
青桐幼苗对水分胁迫的生长及生理响应
2021-03-22 05:41朱孟一傅松玲
安徽农业大学学报 2021年6期
朱孟一,傅松玲*,孟 然,倪 正,张 荣,任 杰
青桐幼苗对水分胁迫的生长及生理响应
朱孟一1,傅松玲1*,孟 然1,倪 正1,张 荣2,任 杰3
(1. 安徽农业大学林学与园林学院,合肥 230036:2. 巢湖市林业技术推广中心,巢湖 238008;3. 安徽省农业科学院农业工程研究所,合肥 230031)
青桐观赏效果好,但对其受水分胁迫后的响应机理研究较少,制约了该物种下游产业的发展。为了探究青桐幼苗耐水程度及作用机理,对30%田间持水量(T1)、70%田间持水量(T2)、100%田间持水量(T3)和130%田间持水量(T4)水分胁迫下的青桐幼苗生长指标(株高、茎粗、叶面积)、生物量分配(叶生物量比、茎生物量比、根生物量比)与光合指标(净光合速率、蒸腾速率、胞间CO2浓度、气孔导度)进行分析。结果表明,随着水分胁迫程度的增加,幼苗的各指标表现为先上升后下降的趋势,当土壤达到T2条件时,幼苗各指标综合表现最佳;T3条件下青桐幼苗净光合速率最高,地上生物量分配较T2条件下高出4%;T2条件下,青桐幼苗叶面积最大,达到196.186 mm2,比叶面积最大,达到512.48,总生物量积累最多,达到17.14 g。当土壤水分达到T2条件时,最适合青桐幼苗生物量积累,可以为幼苗顺利过冬提供充足的储备碳;T4条件下胁迫时间达到10 d,幼苗生长情况出现拐点,胁迫前10 d内幼苗生长较快,但当水分持续性胁迫超过10 d后,幼苗生长趋势减弱,同时萌发出气生根用以响应淹水情况;T1条件下,对青桐幼苗影响极大,最不适合幼苗生长发育。研究结果为青桐幼苗受水分胁迫后的响应机制提供了理论基础。
青桐;水分胁迫;生长指标;生物量分配;光合指标
青桐(Linn. f.),别名中国梧桐,安徽省乡土树种。其树干端直,树皮绿色且平滑,叶大冠美。由于观赏效果好、管理成本低,天然分布于山区丘陵,现广泛用于城市行道树,巢湖市2017年将青桐定为市树。近年来巢湖夏季汛期水淹严重,甚至大部分生活区域被潮水淹没,人们生活严重受阻,城市绿化更是受到严重破坏。青桐作为巢湖市树又为肉质根系植物,水分胁迫对青桐生长及生理特性的影响相关研究还鲜见报道,因此对其耐水程度亟待研究。水分是影响植物生长发育的重要影响因子,科学有效地管理更加有利于减少干旱及涝渍的影响。目前,国内对青桐的研究更多在于对其育苗栽培技术方面的探索,对青桐幼苗在生长季初、旺盛期、末期受到自然性灾害的生理生化响应机制及应对措施鲜有研究。
水分胁迫的发生时间、持续时间、发生强度等均会对植物体的生长发育有较大的影响[1]。张婷等[1]研究指出,在生长季末期刺槐幼苗受到干旱胁迫后,NSC(非结构性碳水化合物)含量显著降低对刺槐幼苗休眠期代谢、应对其他环境胁迫以及春季叶片萌发产生严重的不良影响。Anderegg等[2]研究发现,生长季初期干旱胁迫会显著增加乔木NSC含量。Galvez等[3]发现,生长季旺盛时期干旱胁迫对乔木NSC含量没有显著影响。目前对植物体的生长季初期与生长季旺盛期进行的研究较多,植物体在此阶段下生长代谢旺盛,当其受到胁迫后表现也更为显著;相比之下,植物在生长季末期生长代谢变慢,胁迫后的表现也较不明显,但对植物体生长季末期受胁迫后的反应机制研究是必要的,有利于构建植物体生长季各阶段受胁迫后的响应机制完整蓝图。因此,本研究以一年生实生青桐苗为材料进行持续性水淹胁迫,设置4个水分梯度,模拟自然水分胁迫条件进行控水栽培,通过研究青桐幼苗对不同水分胁迫下的生长指标、生物量分配和光合指标等的响应,探寻青桐幼苗应对水分胁迫的生长及生理响应机制,为筛选抗涝抗旱的绿植提供参考依据[4]。
1 材料与方法
1.1 试验地概况及试验设计
试验样地位于安徽省合肥市安徽农业大学农萃园,31°48′ N,117°11′ E。北亚热带季风气候,年降水量950 mm,年平均气温为15.7~17.1 ℃,7月最高,为28.2~28.6 ℃,1月最低,为2.5~2.9 ℃。2020年6月初,将一年生种子繁殖的青桐幼苗移栽于Φ19 cm×H17 cm的育苗穴盆,每个盆钵土壤选择1:3比例的营养土与园区土壤混合物。试验前保证幼苗水分、光照等条件保持一致,并进行除虫除草等田间管理,3个月后选取生长良好、生长状况(株高、径粗和叶面积)相似的幼苗进行水胁迫试验。
2020年9月生长季末期开始进行水分胁迫试验。先从安徽农业大学农萃园试验样地取1 m以下的土壤,算出田间持水量为0.21 g·g-1,然后设置为4个试验水分梯度[5],分别是30%田间持水量(T1)、70%田间持水量(T2)、100%田间持水量(T3)和130%田间持水量(T4),每个处理5盆,重复3次。水分控制的方法采用EM50数据采集器[6]进行换算测定。从试验起(0 d)至试验结束(30 d),每日6:30和18:30用校准后的EM50对盆栽进行水分测量,保证土壤水分稳定;当盆内水分不足时,少量多次的加水至50 mL,等待2 min后再用校准后的EM50确认盆内土壤水分含量,直至达到设定的水分含量。
1.2 试验方法
1.2.1 生长指标测定 从试验初期(0 d)起至试验末期(30 d)止,每5 d于早上7:00—9:00之间测量并记录1次青桐幼苗各生长指标数据,每组处理重复3次,共测定12次。用直尺测量记录初始株高、试验后株高,用游标卡尺测量记录初始茎粗、试验后茎粗,计算出株高变化量、茎粗变化量;人工计数出测量的各株幼苗的叶数后,选取每株具有代表性的3片叶片,用YMJ-C型叶面积测定仪分别测量并记录叶面积、叶长、叶宽和叶长宽比,并计算出该株幼苗的总叶面积。
1.2.2 光合指标测定 在测量记录幼苗生长指标后,随即进行光合指标测定。用Li-COR6400便携式光合分析仪(Li-6400, Li-Cor, Lincoln, NE, USA)参考李旭[7]的研究,于9:00—11:00测净光合速率(n)、蒸腾速率(r)、胞间二氧化碳浓度(i)、气孔导度(s)、蒸汽压亏缺(VPD)参数,并计算出瞬时水分利用效率(WUE)=n/r。基于前期预试验结果,将测定时的光强参数设定为1 300 μmol·m-2·s-1,叶室温度设置为26 ℃,CO2浓度设定为500 mg·L-1。记录数据之前,将幼苗叶片在叶室内稳定5~10 min,使各个气体交换参数稳定[7]。
1.2.3 生物量分配测定 在测量记录幼苗生长指标及光合指标后,随即进行生物量指标测定。将12株幼苗整株挖出洗净,将每株幼苗的叶、茎、根3个部分分别装入牛皮纸袋内并对应标记,105 ℃杀青30 min后,在80 ℃下烘干至少72 h至恒重后称重记录[2],并计算叶生物量比(LBR,叶重/总生物量)、茎生物量比(SBR,茎重/总生物量)和根生物量比(RBR,根重/总生物量)。根据生长指标得出该株幼苗的叶片总面积,计算出比叶面积(SLA,总叶面积/叶重)。
1.2.4 数据统计分析 用Excel整理数据,用SPSS20分析,Origin2019进行绘图。
2 结果与分析
2.1 生长指标对水分胁迫的响应
不同水胁迫下青桐幼苗生长及其形态特征存在着明显差异(图1),随着胁迫时间不断增加,各生长指标的增幅逐渐减少。其中,在T2和T3条件下,青桐幼苗随胁迫时间增加,株高变化量、茎粗变化量、叶宽、叶长以及叶面积均呈现上升趋势;在T1条件下,青桐幼苗随着水分胁迫时间增加,其生长指标趋势均呈下降趋势,且受胁迫20 d的株高、茎粗指标显著低于(< 0.05)0 d;在T4条件下,青桐幼苗随着水分胁迫时间增加,其生长指标趋势均呈先上升后下降的趋势,且受胁迫20 d的叶面积指标显著低于(< 0.05)0 d。造成这种情况可能是,在T1条件下的土壤水分含量低极,不适宜青桐幼苗的生长发育;而T4条件下的土壤水分含量过高,青桐幼苗在受到极端水淹胁迫5 d内,因土壤水分充足,有利于促进幼苗生长发育,但随着胁迫时间超过5 d后,土壤透气性发生变化进而影响青桐幼苗根系呼吸,导致随着胁迫时间增加青桐幼苗生长受限。
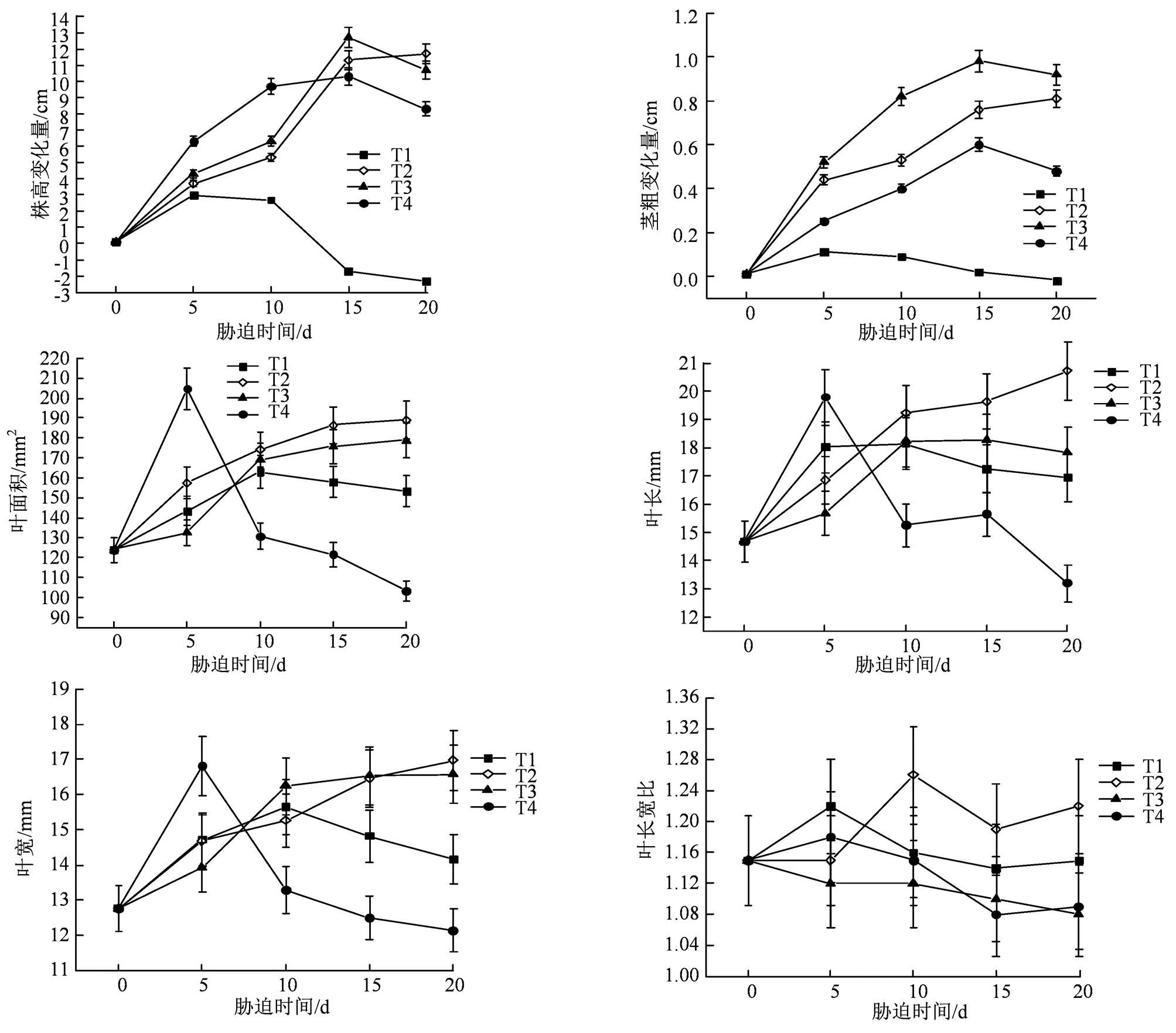
图中数据均为平均值±标准误, n=3。
Figure 1 The changes of the growth morphology ofseedlings during the whole experiment periods
单因素方差分析结果表明(表1),在4种水分胁迫条件下青桐幼苗生长情况出现较大差异。T1条件下,幼苗各生长指标受影响程度较大,株高变化量、茎粗变化量均显著低于(< 0.05)其他3种条件,分别为(–0.16±0.33)cm与(–0.13±0.01)cm,说明当幼苗长期处于T1条件下时,土壤水分含量不足,极不适宜青桐幼苗生长发育;T2、T3条件下生长指标综合表现较佳,其中株高变化量、叶长显著高于(< 0.05)其他两种条件,其中T2条件叶面积指标显著高于(< 0.05)其他3种条件,说明在T2、T3条件下时土壤水分含量较适宜幼苗生长发育;T4条件下,其茎粗变化量、叶长、叶宽、长宽比均显著低于(< 0.05)T2、T3条件,但显著高于(< 0.05)T1条件,而叶面积指标显著低于(< 0.05)其他3种条件,说明青桐幼苗长期处于T4条件下会影响其正常生长发育。从幼苗生长指标综合结果来看,青桐幼苗最适的条件从高到低为T2>T3>T4>T1。

表1 不同水分处理的青桐幼苗生长指标比较
注:表中数据为平均值±标准误,= 3,经过Duncan多重比较,不同字母表示差异显著(< 0.05)。下同。
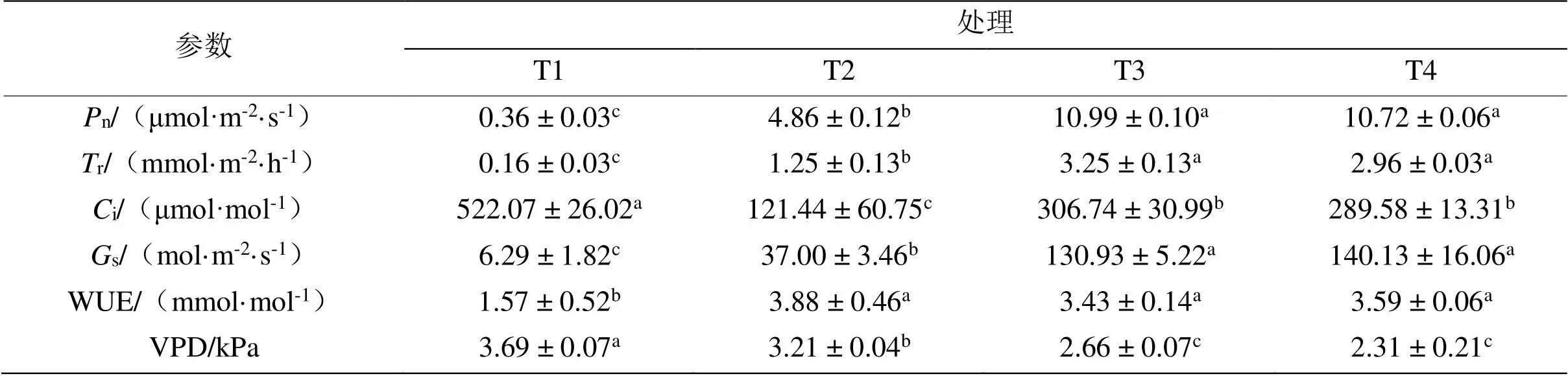
表2 不同水分处理的青桐幼苗光合指标比较
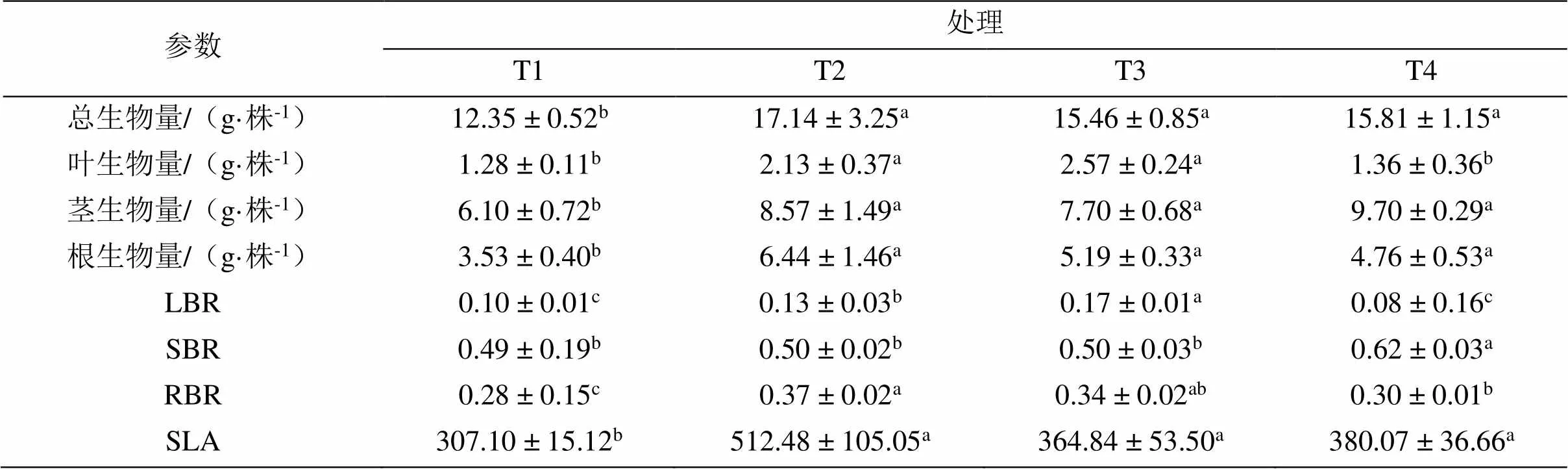
表3 不同水分处理生物量累计及其分配指标比较
2.2 光合指标对水分胁迫的响应
植物气孔导度在光合作用中起着重要的调节作用,它在控制水分损失和获得碳素即生物量产生之间的平衡中起着关键的作用[9]。影响植物气孔导度的因素有很多,其中土壤水分是重要影响因素之一[10],在植物受到轻度至中度的干旱胁迫的条件,气孔关闭是对干旱最早的响应[11]。由表2可知,青桐幼苗受胁迫后,其气孔导度从高到低一次表现为T4 > T3 > T2 > T1。试验末期,T1水分条件的n、r、s与WUE均显著低于(< 0.05)其他3种水分条件,s与VPD显著高于(< 0.05)其他3种水分条件;可以得出,在T1水分条件下,青桐幼苗处于缺水阶段不利于生长发育和生物量积累;T3与T4两种水分条件幼苗光合指标之间无明显差异(> 0.05),但对比总结发现,土壤水分含量较高的条件(T3与T4水分条件)幼苗的n、r、i、s与VPD含量均显著高于(< 0.05)土壤水分含量低的条件(T1与T2水分条件)。
从幼苗光合指标综合看出,青桐有较好的耐短期水涝的特性。在4种处理条件下,T1水分条件最不适合青桐幼苗生长,T2水分条件最适宜幼苗生长,T3水分条件稍次于T2水分条件,T4水分条件较一般。
2.3 生物量指标对水分胁迫的响应
不同的水分胁迫显著影响了青桐幼苗的生物量累积及其分配情况。由表3可知,4种处理下,总生物量在12.35 ~ 17.14 g之间,表现为T2 > T4 > T3 > T1,其中T1水分条件下幼苗总生物量与茎生物量显著低于(< 0.05)其他3种水分处理条件,分别为12.35 g与6.10 g;在青桐幼苗的叶生物量表现上,T2水分条件与T3水分条件显著高于(< 0.05)T1水分条件与T4水分条件;由于土壤过于干旱导致大量侧根失水死亡,使得T1水分条件根生物量与RBR均表现出显著低于(< 0.05)其他3种水分条件;随着水分胁迫程度的增加,LBR、RBR与SLA表现为先增加后降低的趋势,T2与T3水分条件下,LBR显著高于T1与T4水分条件;SBR表现为随着水分胁迫程度增加而增加,其中T4水分条件SBR表现最优,显著高于(< 0.05)其他3种水分条件,达到0.62。
3 讨论与结论
土壤水分条件直接影响了植物的生长情况,植物通过调节生长形态来适应土壤水分胁迫。T1(30%田间持水量)水分条件下,生长形态表现为须根发达的特点,但随着胁迫时间加强,侧根与须根多表现干枯死亡,幼苗受影响情况最为明显,生长状况不佳,最不适宜青桐幼苗生长发育;青桐幼苗在T2(70%田间持水量)水分条件下与T3(100%田间持水量)水分条件下的整体生长表现良好,其生长状态及生长指标随着胁迫时间增加均呈现上升趋势,土壤水分供应充足、叶子较多且叶面积大、光能利用效率较高,一定程度上维持了较高的碳获取能力;T4(120%田间持水量)水分条件下,幼苗生长状态发生变化,表现为部分叶片出现萎蔫且露出水面的部分萌生出部分气生根。结果为青桐幼苗在受到持续性水淹胁迫时的应对机制提供了新的思路。
水分条件显著影响青桐幼苗的气体交换和资源利用效率[8]。在T1(30%田间持水量)水分条件,为减少蒸腾作用叶片气孔关闭,净光合速率(n)显著降低,极大地影响青桐幼苗的总生物量积累;青桐幼苗受到极端水淹胁迫T4条件时,根的呼吸受到影响,幼苗通过改变其根部生长形态应对淹水胁迫,通过观察此阶段叶片的光合指标,净光合速率(n)、蒸腾速率(r)、胞间二氧化碳浓度(i)、气孔导度(s)以及蒸汽压亏缺(VPD)参数,与T2水分条件(70%田间持水量)有明显差异,但相互之间总生物量的积累无明显差异。可以推断出,青桐幼苗在受到持续淹水胁迫时,通过一定的响应措施寻求新的生存途径[12]。
土壤水分含量直接(光合原料减少)或间接(气孔关闭、酶失活等)影响了植物的光合作用,进而影响生物量的积累。在T1(30%田间持水量)水分条件下,土壤水分缺乏直接影响幼苗进行光合作用,导致幼苗总生物量累计较少;T2(70%田间持水量)水分条件下总生物量积累最多,株高与基径比例的降低有利于增加光能的利用效率[13],幼苗主要表现为对根生物量的积累;T3(100%田间持水量)水分充足条件下幼苗状态较好,生物量分配主要是对于叶生物量的累计;T4(120%田间持水量)水分条件过高的土壤湿度影响根系正常呼吸作用,进而影响幼苗正常的光合作用,使幼苗的总生物量积累受影响,此条件下幼苗生长状态表现为株高增长量受限,茎粗增长量影响不大,与Wang等[14]水分胁迫明显限制了幼苗的纵向生长、但对径向生长影响相对较小的结论相一致。因此,青桐对水分胁迫的强度和持续时间的有一定的响应规律,进而形成了不同的应对策略。
综上所述,幼苗受到极端干旱胁迫时,其生长指标及其生理指标均受显著影响。一方面幼苗通过改变自身形态来响应胁迫,说明青桐幼苗有适应干旱环境的潜力[15];另一方面,随着干旱胁迫时间的延长,幼苗根部、苗高、生物量分配以及光合指标等显著低于正常情况T2(70%田间持水量)水分条件,说明青桐幼苗抗旱性有限,需要在幼苗栽培过程中注意保水或补水。同理,青桐幼苗受也具有一定的抗涝能力,能够对水淹胁迫做出相应的适应性变化,可以在轻度淹水地区种植,但是长期受涝会对其根部产生影响,从而造成伤害[16]。可以通过加强良种选育与推广,选择耐涝品种;涝害发生后及时排水补救,恢复根系呼吸,同时补充肥料,加速代谢;推进青桐抗涝基因型鉴定与筛选的分子层面研究,有望培育出抗旱又抗涝的新品种[17]。
[1] 张婷, 曹扬, 陈云明, 等. 生长季末期干旱胁迫对刺槐幼苗非结构性碳水化合物的影响[J]. 水土保持学报, 2016, 30(5): 297-304.
[2] ANDEREGG W R, ANDEREGG L D. Hydraulic and carbohydrate changes in experimental drought-induced mortality of saplings in two conifer species[J]. Tree Physiol, 2013, 33(3): 252-260.
[3] GALVEZ D A, LANDHÄUSSER S M, TYREE M T. Root carbon reserve dynamics in aspen seedlings: does simulated drought induce reserve limitation?[J]. Tree Physiol, 2011, 31(3): 250-257.
[4] 杨曼, 张佑麟, 徐振东, 等. 水分胁迫对黑壳楠和香樟幼苗生理特性的影响[J]. 南方农业学报, 2015, 46(8): 1449-1454.
[5] 杨斌. 刺槐幼苗对水分胁迫的响应:基于生长、生理及非结构性碳的分配与动态[D]. 杨凌: 西北农林科技大学, 2019.
[6] 陈叶, 闫芳, 张彩丽, 等. 唐古特瑞香幼苗对干旱胁迫的生理响应[J]. 中药材, 2020, 43(6): 1312-1315.
[7] 李旭. 南亚热带季风常绿阔叶林优势树种叶片功能性状对增温的响应[D]. 合肥: 安徽农业大学, 2020.
[8] 王寒茹, 刘华, 李丕军, 等. 4个核桃类树种苗木应对持续干旱的生理响应[J]. 安徽农业大学学报, 2020, 47(5): 722-729.
[9] 徐飞, 郭卫华, 徐伟红, 等. 刺槐幼苗形态、生物量分配和光合特性对水分胁迫的响应[J]. 北京林业大学学报, 2010, 32(1): 24-30.
[10] ZHANG X L, ZANG R G, LI C Y. Population differences in physiological and morphological adaptations ofseedlings in response to progressive drought stress[J]. Plant Sci, 2004, 166(3): 791-797.
[11] 吴大千, 徐飞, 郭卫华, 等. 中国北方城市常见绿化植物夏季气孔导度影响因素及模型比较[J]. 生态学报, 2007, 27(10): 4141-4148.
[12] 朱政, 蒋家月, 江昌俊, 等. 低温胁迫对茶树叶片SOD、可溶性蛋白和可溶性糖含量的影响[J]. 安徽农业大学学报, 2011, 38(1): 24-26.
[13] FLEXAS J, MEDRANO H. Drought-inhibition of photosynthesis in C3 plants: stomatal and non-stomatal limitations revisited[J]. Ann Bot, 2002, 89(2): 183-189.
[14] WANG G G, BAUERLE W L, MUDDER B T. Effects of light acclimation on the photosynthesis, growth, and biomass allocation in American chestnut () seedlings[J]. For Ecol Manag, 2006, 226(1/2/3): 173-180.
[15] 曹林青. 油茶对干旱胁迫的生理生态响应[D]. 北京: 中国林业科学研究院, 2017.
[16] 杨斌. 刺槐幼苗对水分胁迫的响应:基于生长、生理及非结构性碳的分配与动态[D]. 杨凌: 西北农林科技大学, 2019.
[17] 张阳, 李瑞莲, 张德胜, 等. 涝渍对植物影响研究进展[J]. 作物研究, 2011, 25(4): 420-424.
Growth and photosynthetic characteristics of the seedlings ofunder water stress
ZHU Mengyi1, FU Songling1, MENG Ran1, NI Zheng1, ZHANG Rong2, REN Jie3
(1. School of Forestry and Landscape Architecture, Anhui Agricultural University, Hefei 230036; 2. Forestry Science and Technology Extension Center of Chaohu City, Chaohu 238008;3. Institute of Agricultural Engineering, Anhui Academy of Agricultural Sciences, Hefei 230031)
is an important tree species for afforestation, but the lack of research on its response mechanism after water stress limits the development of its downstream industry. In order to explore the water tolerance degree and mechanism of, we analyzed the growth indexes of seedlings (plant height, stem diameter and leaf area), biomass allocation (leaf biomass ratio, stem biomass ratio, root biomass ratio, root biomass ratio and root biomass ratio) and photosynthetic index (net photosynthetic rate, transpiration rate, intercellular CO2concentration and stomatal conductance) under 30% field capacity (T1), 70% field capacity (T2), 100% field capacity (T3) and 130% field capacity (T4). The results showed that with the increase of water stress degree, all indexes of the seedlings firstly increased and then decreased, and the seedlings had best performance under T2 condition; the net photosynthetic rate was the highest under T3 condition, and the above-ground biomass allocation was 4% higher than that under T2 condition; under T2 condition, the leaf area and specific leaf area of the seedlings reached the maximum of 196.186 mm2and 512.48, respectively, and the total biomass accumulation reached the maximum of 17.14 g. T2 condition was the most suitable for the biomass accumulation of the seedlings, which could provide sufficient carbon reserve for the seedlings to successfully survive the winter; under T4 condition, the seedlings grew rapidly within 10 days and the growth trend weakened after 10 days, meanwhile, the aerial root emerged in response to flooding; T1 was the most unsuitable condition for the development ofseedling. This study provides a theoretical basis for the responding mechanism of the seedlings under water stress.
; water stress; growth index; biomass allocation; photosynthetic indexes
S718.43;S792.37
A
1672-352X (2021)06-0883-06
10.13610/j.cnki.1672-352x.20220106.016
2022-1-6 20:43:34
[URL] https://kns.cnki.net/kcms/detail/34.1162.s.20220106.1254.032.html
2021-03-12
安徽省科技重大专项项目(18030701187)资助。
朱孟一,硕士研究生。E-mail:mengyizhu@ahau.edu.cn
通信作者:傅松玲, 教授,博士生导师。E-mail:fusongling@ahau.edu.cn
猜你喜欢
思维与智慧(2022年21期)2022-07-28
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
新疆农业科学(2020年1期)2020-02-14
高原山地气象研究(2016年2期)2016-11-10
初中生世界(2016年11期)2016-04-11
初中生世界·八年级(2016年3期)2016-03-23
中国麻业科学(2015年5期)2015-12-28
支点(2015年10期)2015-10-16
塔里木大学学报(2014年3期)2014-03-11
自然资源遥感(2014年2期)2014-02-27
