
库姆塔格沙漠东南部胡杨液流变化及其对环境因子的响应
2021-03-26 06:53黄雅茹马迎宾李永华辛智鸣赵纳祺
草业科学 2021年2期
黄雅茹,马迎宾,李永华,辛智鸣,赵纳祺,杨 战,马 媛,吴 静,董 雪,李 帅
(1. 中国林业科学研究院沙漠林业实验中心 / 内蒙古磴口荒漠生态系统国家定位观测研究站,内蒙古 磴口 015200;2. 乌兰布和沙漠综合治理国家长期科研基地,内蒙古 磴口 015200;3. 中国林业科学研究院荒漠化研究所,北京 100091;4. 库姆塔格荒漠生态系统国家定位观测研究站,甘肃 敦煌 736200;5. 甘肃敦煌荒漠生态系统国家定位观测研究站,甘肃 敦煌 736200)
胡杨(Populus euphratica)是杨柳科乔木,作为极干旱区唯一的野生乔木种,其特点是耐干旱、耐寒、耐盐碱、喜光、抗风沙。胡杨是水土保持与防风固沙优良树种,对调节气候、稳定干旱区生态系统平衡、维持物种多样性发挥重要作用[1]。胡杨生态效益巨大,对水分需求量也较高[2]。干旱区水资源短缺,胡杨生长过程中应对干旱胁迫的水分利用策略及耗水特性是胡杨合理利用水资源的重要基础[3-4]。茎流计可以被用来监测树干液流,是研究树木耗水的主要方法之一,是衡量植物体内水分动态变化及其生理功能的关键指标,植物对水分的利用特征及其体内的水分传输状况通过树干液流速率来体现[5]。热扩散法操作容易,时间分辨率及准确度高,不影响树木的自然生长,能够实现长期的、连续性的自动监测[6-8]。
有关黑河、塔里木河胡杨树干液流的研究较多,研究发现胡杨液流速率日变化呈现明显的昼高夜低趋势[9-12],呈单峰型曲线[13],阴雨天,启动时间晚于晴天,停止时间早于晴天,峰值低于晴天[11],胡杨液流速率在夏季的启动时间、到达峰值的时间、液流停止时间均早于春季和秋季[11];整个生长季胡杨树干液流主要受光合有效辐射、土壤水分、风速、气温、相对湿度、水汽压亏缺的影响[10-11],液流速率与太阳辐射、气温、风速和土壤温度显著正相关,与空气湿度显著负相关[12-13]。胡杨夜间存在明显的液流[9-11],这是为了恢复植物体内的水分平衡,补充白天蒸腾消耗的水分[11]。白天,胡杨具有非常强烈的蒸腾作用,使得植物水分失衡,缺水严重,胡杨为了维持正常生长,必须补充足够的水分,因此在夜间胡杨根系在根压的作用下不断的吸水[9]。
前人对黑河、塔里木河的胡杨液流速率进行了较多研究,主要研究了液流速率的日变化、不同季节液流启动时间及影响因子[9-14],而对于不同季节液流速率对环境因子的非对称响应缺乏更为深入详细的分析。崔木土沟是甘肃敦煌西湖国家级自然保护区的主要保护区域之一,胡杨是库姆塔格沙漠东南部崔木土沟的主要建群种,构成了该区荒漠河岸生态系统的主体,对于稳定荒漠河流地带的生态平衡有重要作用,库姆塔格沙漠东南缘崔木土沟的胡杨沿沟道分布,主要依靠地下水生存,而且必须不断与地下水接触[15],但其水分传输规律、与环境因子的响应有待分析,精准测算单株植物的蒸腾耗水量前提条件就是摸清不同季节植物树干液流及其对环境因子的非对称响应规律,研究树木液流动态规律及其对环境因子的响应对河岸胡杨林的正常生长以及合理保护种群结构具有重要价值,对科学合理利用有限水资源具有重要意义。
本研究通过对库姆塔格沙漠东南缘季节性河流崔木土沟胡杨树干液流及太阳辐射、空气温度、空气相对湿度、风速的实时动态监测,揭示胡杨液流速率对环境因子的响应规律,旨在为干旱区荒漠河岸林胡杨林保护提供科学合理的依据,为当地合理利用与分配水资源提供技术支撑。
1 材料与方法
1.1 研究区概况
库姆塔格沙漠东南缘季节性河流崔木土沟全长约140 km,崔木土沟穿过崔木土山最终进入甘肃敦煌西湖国家级自然保护区。库姆塔格沙漠东南缘阿尔金山北麓是崔木土沟的发源地,行政区划属于甘肃省阿克塞哈萨克族自治县。研究区在没有发生大的降水时,阿尔金山冰雪融水是地表径流的主要来源,地表径流出山后,消散于山前洪积扇,随后,形成出露泉水(冲蚀沟内),流经15~20 km 后,出露的泉水很快消失,此时如果有较大的降水,便会形成地表径流出现在洪积扇内,最终在冲蚀沟内汇集[15]。降水、出露泉水、冰雪融水补给崔木土沟的径流。崔木土沟上游分布着4 条支流,分别是七里沟、大龙沟、红柳沟、柳城子沟[16]。棕漠土主要分布范围是山前洪积扇,同时也分布着盐土、灰棕漠土、风沙土,但棕漠土是主要的土壤类型[17]。研究区植物以胡杨、柽柳(Tamarix chinensis)、红砂(Reaumuria soongorica)为主,沿季节性河流崔木土沟沟道分布。
研究站点位于崔木土沟,沟道内有露泉水补给。胡杨样地位于沟道二级台地上,地下水埋深2~5 m。本研究的胡杨分布在崔木土沟上游,胡杨呈不连续片状分布,除胡杨外附近主要植物物种有柽柳和芦苇(Phragmites communis)。根据阿克塞县和敦煌市气象资料统计记录,崔木土沟上游年平均气温3.9 ℃,年平均降水量为 215.5 mm,蒸发量为2 500 mm;下游年平均气温9.4 ℃,年平均降水量为39.9 mm,蒸发量为2 486 mm[15]。本研究选取的胡杨林,样地面积为100 m × 100 m,林分密度为92株·hm−2,平均胸径为17.36 cm,平均树高为5.2 m,平均冠幅为2.10 m × 2.31 m。胸径分布频率如图1 所示,弧线是正态分布曲线,每个柱子代表胡杨胸径的一个径级。然后按照径阶分布测定8 棵胡杨标准木的胸径,分别为20.0、21.4、12.8、23.5、18.8、13.0、16.3、15.4 cm,树皮厚度在0.3~0.8 cm。胸径测定采用胸径尺,树皮厚度测定采用游标卡尺。
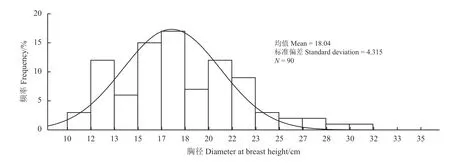
图1 胡杨胸径分布频率Figure 1 Frequency distribution of diameter at breast height for Populus euphratica
1.2 研究方法
胡杨液流速率测定采用Plant Sensors PS-TDP8,通过测定上、下探针之间的温差值,然后通过公式计算液流速率,上部的探针用一恒定电流加热,下部探针不加热。上、下温差值最大时,表示液流速率很小或等于零,上、下温差值最小时表示液流速率增大。PS-TDP8 树木茎流监测系统探针长度2 cm,探针安装方位是树干北侧,安装探针时距地面50 cm。探针的安装参照PS-TDP8 茎流系统说明书,探针安装完后先用泡沫软塑料包裹,再用锡纸包裹[18],每10 s 获取一次数据,每10 min 进行平均,并存储数据,数据采集仪为CR300S。气象因子(太阳总辐射TR、空气温度Ta、风速Ws、空气相对湿度RH)通过HOBO 小型自动气象站(美国生产)监测。液流监测时间为2018 年4 月1 日 − 10 月31 日。饱和水汽压差(VPD)公式如下:

式(1)中:Ta为空气温度(℃);RH 为空气相对湿度(%);a、b、c 为 常数,取 值 为0.611 kPa、17.502 kPa、237.3 ℃[19]。
1.3 液流速率计算

式(2)中: VS为树干边材液流速率(m·s−1),ΔT 为两探针间的瞬时温差值,ΔTm为24 h 内最大探针温差值。采用生长锥测定了90 株胡杨的边材厚度,根据边材厚度rs及胸径r 的实测值建立关系线性回归方程,R2= 0.887,得出k = 2.410 1、b = 1.262 8,边材面积As公式如下:

式(3)中:As为边材面积,r 是胸径,rb为树皮厚度,rs为边材厚度;式(4)为边材面积As与胸径r 的关系式。
1.4 数据处理
采用Excel 对胡杨液流速率与太阳总辐射、空气温度、风速、空气相对湿度进行数据处理及作图,采用SPSS 17.0 对胡杨液流速率与气象因子相关性进行分析,分析不同季节胡杨液流速率与太阳总辐射、空气温度、风速、空气相对湿度的回归模型。液流速率日均值为每日00: 00 − 24: 00 的平均值,液流速率连续日变化中春季为4 月16 日 − 20 日,夏季为7 月16 日 − 20 日,秋季为9 月16 日 − 20 日。液流速率与气象因子的相关系数采用每小时的液流速率数据。液流速率与气象因子的非对称响应采用日均值。
2 结果与分析
2.1 胡杨不同季节液流速率连续日变化
胡杨春季液流速率最小,秋季最大(图2)。春季波峰不明显,变化较平稳,秋季波峰明显,且液流启动后急剧上升到峰值,随后急剧下降,峰值持续时间短,夏季和春季液流启动后急剧上升到峰值,维持一段时间,峰值持续时间较长。夏季7 月份液流的启动时间最早,持续时间最长,液流启动时间为06: 00,08: 40 − 19: 30 出现波峰,最大液流速率在30.072~31.670 cm·h−1,夜间存在较高液流。春季液流启动时间为06: 10,09: 10 − 15: 30 之间达到最高值,最大液流速率为14.569 cm·h−1。秋季液流启动时间最晚,峰值最大,启动时间为06: 30,09: 50 −15: 10 之间出现峰值,最大液流速率为42.465 cm·h−1。不论春季、夏季和秋季,夜间有较高液流存在,春季夜间液流速率占全天的17.06%,夏季夜间液流速率占全天的18.72%,秋季夜间液流速率占全天的16.90%。
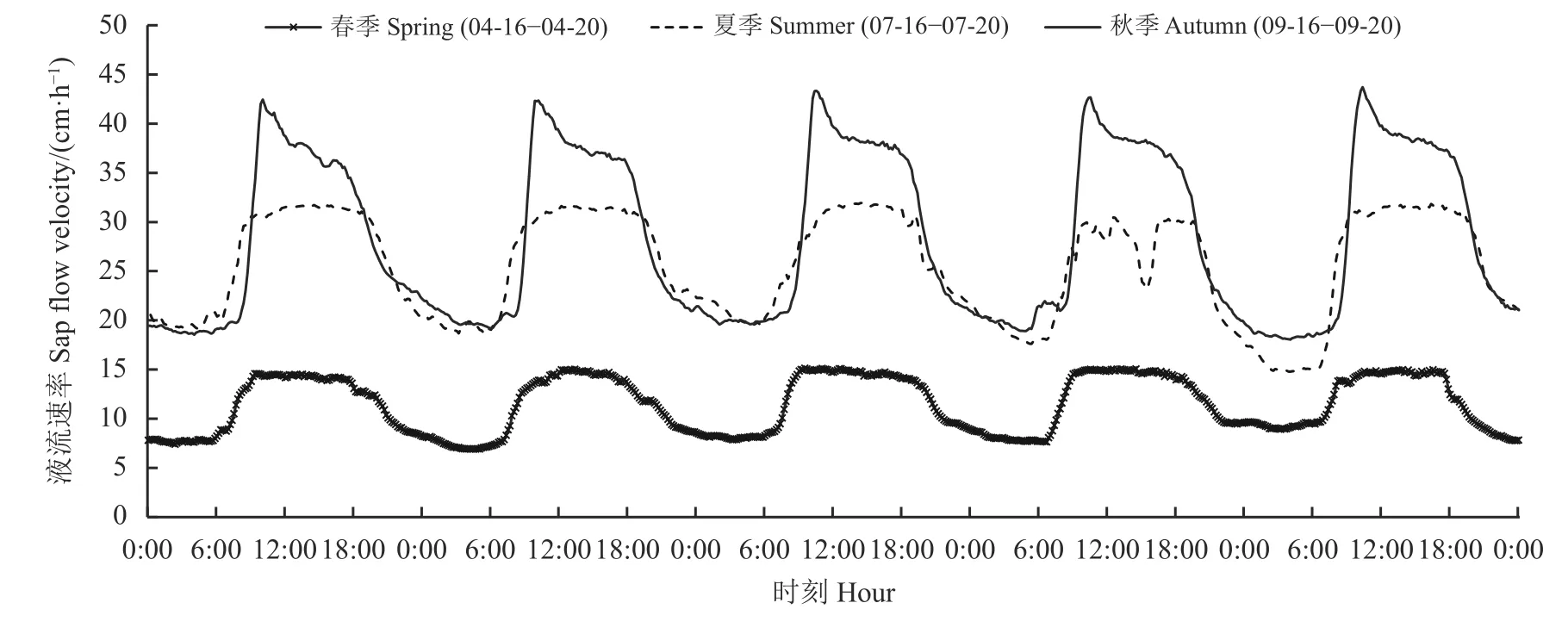
图2 胡杨不同季节液流速率连续日变化Figure 2 Continuous diurnal variation of sap flow velocity of Populus euphratica in different seasons
2.2 胡杨不同直径液流速率连日变化规律
不同直径胡杨液流速率具有相同的变化规律,呈几字“宽峰型”,没有明显的“午休”现象(图3)。液流速率白天高夜间低,夜间的变化幅度小。胡杨液流启动时间为05: 10 − 07: 10,峰值持续时间较长,第1 个峰值出现时间为11: 10 − 12: 10。在19: 00 − 20: 00液流急剧下降,21: 00 − 22: 00 下降至最小值,然后趋于稳定,一直维持稳定到第2 天液流启动时间。夜间也保持较高的液流速率(10.994~26.035 cm·h−1)。以8 月10 日为例,直径20.0、21.4、12.8、13.0、16.3、15.4 cm 胡杨茎干液流日平均值分别为30.894、35.682、25.103、30.051、29.336、32.001 cm·h−1。直 径21.4 cm 胡杨,液流启动时间为05: 10,第1 个峰值出现时间为07: 40,19: 00 以后缓慢下降,最低值出现在23: 20,此时,液流速率变化不大;23: 30 后,胡杨液流速率较为稳定,在夜间进行缓慢地输送,00: 00 − 06: 50 是 液 流的 最 小值,其 值 在21.972~27.513 cm·h−1。

图3 不同直径胡杨液流速率连日变化规律(2018 年8 月9 日 − 13 日)Figure 3 Diurnal fluctuation of sap flow velocity of Populus euphratica (Aug. 9th − 13th, 2018)
2.3 生长季胡杨液流日均值变化
胡杨液流速率日均值季节变化明显(图4 和图5)。胡杨液流速率8 月最大(8.180 cm·h−1),其次为9 月(7.680 cm·h−1),液流速率4 月最小(0.562 cm·h−1),7 月为6.646 cm·h−1,6 月为2.450 cm·h−1。日均液流速率最高值出现在8 月6 日,4 月18 日为最小日均液流速率,为0.189 cm·h−1,整个观测期间日均液流速率为4.428 cm·h−1。液流速率在降雨时减小,降雨后升高较为快速,8 月14 日至15 日有降雨,液流速率逐渐降低至6.122 cm·h−1,而在8 月17 日液流速率升高至8.546 cm·h−1,是降雨后最低液流速率的1.40 倍。
2.4 不同季节胡杨液流与气象因子的变化规律
2.4.1 不同季节胡杨液流与太阳辐射的变化规律
春季、夏季、秋季胡杨液流速率变化趋势与太阳辐射变化趋势一致(图6),胡杨液流速率随太阳辐射增加而增加。不同季节液流速率与太阳辐射存在时滞,液流速率到达峰值的时间比太阳辐射到达峰值的时间早。
2.4.2 胡杨不同季节液流速率与空气温度的变化规律

图4 2018 年生长季胡杨液流速率日均值变化Figure 4 Daily average change of sap flow velocity of Populus euphratica in the growing season of 2018
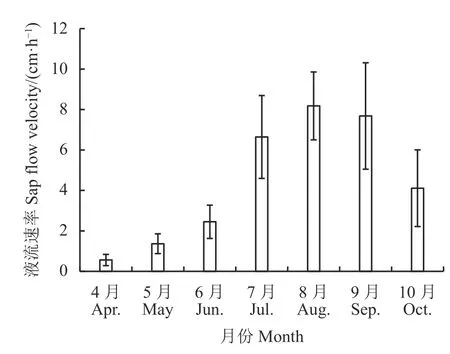
图5 2018 年生长季胡杨液流速率月均值Figure 5 Monthly average change of sap flow velocity of Populus euphratica in the growing season of 2018
胡杨不同季节液流速率与空气温度的变化规律基本相同(图6),空气温度增加,胡杨液流速率增加,空气温度降低,液流速率减小。春季、夏季、秋季胡杨液流速率峰值出现时间比空气温度峰值出现时间早,液流速率与空气温度存在时滞效应。产生时滞效应主要是因为,环境因子改变时液流速率的变化需经过叶片、枝条、树干的水容调节,而且植物液流速率会受水分或光胁迫,从而表现为液流密度相对于气象因子的滞后或提前。
2.4.3 胡杨不同季节液流速率与空气相对湿度的变化规律
液流速率变化趋势与空气相对湿度变化趋势相反(图6)。空气相对湿度增加,液流速率减小,空气相对湿度减小,液流速率增加。春季、夏季、秋季液流速率峰值出现时间早于空气相对湿度最低值出现时间,液流速率与空气相对湿度存在时滞效应。
2.4.4 不同季节胡杨液流与风速的变化规律
液流速率变化趋势与风速相同(图6)。白天液流速率随着风速的增大而增大,液流速率与风速呈正相关关系。
2.4.5 胡杨不同季节液流速率与饱和水气压差的变化规律
胡杨液流速率变化规律与饱和水气压差的变化规律大致相同(图6)。饱和水气压差增大,液流速率增加。不同季节液流速率与饱和水气压差存在时滞,液流速率峰值提前于饱和水气压差峰值的到来,这也是胡杨适应干旱环境的表现,胡杨液流传输过程在饱和水气压差到达峰值之前就达到了峰值,避开了峰值叠加效应。
2.5 胡杨液流速率与气象因子的非对称性响应
不同季节胡杨树干液流速率与太阳辐射、空气温度、空气相对湿度、风速、饱和水气压差日变化响应存在差异。春季、夏季、秋季,胡杨液流速率与空气温度、空气相对湿度、风速、饱和水气压差的日变化响应呈顺时针环状,用向上、向下的箭头表示(图7),在相同空气温度、空气相对湿度、风速、饱和水气压差条件下,上升阶段所对应的液流速率较大,春季、夏季、秋季,胡杨液流速率与太阳辐射响应的非对称性减弱,并且呈逆时针环状,主要是因为液流速率与太阳辐射的日变化更为一致。在同一太阳辐射值时,上升阶段所对应的液流速率较小。春季的环开度最小,秋季的环开度最大。

图6 树干液流速率与气象因子的变化Figure 6 Changes in stem sap flow velocity and meteorological factors
2.6 液流速率与气象因子的相关系数
春季、夏季、秋季胡杨液流速率均与太阳总辐射相关系数最高(表1),春季、夏季胡杨茎干液流与风速相关系数最小,秋季与空气相对湿度相关系数最小;液流速率与太阳辐射、空气温度、空气相对湿度、风速、饱和水气压差相关性均达到显著水平。采用SPSS 17.0 中的逐步回归分析,因变量设置为液流速率,自变量设置为气象因子,春季、夏季、冬季的回归模型如下:

图7 液流速率与气象因子的非对称性响应Figure 7 Hysteresis between sap flow velocity and meteorological factors
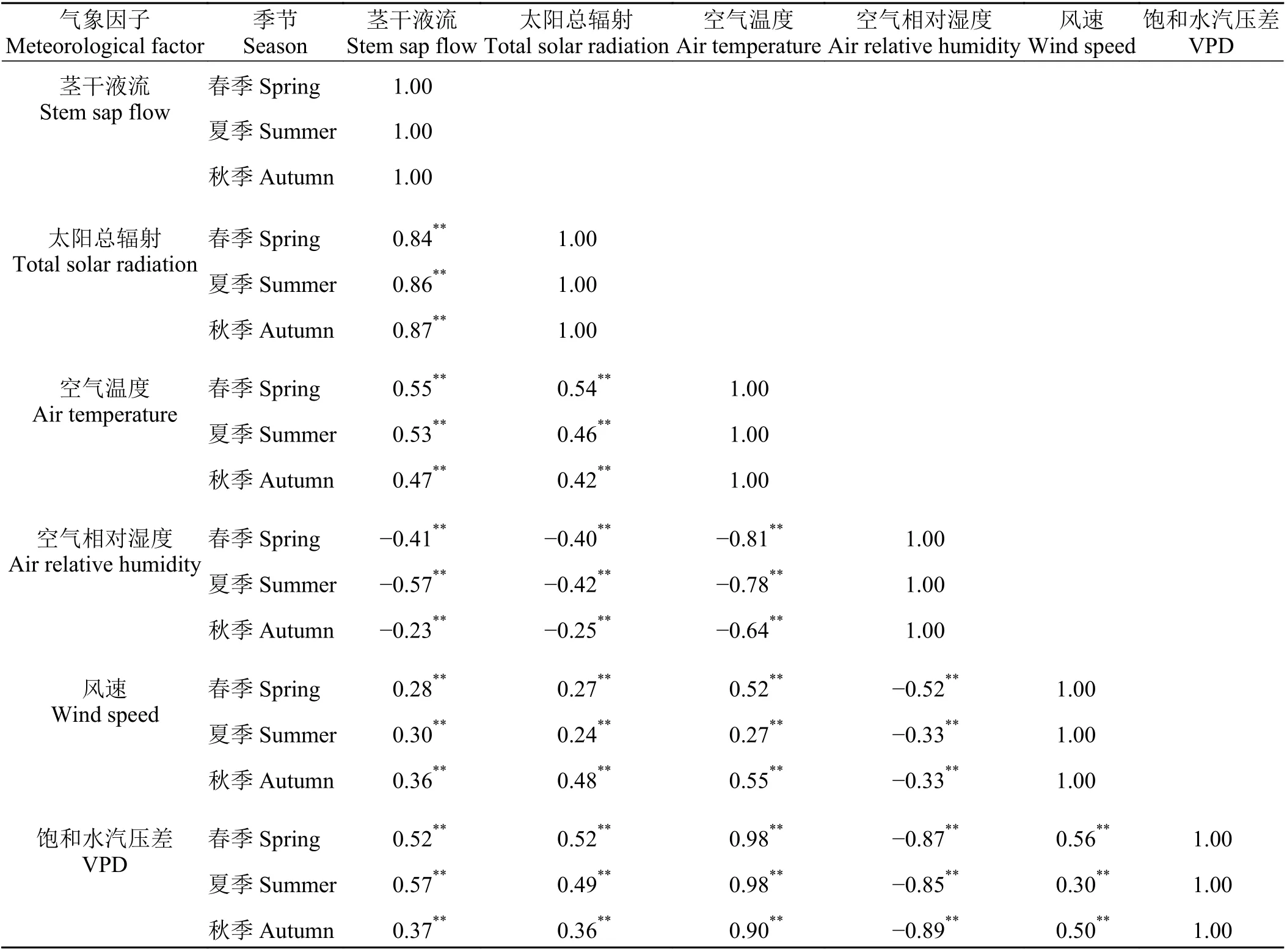
表1 液流速率与气象因子相关系数(n = 433)Table 1 Correlation coefficients between sap flow velocity and meteorological factors (n = 433)
春季:y = 7.839 + 0.007x1+ 0.090x2(R2= 0.719);
夏季:y = 24.606 + 0.012x1− 0.095x3(R2= 0.787);
秋季:y = 17.991 + 0.024x1+ 0.219x2(R2= 0.764)。
式 中:y 是 茎 干 液 流(cm·h−1);x1是 太 阳 总 辐 射(W·m−2);x2是空气温度(℃);x3是空气相对湿度(%)。
3 讨论与结论
3.1 讨论
赵春彦等[18]对胡杨树干液流的研究表明,胡杨树干液流速率的日变化呈几字“宽峰型”波动曲线,9 月份胡杨液流16: 00 左右到达峰值,峰值为38.096 cm·h−1。本研究结果表明,胡杨树干液流变化呈几字“宽峰型”,9 月秋季液流09: 50 − 15: 10 出现峰值,最大液流速率为42.465 cm·h−1。赵春彦等[11]研究发现,夏季胡杨树干液流的启动时间、到达峰值的时间均早于春季和秋季,这与本研究结果一致。
胡杨液流速率白天高夜间低,液流速率变化幅度白天大夜间小,这是由于气温白天高夜间低,植物蒸腾作用随温度的增加而增加,胡杨白天蒸腾作用强烈,使胡杨缺水严重,为了维持正常生长,夜间在根压的作用下不断的吸水来保持水分平衡[9]。李浩等[19]研究发现,夏季植物为了补充水分,夜间树干保持一定的液流来维持正常的生理活动。本研究中,夜间胡杨维持着一定的液流速率,这与司建华等[9]、赵春彦等[11]的研究结果一致。胡杨夜间存在较高的液流速率(10.994~26.035 cm·h−1),干旱区植物白天蒸腾作用强烈,失水严重,夜间通过根系吸水补充水分,因此夜间存在液流速率,胡杨耐旱性得到提高。
前人研究结果显示,土壤含水量、太阳总辐射、空气温湿度对植物液流速率产生主要影响,植物树干液流月变化主要受土壤水分影响,瞬时液流速率受气象因子影响[20-24]。胡杨液流速率与气象因子变化规律相同,说明气象因子对胡杨液流速率有明显的影响。太阳总辐射、空气温度与液流速率呈正相关关系,空气相对湿度与液流速率呈负相关关系。这与朱亚等[10]和买尔当·克依木等[12]研究结果一致。太阳辐射对植物光合、蒸腾作用产生直接影响,且呈正相关关系,太阳辐射增加,植物光合、蒸腾速率增加,液流速率也增加;空气温度主要对叶表温度产生作用,间接对蒸腾速率产生作用,植物为了降低高温的伤害,降低叶表温度,往往通过增加蒸腾速率来实现,叶片气孔导度增大,当空气温度达32 ℃时,植物叶片气孔阻力增大,而水势会下降,气孔导度减小,蒸腾作用减弱[25],因此茎干液流速率变小,本研究中,夏季午后空气温度在32.3~36.8 ℃,秋季最高气温29.47 ℃,夏季液流速率低于秋季液流速率;空气温度增加,空气相对湿度减小,植物蒸腾作用增加,液流速率增加,液流速率与空气相对湿度呈负相关关系,与空气水汽压差呈正相关关系[11]。风速对植物蒸腾作用也会产生影响,微风时,植物周围的相对湿度被吹走,蒸腾作用增加,风速较大时,植物叶片气孔关闭,植物蒸腾作用降低[26],因此,风速也会影响植物茎干液流的变化。
赵春彦等[11]研究表明,黑河下游胡杨春、夏、秋季液流速率的变化趋势一致,但夏季的变化幅度大于春季和秋季。买尔当·克依木等[12]研究表明,塔里木河夏季的液流速率最高,变化幅度高于春季、秋季。本研究结果显示,秋季的液流速率高于春季、夏季,秋季的变化幅度大于春季、夏季,与黑河、塔里木河的研究结果不同,这主要是因为胡杨生长的环境不同,黑河年平均降水量为37.4 mm,蒸发量为3 390 mm[11],塔里木河年平均降水量为41~76 mm,蒸发量为2 800 mm[12],敦煌崔木土沟上游山区年平均降水量为215.5 mm,蒸发量为2 500 mm,库姆塔格沙漠东南缘崔木土沟降水量高于黑河、塔里木河,蒸发量低于黑河、塔里木河,水分条件略优于黑河、塔里木河,就崔木土沟胡杨而言,可能是7 月到9 月雨季过后土壤水分得到较大补充,加之太阳辐射强度也较高,导致液流速率也较高,因此秋季液流速率高于夏季、春季。
本研究中,春季、夏季、秋季胡杨树干液流速率与太阳辐射、空气温度、空气相对湿度、风速的响应呈环状,是非对称性的。本研究区位于极干旱区的库姆塔格沙漠东南部,高温少雨,空气湿度较低,土壤水分条件也存在差异,再加上其他环境条件的限制,导致了胡杨液流速率对环境因子的响应规律存在差异。植物蒸腾速率受光照影响,光强增大,蒸腾速率增加,光强继续增加,光强增加到一定程度,也会抑制蒸腾速率(气孔导度受限制),夏季光照较强,部分气孔关闭,从而影响茎干液流变化[26]。胡杨液流速率随饱和水气压差、空气温湿度、太阳辐射的升高,大致呈直线趋势,这说明液流增加的主要驱动力是空气温度、太阳辐射、空气相对湿度,尚不足以诱导植物气孔调节行为。O’Brien 等[27]对热带雨林植物(10 种)树干液流变化研究发现非对称性变化最大的是液流与太阳辐射,这主要是由于热带雨林具有非常深的垂直结构,在冠层外和冠层内光照条件存在很大差别,而极干旱区荒漠地区的环境,空气温度及相对湿度条件与光照条件比对胡杨树干水分传输过程的影响更大。胡杨液流速率与气象因子的非对称性,春季的环开度最小,秋季的环开度最大,这主要是由于植物气孔对蒸腾调节的结果[28],非对称性的程度取决于太阳辐射和饱和水气压差的协同作用,与植物茎干的水分存在更加紧密地相关性[29-31]。不同季节胡杨对太阳辐射强度、空气温度、空气相对湿度、风速、饱和水气压差的响应规律存在差异,说明胡杨为了保持水分平衡应对水分亏缺的策略不同,在一定条件下(极端干旱)通过气孔调节降低水分损失,虽然样地处于极端干旱区,下层大气十分干燥,林冠层叶气界面水分散失很快,但崔木土沟供水良好,林冠层空气动力学条件相近,使得胡杨林蒸腾主要受叶面气孔控制[32]。
在自然环境中,如果水分供应充足,胡杨不会限制自身的蒸腾作用,相反会充分利用水分,当水分供应不足时,胡杨的蒸腾耗水也会相应减少,但不危及胡杨的存活,在这种状态下胡杨的生态功能也不会因此而降低,但是却减小了水分消耗,节约了水资源[[33-36]。这就会达到一个生态效应和水分利用的最优组合。从崔木土沟上游胡杨林目前自身的水分生理状态来说,以节约水资源为前提,以维持植物的基本存活为基础,降低胡杨耗水量,如果按照这样的水分管理模式,胡杨林还存在较大的节水空间。为了能够实现其生态效益和崔木土沟水资源的合理利用,应对胡杨采取适当的管护措施,为合理有效地利用水资源提供决策依据。因此,从崔木土沟水资源合理利用的角度出发,以最小的耗水量为前提,发挥最大的生态功能,下一步需要对胡杨最小耗水量问题进行深入研究。
3.2 结论
1) 胡杨液流速率变化规律呈几字“宽峰型”,没有明显的“午休”现象。胡杨白天液流速率较高,夜间液流速率较低。胡杨平均液流速率8 月最大,5 月最小。液流速率降雨时减小,降雨后增加。
2) 胡杨夏季液流的启动时间最早,持续时间最长,液流启动在早晨06:00,秋季液流启动时间最晚,峰值最大。春季、夏季、秋季夜间存在较高液流。
3) 胡杨液流速率变化趋势与周围气象因子变化趋势基本一致。胡杨树干液流速率和太阳辐射、风速、空气温度、饱和水气压差存在呈正相关关系,树干液流速率和空气湿度存在负相关关系。夏季太阳总辐射与空气相对湿度对胡杨液流速率产生关键影响,太阳总辐射和空气温度是影响春季、秋季胡杨液流速率的主要气象因子,均达到显著水平。
4) 春季、夏季、秋季,胡杨液流速率与空气温度、空气相对湿度、风速、饱和水气压差的日变化响应呈顺时针环状,与太阳辐射响应的非对称性减弱,并且呈逆时针环状。不同季节胡杨对太阳辐射强度、空气温度、空气相对湿度、风速、饱和水气压差的响应规律存在差异,表明胡杨为了保持水分平衡应对水分亏缺的策略不同,虽然样地处于极端干旱区,下层大气十分干燥,林冠层叶气界面水分散失很快,但崔木土沟供水良好,林冠层空气动力学条件相近,使得胡杨林蒸腾主要受叶面气孔控制,在一定条件下(极端干旱)通过气孔调节降低水分损失。
本研究结果对于明确干旱沙漠环境中胡杨水分传输对环境因子的响应规律、及更深入地认识胡杨适应干旱环境的水分利用策略具有重要意义。
猜你喜欢
金桥(2021年11期)2021-11-20
海洋通报(2020年5期)2021-01-14
北广人物(2020年47期)2020-12-09
创新作文(1-2年级)(2018年6期)2018-01-22
甘肃林业(2016年4期)2016-11-07
西南交通大学学报(2016年4期)2016-06-15
电源技术(2015年5期)2015-08-22
电源技术(2015年7期)2015-08-22
电测与仪表(2015年8期)2015-04-09
电网与清洁能源(2015年3期)2015-02-28
