
苹果炭疽病抗性miRNA的筛选
2021-03-26 10:47张亚楠陈新慧
中国农学通报 2021年7期
张亚楠,陈新慧,张 杰
(农业应用新技术北京市重点实验室/植物生产国家级实验教学示范中心/北京农学院植物科学技术学院,北京102206)
0 引言
苹果炭疽病是苹果生产中的主要病害之一,其主要致病菌为苹果炭疽病菌(Colletotrichum gloeosporioides),以危害苹果叶片和果实为主,目前针对该病的防控措施主要为施用化学药剂,生产上多以波尔多液和多菌灵交替使用进行病害防治,具有一定预防效果,但需要喷施3~4次,不仅加重果园管理的工作量,也增加了生产成本,且频繁施用药剂不仅使种植环境恶化,更给食品安全带来极大的隐患[1]。苹果炭疽病因发病迅速、潜育期短的特征使苹果生产者头痛不已,病原菌一旦侵入寄主组织,几乎没有用药防治的时间,常带来减产甚至绝产的危害。目前,利用植物诱导抗病机制控制病害,能减轻生态环境污染,具有持效期长、抗病谱广等优点,是一种植物病害防治的新策略。
miRNA是一类内源的调节基因表达的小RNA,长度19~25个核苷酸(nt),对靶基因mRNA的作用主要取决于它与靶基因转录体序列互补的程度,主要有3种方式,第一种是通过碱基互补切割靶基因,第二种抑制靶基因的翻译但不完全互补结合,而是阻遏翻译但不影响mRNA稳定性,第三种为结合抑制,miRNA通过这些方式在真核基因表达调控中广泛存在并具有多样性[2]。miRNA能否正常表达,决定了植物是否正常生长发育,已有研究表明,miRNA广泛参与调节植物生长、器官建成和形态发育,转录因子的表达情况是其主要调控途径[3]。植物并非总是在正常适宜的环境中,在其整个生命周期中,会面临各种各样的变化,地理因素、气候条件,尤其是人类活动和工农业发展也造成了诸多环境问题和污染,而这些因素的变化超出了植物维持正常生命所能自动调节的范围,使其生长受限、病变甚至死亡。植物所遭受的胁迫通常为生物逆境(病毒、虫害、杂草等)和理化因素逆境(极端天气气候、离子辐射、化学性盐碱土危害、大气、水体、土壤污染、高温热害、低温冷害和冻害、淹涝灾害和干旱等)。逆境胁迫会诱使植物内的基因表达发生改变,从而引起形态结构变化和某些代谢变化,如水分代谢、光合作用、呼吸作用和物质代谢。而植物的生理适应是多方向的,主要包括渗透调节、植物激素和蛋白质的适应性变化。在胁迫条件下miRNA同样可以调控植物的生长发育,从而增强植物自身对逆境胁迫的适应能力,分为避逆性和抗逆性[4]。miRNA可以在相对较短的时间内对植物所遭受的外界胁迫做出快速响应,通常24 h内即有较明显变化,相应地调整植物相关形态等来应对,此前已报道很多miRNA参与调节植物抗病的分子机制研究[5],如受到引起番茄细菌性叶斑病(Pseudomonas syringaepv.tomato)的病原菌DC3000侵染时,抑制生长素信号,部分由miR393介导的,可能特别促进对强致病菌DC3000的抗性,但与小种特异性抗性无关[6]。在拟南芥中,miR398受到蔗糖的正调控,导致CSD1和CSD2的mRNA和蛋白质的积累减少,由于CSD酶调节ROS,miR398的一个作用可能是影响ROS信号的传导或衰减,在植株遭受氧化胁迫中起着至关重要的作用[7]。已有研究表明,Pst DC3000(avrRpml或avrRpt2)病原菌侵染模式植物拟南芥时,miRNA398通过改变CSD表达水平来提高植物应对生物胁迫威胁时的抗性[8]。植物miRNAs的一个子集(miR482/2118超家族)可以针对NB-LRR基因调节植物免疫,在番茄中miR482/2118通过NBS-LRR来抵抗番茄晚疫病菌的侵染[9-12]。而在马铃薯中,miR160通过靶向StARF10,影响防御和生长路径的平衡,这可能是马铃薯植物在疫霉病菌感染过程中调节局部和SAR防御反应的途径之一[13]。在水稻中Osa-miR398b通过多种超氧化物歧化酶促进H2O2的产生,从而提高水稻对稻瘟病菌的抗性[14]。同样,水稻中OsamiR1873通过LOC_Os05g01790微调水稻免疫生长平衡,阻断Osa-miR1873可以提高稻瘟病抗性,并且产量不受其显著影响[15]。
因为复杂的致病性,对果树抗病机制仍缺乏深入了解,miRNA调控果树抗病的相关分子机制尚不清楚,对抗病基因资源也缺乏全面的掌握和了解。本研究拟以威胁苹果属植物生长发育的重要病原菌胶胞炭疽为研究对象,测定病原菌侵染后苹果属植物中抗性相关miRNA的表达量变化,研究miRNA在抗炭疽病中所起到的作用,旨在揭示苹果属植物炭疽病抗性的分子调控机制提供新的思路,为苹果属植物的抗炭疽病寻找新的突破口。
1 材料与方法
1.1 试验材料
1.1.1 植物材料‘八棱脆’海棠和‘嘎啦’苹果组培苗由实验室继代培养保存。
1.1.2 炭疽病菌株 2019年10月10日在北京市顺义区木林镇资源圃采集发病植株,取样并进行病原菌分离与鉴定。胶胞炭疽病菌在PDA培养基(土豆200 g,葡萄糖20 g,琼脂20 g,煮软后8层纱布过滤定容至1000 mL,高压灭菌锅115℃20 min)上培养。制备胶胞炭疽病菌孢子悬浮液侵染‘八棱脆’海棠和‘嘎啦’苹果组培苗,4天后发病取样,在液氮中速冻,-80℃保存。
1.2 试验方法
1.2.1 胶胞炭疽孢子悬浮液制备 将纯化鉴定过的胶胞炭疽菌株接种于PDA平板上,在28℃恒温箱中培养7天,PDA平板上加入适量无菌水用无菌的玻璃涂布刮取表面菌丝,无菌3层纱布过滤制备分生孢子悬浮液。胶胞炭疽菌孢子悬浮液浓度为1×106个孢子/mL,无菌水为空白对照。
1.2.2 接种处理与取样 试验设‘八棱脆’(抗性品种)空白对照、‘八棱脆’接种处理、‘嘎啦’(感病品种)空白对照、‘嘎啦’接种4个处理,每个处理3瓶组培苗,试验设3组重复。用喷壶将106个孢子/mL胶胞炭疽孢子悬浮液均匀喷洒到整个幼苗上,直至滴水状态,喷施等量无菌水的组培苗作为空白对照。接种4天后发病,发病叶片取样后立即在液氮中冷冻并保存于-80℃冰箱。计算病情指数[式(1)],0级,无病斑;1级,病斑面积≤5%;3级,5%<病斑面积≤10%;5级,10%<病斑面积≤25%;7级,25%<病斑面积≤50%;9级,病斑面积>50%。

1.2.3 植物总RNA提取及反转miRNA采用复杂植物多糖多酚植物RNA快速提取试剂盒提取植物叶片总RNA,用全式金试剂反转miRNA。
1.2.4 实时荧光定量PCR按照SYBR染料法试剂说明书进行实时荧光定量PCR检测,miRNA以U6作为参照基因,两步法程序运行,所有样品取样均为3次生物学重复,3次技术重复,所用荧光引物见表1。

表1 试验中所用引物
1.2.5 相关miRNA靶基因预测 通过靶基因预测网站psRNATarget对相关miRNA的靶基因进行预测,通过NCBI比对,找到该基因的详细描述。
2 结果与分析
2.1 接种后苹果属植物发病情况调查
接种胶胞炭疽(图1)孢子悬浮液4天,调查其发病情况,抗性品种‘八棱脆’叶片只有少数病斑病害,病情指数15.4,病叶率27.6%;而感病品种‘嘎啦’则出现大量病斑且植株萎蔫,病情指数39.3,病叶率71.3%(图2)。
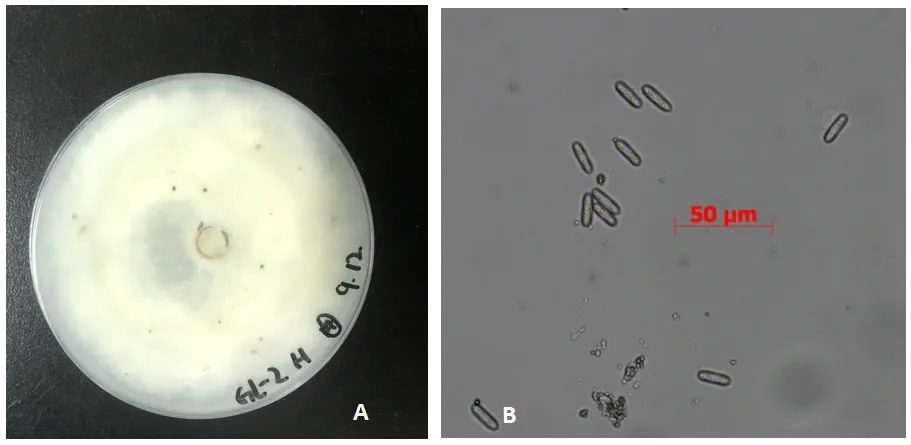
图1 菌株形态学培养特征

图2 苹果属植物接种胶胞炭疽菌后发病情况
2.2 抗病相关的miRNA筛选结果
根据实验室前期工作及相关文献报道,选出18个抗性相关miRNA候选待测,测定其在胶胞炭疽孢子悬浮液侵染后相关表达量变化。结果(图3)显示,在抗性品种‘ 八 棱 脆 ’中 miR390a、miR393a、miR396b/c/f、miR156a/p/t和miR171i下调表达,仅有miR172d和un3729上调表达,其余无显著变化。感病品种‘嘎啦’中miR390a、miR396b/c/f的表达量是上调的,miR160a、miR482b、miR156a/aa/ad/t和miR171i是下调的,其余无显著变化。
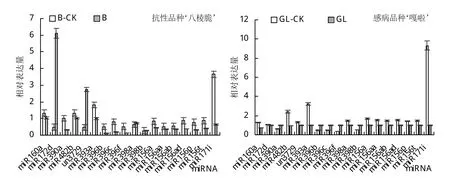
图3 苹果属植物接种胶胞炭疽菌后相关miRNA表达量
2.3 相关miRNA的靶基因预测
miRNA可导致靶mRNA的降解或者阻遏mRNA的翻译,从而抑制蛋白质的合成,达到调控基因的目的,因此靶基因的预测研究是探究miRNA功能的重要方式。通过靶基因预测网站psRNATarger(http://plantgrn.noble.org/psRNATarget/)对相关miRNA的靶基因进行预测,筛选并得到高度互补且假阳性低的靶基因(表2)。分析相关miRNA和其预测靶基因互补关系得到图4。miR390a靶基因多为LRR受体样丝氨酸/苏氨酸蛋白激酶,mi482b靶基因多为抗性蛋白,miR396b/c/f靶基因多为GRF家族和LRR受体样丝氨酸/苏氨酸蛋白激酶,均与抗性相关。

图4 相关miRNA及其靶基因互补性分析
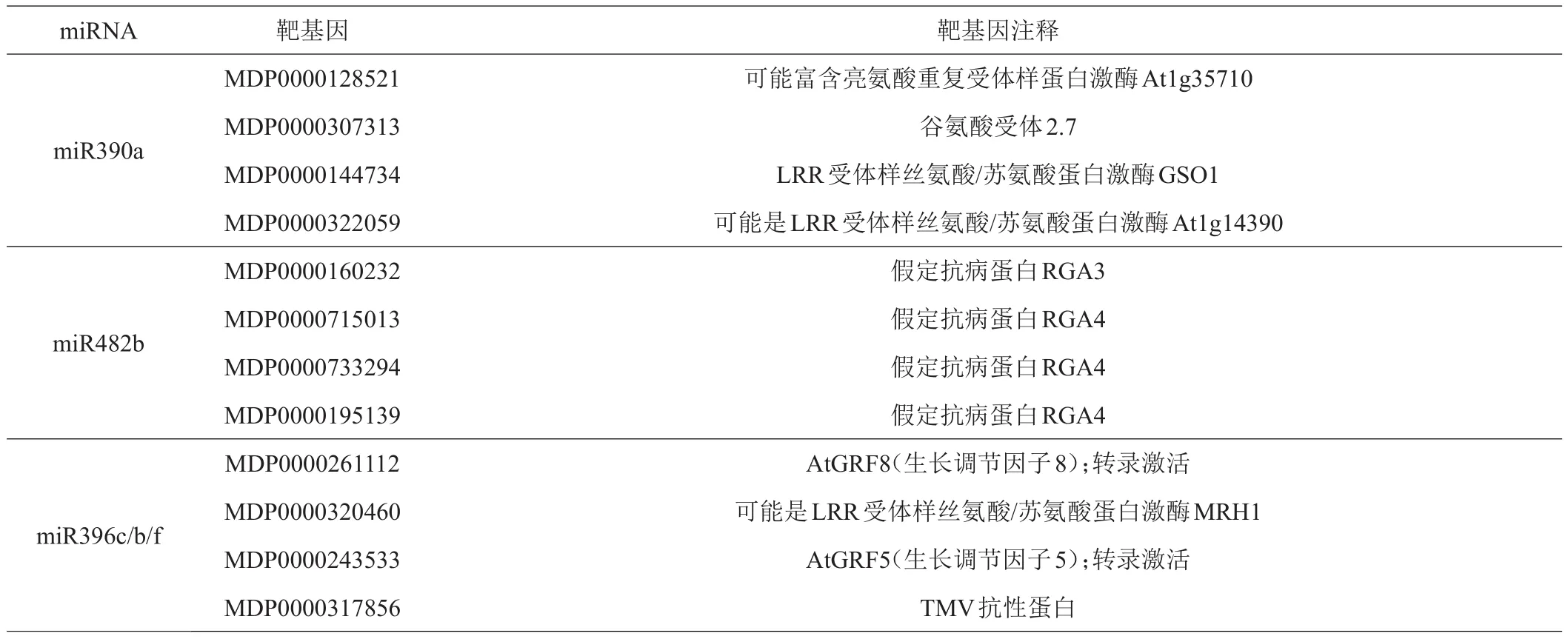
表2 相关靶基因预测
3 讨论
与动物相似,植物依靠细胞表面和细胞内的免疫受体来抵御病原体。植物先天免疫的第一道防线是通过细胞表面局部模式识别受体(PRRs)识别高度保守的微生物/病原体相关分子模式(MAMPs/PAMPs),如细菌鞭毛蛋白或真菌几丁质,从而引发PAMP触发的免疫(PTI),限制病原体的生长[15-16]。高度进化的病原体传递效应蛋白干扰PTI反应[17]。相应的,植物已经进化出细胞内核苷酸结合域富亮氨酸重复(NLR)类受体,识别病原体效应物并激活效应物触发免疫(ETI)[18-20]。NBS-LRR蛋白是一类己知的最大的R蛋白家族。近年来,关于miRNA响应非生物胁迫(如抗寒、干旱、重金属、抗盐等)的研究报道较多,而关于抵抗病原菌侵染及抗病相关研究相对较少。已有研究表明,miRNA参与了多种和抗病相关的基因表达,例如MdmiRLn11,尤其在病原菌感染条件下调控MdNBS基因的表达,并且MdmiRLn11靶向P-loop位点可能在木本植物中调控许多NBS LRR蛋白类基因[21]。同样在苹果中,MdmiRln20通过抑制MdTN1-GLS的表达负调控对GLS的抗性,并首次证明了miRNA在苹果对GLS的响应中发挥了关键作用[22]。杨树ptc-miR472a通过靶向NBS-LRR转录本,在植物对C.gloeosporioides和C.chrysosperma的免疫中发挥关键作用[23]。病毒诱导的miR390靶标基因沉默(VIGS)增加了棉花植物生长素和茉莉酸(JA)的积累,从而增加了对白粉虱侵袭的耐受性[24]。番茄中miR396通过抑制水杨酸和茉莉酸信号的防御相关基因,负调控番茄对晚疫病菌和灰霉菌侵染的耐受性[25]。试验初步鉴定了3个响应苹果属植物胶胞炭疽炭疽病菌侵染相关的miRNA,分别是miR390a、miR482b和miR396b/c/f,在抗性品种中miR390a和miR396b/c/f表达量下调,而在感病品种中表达量上调,且其靶基因均有LRR受体样丝氨酸/苏氨酸蛋白,miR482b靶基因为抗性蛋白,初步认定可能在苹果属植物炭疽病的侵染中起着重要作用,然而更加明确的分子作用机理还需要进一步研究验证。在研究植物miRNA在响应生物胁迫或非生物胁迫的功能过程中,靶基因是至关重要的。目前主要通过预测软件进行初步预测筛选,后续还应进行5’RACE实验,对切割位点进行确认,进而发现miRNA和其真实靶基因的网络调控关系,深入研究其作用机制。
猜你喜欢
今日农业(2022年4期)2022-06-01
当代水产(2022年1期)2022-04-26
小学生必读(中年级版)(2021年12期)2021-03-03
小学生必读(中年级版)(2021年11期)2021-02-22
宁夏医学杂志(2020年4期)2021-01-21
浙江农业学报(2017年1期)2017-05-17
新农业(2016年20期)2016-08-16
衡阳师范学院学报(2016年3期)2016-07-10
西南农业学报(2016年5期)2016-05-17
西南农业学报(2016年6期)2016-04-16
