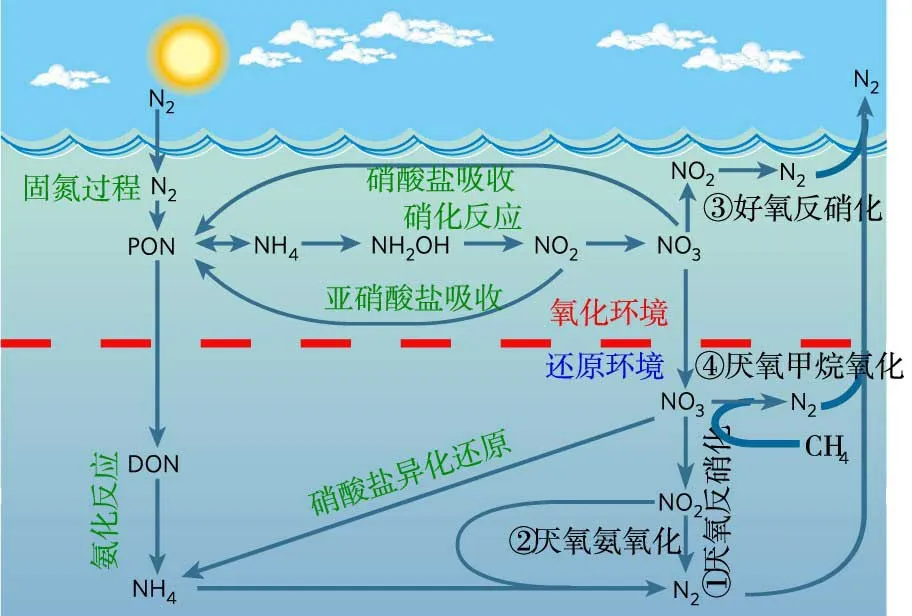
图1 开放水体氮循环及典型脱氮过程(基于Arrigo[9]成果修改)
厌氧反硝化[9]:在厌氧环境下,微生物将硝酸盐逐步转为亚硝酸盐(NO-2)、一氧化氮(NO)、氧化亚氮(N2O)并最终还原为氮气(N2)。
厌氧氨氧化[8]:在厌氧条件下,微生物以为电子受体,直接将氨氮(NH+4)氧化为氮气(N2)。
其中,厌氧反硝化过程一度被认为是自然界最主要的脱氮过程[9];后来逐步研究发现,厌氧氨氧化广泛存在于河流、湖泊和海洋沉积物中,其在海洋沉积物脱氮过程中所占的比例最多可超过50%[10]。
从上述过程来看,厌氧环境和NO-
3是脱氮发生的两个先决条件,因自然水体中NO-3条件容易满足,故溶解氧被认为是指示脱氮能否发生的重要指标,并将其浓度低于0.5~2.0 mg/L作为氧化与还原环境的判定条件[11]。Robertson 发现在溶解氧为5.0 mg/L 的环境中,某些细菌也能将NO-3还原为氮气[12],原因是部分微生物能产生某种周质硝酸盐还原酶(P-Nar),可在有氧环境中促进硝酸盐还原[13],即为自然水体脱氮的第三类过程③好氧反硝化。后来,又有人发现有些微生物在厌氧环境下能够利用甲烷(CH4)将硝酸盐还原为氮气,即④厌氧甲烷氧化[14],因该过程既可消耗甲烷,又能去除硝酸盐,还不产生亚硝酸盐,可在削减氮污染的同时去除温室气体,故在近年来备受国际学者关注。
3 潜流带与沉积物主要脱氮过程
目前研究认为陆域排放的氮、磷污染有70%~80%是在河流流域内被去除的,且主要发生在河网潜流带和湖库沉积物中[5]。“潜流带(Hyporheic zone)”是指位于溪流或河流河床之下并延伸至河溪边岸带的水分饱和的沉积物层,包括侧向潜流带和垂向潜流带[15],如图2所示。因地表水和地下水在此相互混合并发生物质和能量交换,故潜流带也被称为地表水-地下水交换带[16]。研究表明,潜流带伸展体积是地表水体的数倍乃至数百倍[17],水体自初入河流到最后移出,几乎全部与潜流带发生过交换[18]。因此,潜流带是河流中水量交换、溶质迁移、新陈代谢和污染物储存的主要场所,被誉为河流的肝脏[19];同时还是多数河流生物的栖息地,具有重要的生态功能[20]。
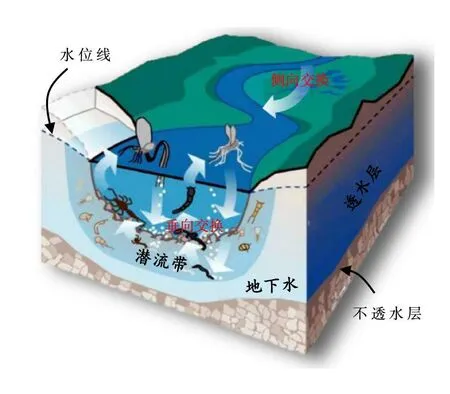
图2 流域河网潜流及其物质交换带示意[26]
潜流带内氮素运移转化过程一直是国际研究的热点,甚至有人认为河流中50%以上的脱氮过程都发生在潜流带中[21]。研究表明,潜流带脱氮过程主要取决于温度、硝态氮浓度、有机碳浓度以及沉积物层的非均质性,其中有机碳浓度是关键因素[22]。潜流带水体交换可通过改变上述条件而影响脱氮过程。例如,地表水向下运动为潜流带提供了溶氧,而地下水则为潜流带提供了厌氧环境,好养与厌氧交替出现加速了潜流带有机质的快速分解[23]。对氮素而言,在溶氧充足的地表水中一般以硝酸盐的形式存在,而在厌氧的地下水中则以氨氮为主要形态[19]。当地表水携带硝酸盐进入潜流带后,经沉积物颗粒的吸附、反硝化及厌氧氨氧化[22],硝态氮得以还原为氮气,或被潜流带表层的藻类、植物利用而被移出水体[24]。潜流带水流交换还可通过控制地表水与地下水的混合比例和其在潜流带中的反应时间长短,进而影响潜流带中有氧和厌氧环境及温度的分布[25]。
对于湖库或海洋,水体脱氮则被认为主要发生在一层很薄的沉积物-上覆水界面上[27]。因湖库或海洋等沉积物较厚、颗粒粒径较小,且水体垂向运动相对较弱,在沉积物内难以形成明显的水流通道,含有溶氧和硝酸盐的上覆水很难直接进入沉积物内部,只能在沉积物表层以扩散为主的方式进行交换[28]。因此,虽然沉积物内部富含有机质和微生物并处于厌氧环境,但因硝酸盐浓度极低而无法发生脱氮反应,但上覆水扩散进入的溶解氧和硝酸盐刚好在沉积物-上覆水界面与沉积物中的氨氮相遇,溶解氧很快消耗并形成厌氧环境[29]。大量研究已在湖库、平原河流或海洋沉积物表层中找到了反硝化、厌氧氨氧化以及厌氧甲烷氧化的证据[30]。
4 水体及其悬浮颗粒脱氮过程
溶解氧极小层(Oxygen minimum zone,OMZ)是目前已知的开放水体脱氮最可能发生的区域[31]。OMZ是指分层水体中存在的溶解氧消耗远大于补给的某层水体,深水水库及海洋中时有发生[32]。因水温和盐度等因素的影响会使分层水体的某层密度突然增大,阻断了上部溶氧向下传输;同时,上层真光层产生的藻类等有机质也因该层水体密度骤增而无法下沉。高浓度有机质和低溶氧补给,很容易导致该层内溶氧浓度骤降,OMZ随即形成[31,33]。例如,Thamdrup等[34]发现虽然OMZ上层水体溶氧可超过250 μmol/L,但在OMZ 层中心区域溶氧最低可为10 nmol/L;而当溶解氧低于25 μmol/L 时硝酸盐还原开始发生,当溶解氧低于20 μmol/L 时厌氧氨氧化开始发生[35]。同时,因OMZ 层内有机碳充足,厌氧氨氧化在该层内可不受碳源限制而成为主要的脱氮过程,厌氧反硝化发生概率反而较小[36]。因此,OMZ层刚好形成了一个非常适合脱氮的环境,有人估算在海洋OMZ内,每年以厌氧氨氧化而产生的氮气总量约为0.8~1.5亿t[37],不到海洋总体积1%的OMZ 层贡献了整个海洋脱氮总量的30%~50%,意义非常重大[38]。
近年来,越来越多的学者发现富氧水体中的悬浮颗粒也可强化水体脱氮,例如,Xia等[39]通过实验证实在富氧水体中加入悬沙能够强化水体脱氮;Bianchi等[40]在富氧水中的悬浮颗粒物内检测到了厌氧微生物和厌氧甲烷氧化产物。除悬沙外,浮游植物(藻类)等颗粒也被证明可促进水体脱氮,例如,Eriksson 等[41]发现加入浮游植物的水体比未加入的脱氮效率要高;Naqvi等[42]研究发现水体浮游植物的存在可诱发水体厌氧甲烷氧化反应。研究表明,如果考虑海洋藻类颗粒对脱氮的影响,每年海洋脱氮总量估算将要增加一倍以上[40]。
相较于潜流带、沉积物和OMZ层等特定的脱氮区域,颗粒物因不具备稳定的好氧、厌氧环境而无法用经典脱氮机制进行解释,加之直接观测数据的匮乏,一度成为科学争论的焦点。理论分析认为,悬浮藻类颗粒孔隙率可达99%[43],其内部有足够的空间供微生物、气体和营养物质富集[44]。Bianchi等[40]指出悬浮藻类颗粒内部可能发生固氮、脱氮、脱硫等多种反应;还有证据表明只有在最严格厌氧环境才可产生的甲烷也可在悬浮藻类颗粒内产生[45],这说明悬浮颗粒物内部结构与生物地球化学过程可能非常复杂。Klawonn 等[46]采用15N示踪方法证实悬浮藻类颗粒内确实存在复杂的生物地球化学过程,并给出了一个内外物质交换与脱氮过程的理论模式:①悬浮藻类颗粒内壁区域有机氮迅速矿化形成氨氮,其一小部分消耗溶氧并转化成硝酸盐,使得颗粒内部核心区处于厌氧环境;②扩散进入的一部分硝酸盐在厌氧环境下通过反硝化和厌氧氨氧化还原成氮气,另一部分则异化还原成氨;③颗粒核心区因溶氧极低而形成绝对厌氧环境,可能发生脱硫反应和产甲烷反应;④在悬浮颗粒内形成了溶氧和硝酸盐等氧化剂自外向内逐渐降低,而氨氮、氮气、甲烷、硫化氢等还原产物逐渐升高的梯度分布模式,如图3所示。
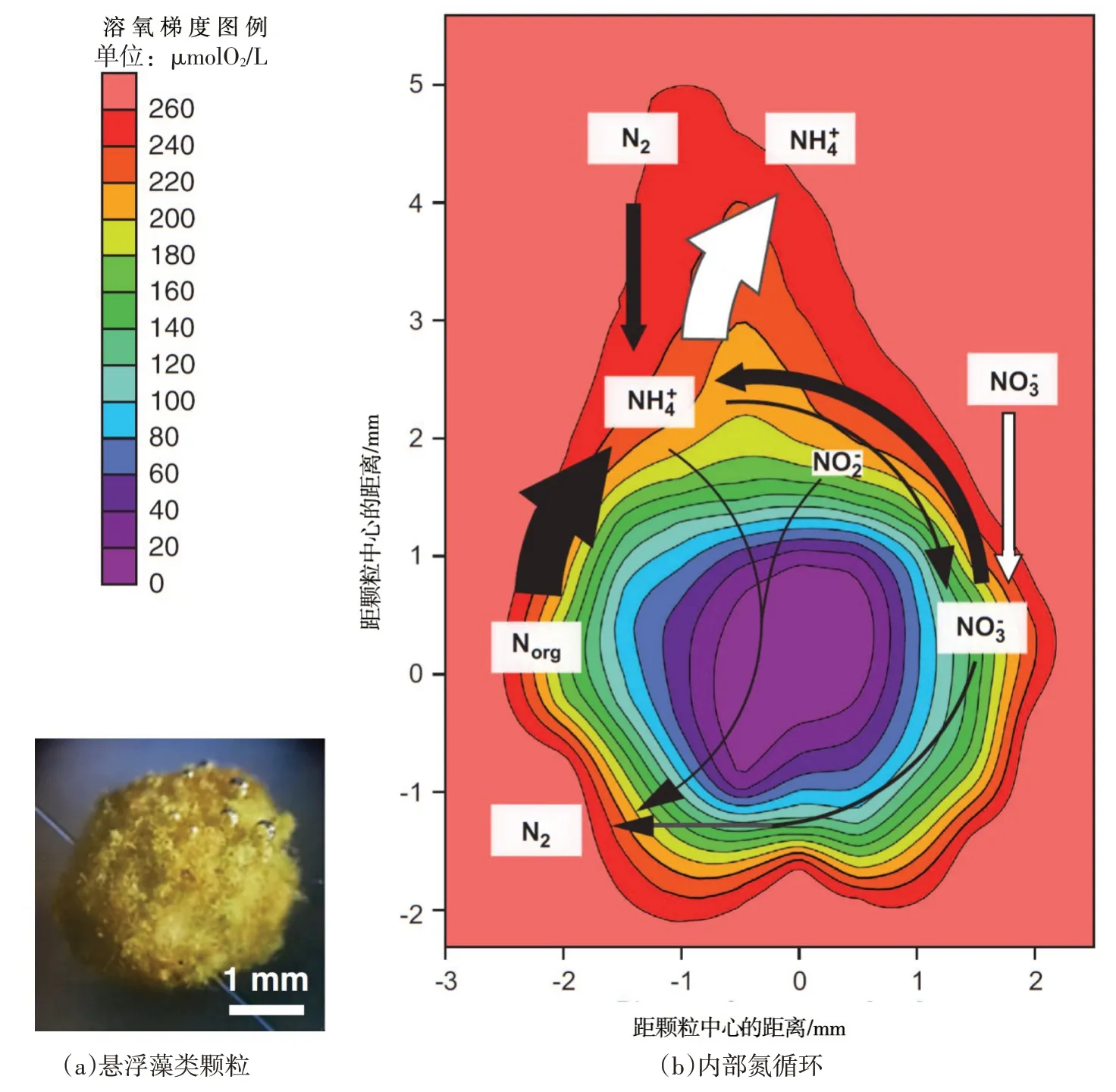
图3 悬浮藻类颗粒及其内部氮循环示意[46]
5 开放水体脱氮影响因素
决定开放水体脱氮过程及效率的主要因素主要包括溶解氧(DO)、碳氮比、硝酸盐浓度、温度和pH值。研究表明,微生物作用下的生物地球化学反应过程应该满足“氧化还原分级(Redox tower)”原理[47],即从绝对氧化到绝对还原环境,依次将发生氧化反应(消耗氧气)、脱氮反应(硝酸盐被还原为氮气)、发酵(产生乙酸)、金属氧化物还原反应(铁、锰等还原)、脱硫反应(硫氧化物还原为硫化氢)和产甲烷反应,因此,溶解氧被认为是决定脱氮是否发生的关键环境因子。一般认为,溶解氧在一定范围内脱氮效率不受影响,但当其下降到某一阈值时反硝化脱氮活性急剧上升[48]。虽不同种属的厌氧反硝化菌具有不同的耐氧阈值,但大部分反硝化细菌在溶解氧低于0.50 mg/L时才产生反应[11]。
因大多数的具有脱氮功能的微生物均为异养菌,故碳氮比很大程度上决定了微生物的脱氮速率。例如,Joo等[49]研究指出碳氮比越高越有利于厌氧反硝化菌的生物脱氮;而Kim等[50]又认为C/N为8 时反硝化活性最高;Bernat等[51]还发现细菌可以利用某些细胞内碳作为碳源进行反硝化,为低碳氮比污废水的生物处理提供了一种新的途径。
硝酸盐作为脱氮的基础物质,其浓度也与脱氮效率有一定关系。Hasegawa 等[52]指出硝酸盐浓度与反硝化速率成正比;诸多研究也表明反硝化速率在硝酸盐较低的环境中受到抑制,并随着硝酸盐的增加而增加[53]。
温度通过影响微生物酶的活性来影响微生物的代谢速率,Herrman 等[54]指出在沉积物环境中,25℃下的反硝化速率是8℃下的2倍;王弘宇等[55]发现一般厌氧反硝化菌在25~35℃的温度范围内具有很高的脱氮效率。
pH值主要通过引起细胞膜上的电荷的变化来决定微生物生长进而影响脱氮速率。一般中性及偏碱性有利于反硝化的进行,在酸性和强碱条件下菌株生长状况差[56]。此外,某些金属离子(如铁、镁、钙等)也可以影响细菌的脱氮活性[57]。
颗粒大小和水动力条件能够相互作用并影响悬浮颗粒的脱氮效率。颗粒大小可影响悬浮颗粒与周围水体的物质交换通量,进而决定颗粒内溶解氧分布梯度和脱氮过程[58]。已有研究表明,在高溶氧环境下,只有在直径大于1 mm的悬浮颗粒内才会出现厌氧环境而发生脱氮;当溶氧低于25 μmol/L时,该直径阈值可为0.1 mm[46];此外,不同颗粒大小还可通过影响颗粒的上浮或下沉速度而改变其在水中的滞留时间来决定脱氮效率[59]。水动力条件虽不能直接影响悬浮颗粒的脱氮过程,但其在决定悬浮颗粒大小、形态、内部物质梯度分布上能起到关键作用。例如,目前备受关注的藻类絮凝体(Algae aggregation)、海洋雪(Marine snow)等都是在静止分层水体中产生的,而在垂向混合较强的水体中一般不会形成较大的悬浮颗粒[60]。还有研究表明,即使水体环境相同,不同紊流下悬浮颗粒物内部溶氧梯度分布也存在显著差异[61]。此外,水体微动力结构还可通过改变悬浮颗粒的运动、聚集和漂浮特征来影响其脱氮过程[62]。
6 水库对流域脱氮贡献的研究展望
河流筑坝形成水库可引起河流水动力的改变,进而改变河流原有的物质场、能量场、化学场和生物场[7],必然对流域脱氮过程产生显著影响。以三峡水库为例,蓄水发电后,水库干流与支流库湾呈现出完全不同的水体类型和水环境状态:干流流速较大,水体垂向掺混较强,悬浮颗粒以外源泥沙和有机碎屑为主[63];支流库湾却因分层异重流的影响,呈现出时空差异显著的特殊分层状态,并由此诱发不同种类的藻类水华现象[64],悬浮颗粒以藻类颗粒为主。对比建库前后入库、出库总氮变化规律发现,在2003年以前,三峡库区污水年排放量与出库(三峡坝址以下南津关)水体总氮浓度变化趋势基本一致,呈正相关(图4(a));三峡水库蓄水后,虽然国家在三峡库区修建了大量污水处理厂,但因现行标准下污水厂尾水氮、磷含量较地表水仍然很高;加之库区处于经济社会加速发展时期,污水排放总量持续增加,理论上三峡水库出库水体总氮浓度也应该逐渐升高;但是,实际监测发现,三峡库区污水年排放量确实在逐年升高,但出库水体总氮浓度却基本维持不变(图4(b)),这说明三峡水库显著降低了向下游输送的氮污染负荷,但相关削减机制尚需进一步探究。

图4 三峡库区污水年排放量和出库水体总氮浓度变化
此外,世界海洋“死亡地带”[3]分布也可间接证实中国水库可能会影响我国入海氮负荷。目前海洋“死亡地带”已增至500余处,总面积达到450万km2,并还在快速蔓延[4]。虽然在中国部分海域已有“死亡地带”的报道[65],但其发生次数和面积较其它海域明显要少,说明中国河流入海营养盐通量可能还未达到“死亡地带”频发的水平,这与中国近年来污染排放总量较高的现实并不一致[1]。中国河流建有大小不一的水库,总数接近10 万座;水库因影响了河流水文过程,进而改变流域氮、磷循环[7],图4所示的结果与上述推测基本吻合,说明入海口氮、磷污染与水库建设可能存在一定关系,但水库建设后究竟能降低入海口多少氮通量,尚需进一步研究。
从理论分析来看,对沉积物及潜流带而言,水库化延长了河流水停留时间,促进有机质沉积,且水体中的生物耗氧过程造成溶解氧的降低;水库通过对泥沙的异化沉积作用,将原河流潜流带的泵吸交换和冲淤交换逐渐转化为湖泊型生物交换和毛细渗透,并将潜流带物质补给模式由水平输运为主型转化为以垂向沉积和扩散为主型[66],如图5所示,这些改变都有利于增大沉积物厌氧区域并促进沉积物的脱氮。对于水体而言,水库造成的水流减缓、水体滞留时间增长、水体垂向成层、浮游植物增多甚至水华发生等都可能影响水体脱氮过程,进而对流域氮、磷循环的产生影响[7]。
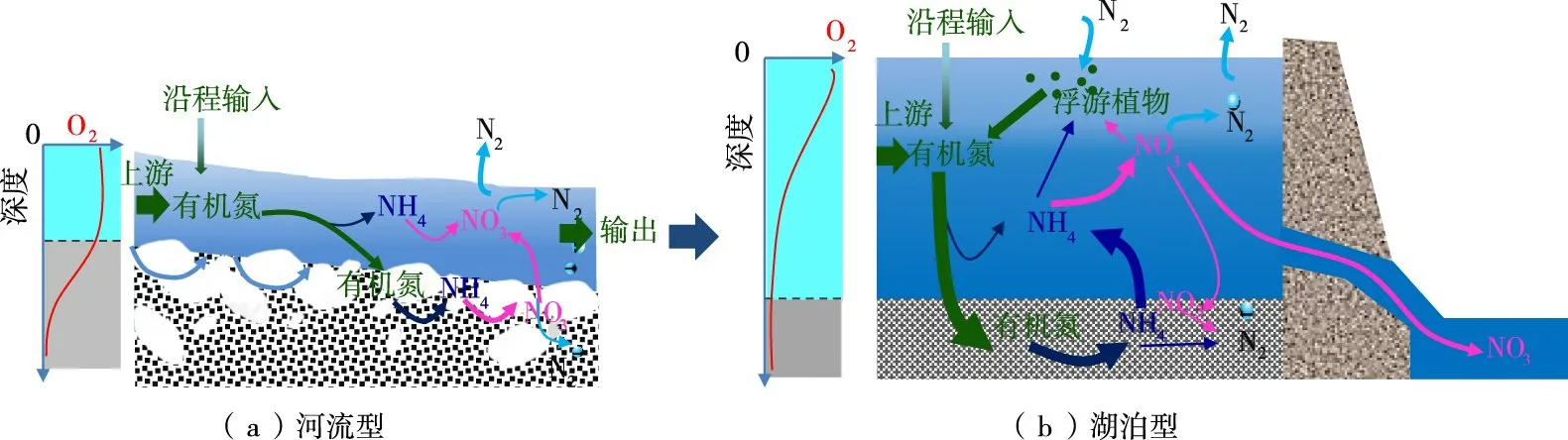
图5 河流型与湖泊型氮循环过程差异示意
因上述有关水库强化水体脱氮机制的理论分析目前尚未有直接证据,为回答“较自然河流,水库建设究竟是强化了脱氮作用还是降低了环境自净力?”这一问题,建议开展如下方面的研究工作。
(1)出入库氮形态持续观测及氮负荷平衡计算。选择大、中、小等典型水库,在入库、出库及库内设置关键性控制断面,对水体、沉积物和大气沉降中的总氮、无机氮、有机氮持续跟踪监测;计算各断面的不同形态氮输出通量和沉积物氮储存量;开展水库氮负荷平衡计算,确定水库氮的输入、输出和滞留等总量。
(2)水库不同载体脱氮机制原位研究方法构建。因受大气高浓度氮的影响,以“氮气”为脱氮直接证据的脱氮机制原位研究非常困难;加之水库的水动力条件和地球化学过程非常复杂,特别是大型水库水体较深,沉积物和深层水体的溶解态、气态氮研究就更为困难;水-气界面氮气释放通量也因受到大气影响而难以直接观测。因此,构建水库沉积物、水体、悬浮物及水-气界面的脱氮机制及界面交换过程研究方法对于研究“水库脱氮”至关重要。
(3)水库脱氮机制及氮移出通量研究。开展不同水库沉积物、水体、悬浮物等载体内的脱氮过程及影响因素研究,阐明水库不同载体内发生厌氧反硝化、好养反硝化、厌氧氨氧化、厌氧甲烷氧化或其它新的脱氮机制的环境条件,科学阐明“水库强化脱氮”的主要机制;同时以“氮气”作为直接证据,研究溶解态、气泡态氮在水库不同载体和界面间的交换、运移和逸出过程及通量。
(4)自然河流与水库脱氮效率对比研究。寻求可与水库开展对比研究的自然河流,开展河流沉积物、水体及悬浮物脱氮机制、效率及氮逸出通量研究,并与水库脱氮机制及效率进行对比,确定水库对流域氮循环的影响机制,并对比分析水库对氮的拦截作用和强化脱氮效率。
猜你喜欢
中老年保健(2022年3期)2022-11-21
科技视界(2021年21期)2021-08-24
海洋通报(2021年2期)2021-07-22
科学与信息化(2020年11期)2020-06-19
科学导报·科学工程与电力(2019年24期)2019-09-10
婚姻与家庭·性情读本(2018年8期)2018-11-09
水能经济(2018年7期)2018-07-12
理论观察(2016年11期)2017-04-17
中国果业信息(2015年12期)2015-01-24
中国新技术新产品(2013年20期)2013-11-16
