
两系亚种间杂交水稻组合的产量和品质性状杂种优势分析
2021-03-15 04:03邢运高王宝祥陈庭木杨波刘艳徐波孙志广迟铭李健李景芳方兆伟卢百关徐大勇
江苏农业科学 2021年1期
邢运高 王宝祥 陈庭木 杨波 刘艳 徐波 孙志广 迟铭 李健 李景芳 方兆伟 卢百关 徐大勇

摘要:为了明确两系亚种间杂交水稻组合主要农艺性状、产量性状以及品质性状的杂种优势,以广亲和两系不育系9311S的40个中熟中粳稻及由其配置的40个杂交F1为材料,分析播始天数、株高、穗长、单株有效穗数、每穗实粒数、每穗总粒数、结实率、千粒质量、一次枝梗数、二次枝梗数、单株产量、糙米率、长宽比、粒长、粒宽、透明度、垩白粒率、垩白度等18个性状的超优亲优势和竞争优势表现。结果表明,生育期存在正向超亲优势和竞争优势。株高、穗长、单株有效穗数、每穗实粒数、每穗总粒数、一次枝梗数、二次枝梗数、单株产量等8个性状存在正向超优亲优势和竞争优势,而结实率与千粒质量表现出负向超亲优势和竞争优势。品质性状中长宽比、粒长和垩白粒率存在正向超优亲优势,而糙米率、粒宽、透明度、垩白度表现出负向超优亲优势。长宽比、粒长存在正向竞争优势,糙米率、粒宽、透明度、垩白粒率、垩白度表现出负向竞争优势。与父本相比,两系亚种间杂交组合的一次枝梗数、二次枝梗数以及穗粒数明显增加,但是结实率和千粒质量明显降低,提高父本千粒质量是恢复系产量性状改良的重点。杂交组合品质优于父本是因为杂种的粒型变长,粒宽变窄,长宽比增加,有利于提升稻米的品质。
关键词:亚种间杂交稻;产量;品质;超优亲优势;竞争优势
中图分类号:S511.035.1文献标志码:A
文章编号:1002-1302(2021)01-0059-05
作者簡介:邢运高(1988—),男,安徽庐江人,硕士,助理研究员,研究方向为水稻遗传育种。E-mail:1009965234@qq.com。
通信作者:徐大勇,博士,研究员,主要从事水稻新品种培育研究与推广工作。E-mail:xudayong3030@sina.com
利用杂种优势是提高水稻产量的有效途径之一。由于籼粳之间差异显著,其亚种间杂种F1在营养器官和产量上表现出强的杂种优势[1]。近年来,已有不少籼粳间杂种利用成功的例子。浙江省科研单位利用粳型三系不育系与籼粳中间型恢复系配组,选育出甬优8号、春优84、嘉优中科1号等典型的籼粳杂交组合。利用强的籼粳杂种优势是两系法杂交水稻育种的战略重点,应用广亲和基因与两系光温敏不育系可以实现籼粳亚种间杂种优势的直接利用[2]。目前两系法利用籼粳亚种杂种优势主要是利用携带广亲和基因、具有粳型亲缘的籼型两系不育系与籼型或者偏籼型材料配组。如两优培九、Y两优1号、Y两优2号以及超优千号等[3-6]。浙江省嘉兴市农业科学院利用广亲和籼型两系不育系培矮64S与粳稻R2859组配,育成的培两优2859,其耐寒性明显好于籼稻,抗倒性好,不早衰,2006年通过浙江省审定[7-8]。当前两系籼粳亚种间杂交稻与三系法籼粳亚种间杂交稻相比,典型的籼粳亚种间杂交稻组合较少,还有较大的提升空间[8]。祁玉良等认为以早中熟品种为父本,能培育出强优势的亚种间杂交组合[9]。本试验以广亲和两系不育系9311S为母本,常规中熟中粳稻品种为父本,对两系亚种间杂交组合与其父本的产量与品质性状进行比较研究,并进行杂种优势分析,以期为两系亚种间杂交稻组合的选育提供理论依据。
1材料与方法
1.1供试材料
以两系不育系9311S为母本(9311S是由广亲和两系不育系培矮64S为不育基因供体,以9311矮秆突变体矮9311为轮回亲本,回交转育而成的温敏型广亲和籼型两系不育系。9311S属于偏籼类型,开花习性好,柱头外露率高,柱头较大,异交结实率高),以黄淮稻区40份中熟中粳稻为父本,2018年春天配制40份亚种间杂交种(F1)。竞争优势的对照材料为国家黄淮海粳稻区试对照品种徐稻3号。
1.2田间种植
2018年正季在连云港市农业科学院东辛基地种植40份F1以及父母本。F1与相应的父本成对相邻种植,5月10日播种,6月12日移栽,行株距为23cm×17cm,随机排列,单本移栽,种植6行,每行10株,重复3次。田间管理如一般大田管理。
1.3试验数据处理
水稻成熟后每个小区取中间10株考察株高、穗长、单株有效穗数、穗实粒数、穗总粒数、一次枝梗数、二次枝梗数、结实率、千粒质量、单株产量。外观品质利用万深SC-E米质判定仪测定。超亲优势及对照优势的计算公式如下:超优亲优势=(F1均值-优势亲本均值)/优势亲本均值×100%。一般而言,株高越高抗倒性越差,垩白粒率、垩白粒数值越小品质越优,因此株高、垩白粒率、垩白粒超优亲优势应以低亲为优[10];竞争优势=(F1均值-对照均值)/对照均值×100%。
2结果与分析
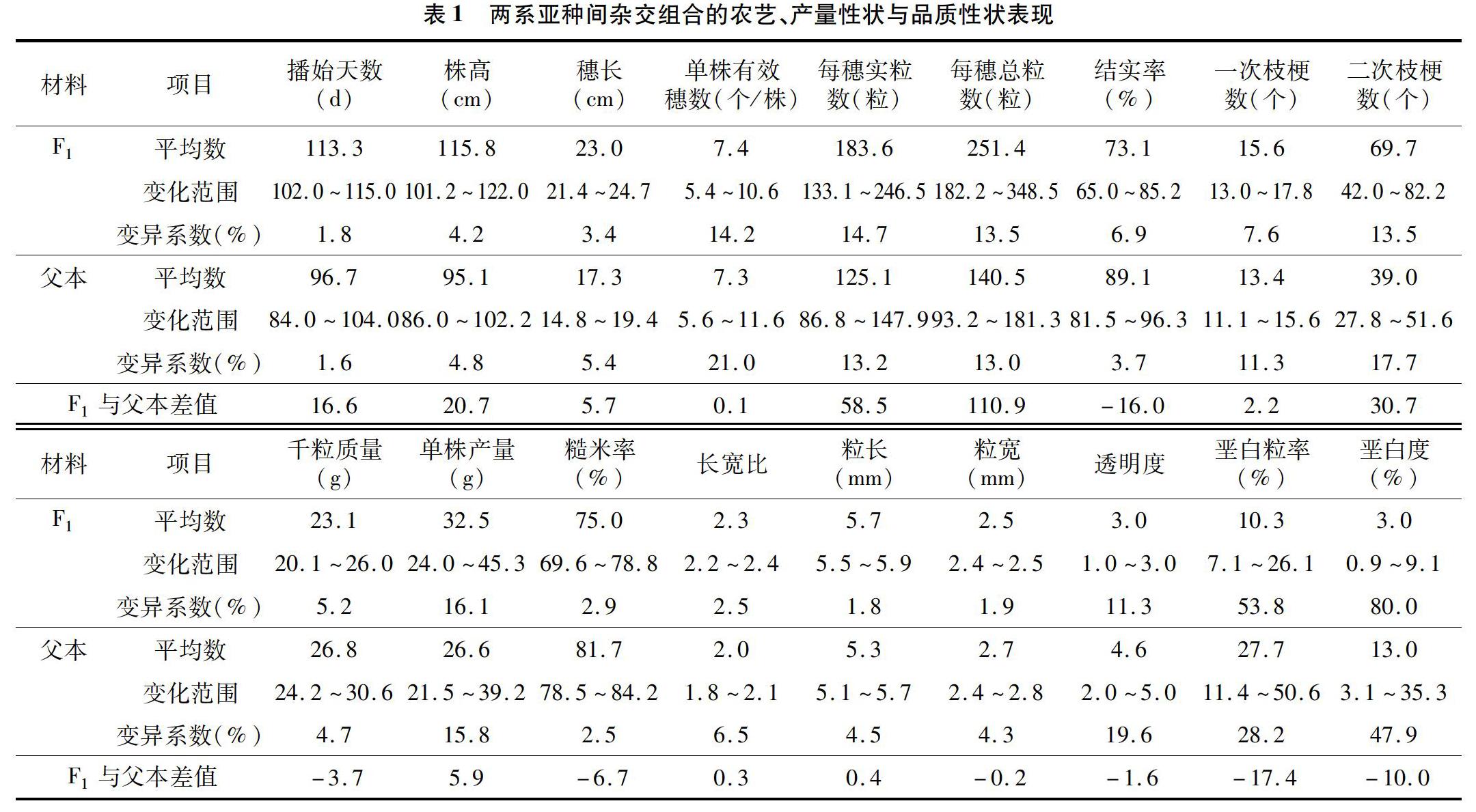
2.1两系亚种间杂交组合的农艺、产量性状与品质性状表现
两系亚种间杂交组合及其父本的农艺、产量和品质性状的表现见表1。父本18个性状变异系数变化范围是1.6%~47.9%,表明父本的遗传差异较大。其中农艺性状变异系数变化范围为1.6%~5.4%,产量性状变异系数变化范围为3.7%~17.7%,品质性状变异系数变化范围为2.5%~47.9%,因此,农艺性状间的遗传差异较小,而品质性状间的遗传差异较大。F1的18个性状变异系数变化范围是1.8%~80.0%,其中播始天数、粒长、粒宽、长宽比的变异系数较小,垩白粒率和垩白度的变异系数较大,说明播始天数、粒长、粒宽、长宽比的遗传变异较小,垩白粒率和垩白度的遗传变异较大。F1与父本平均数差值来看,农艺性状的平均值都超过了父本,其中株高平均值比父本增加20.7cm,穗长比父本增加5.7cm;8个产量性状中,结实率和千粒质量没有超过父本,结实率比父本减少16百分点,千粒质量比父本减少3.7g,其他6个性状的平均值都超过了父本。每穗实粒数、每穗总粒数、一次枝梗数、二次枝梗数分别比父本增加58.5粒、110.9粒、2.2个和30.7个,说明亚种间杂交种的穗型变长、变大,大穗优势明显。7个品质性状中仅粒长、长宽比2个性状超过了父本,粒长平均值比父本增加0.4mm,长宽比平均值比父本增加了0.3mm,而粒宽平均值比父本减少了0.2mm,说明杂种的粒型变长,粒宽变窄,长宽比增加,有利于提升稻米的外观品质。糙米率平均值比父本减少了6.7百分点,在选育亚种间杂交组合时,要注意对糙米率的筛选。垩白度和垩白粒率的平均值分别比父本减少了10.0和17.4百分点,说明两系亚种间杂交种的外观品质整体优于父本。
2.2两系亚种间杂交组合农艺性状与产量性状的超优亲优势与竞争优势
由表2可以看出:(1)11个性状超优亲优势变异系数变化范围为10.6%~486.0%,竞争优势变异系数变化范围为10.1%~114.2%,表明11个性状的超优亲优势和竞争优势在组合间表现差异较大,差异程度因性状不同而异。超优亲优势变异系数中单株有效穗数最大,而株高最小。竞争优势变异系数中单株有效穗数最大,穗长最小。(2)生育期和农艺性状的正向超优亲频率和超标出现频率为100%,表现出明显的杂种优势。株高的超低亲优势明显,达到了65%,表现出籼粳交后代株高超亲的一般规律。每穗实粒数、每穗总粒数和二次枝梗数等3个产量性状的正向超优亲频率和超标出现频率皆为100%。二次枝梗数超亲优势最大,其超亲优势达82.4%;其次是每穗实粒数,其超高亲优势达80.8%。40个组合的每穗实粒数和二次枝梗数的优势均高于徐稻3号,竞争优势平均值分别为94.1%、145.3%,说明亚种间杂交种穗大粒多,库容大的特征。(3)结实率和千粒质量的超亲优势和竞争优势为负值,超优亲频率和超标频率为0%,表现出明显的负向优势。总的来说,生育期存在正向超亲优势和竞争优势;农艺和产量性状中株高、穗长、单株有效穗数、每穗实粒数、每穗总粒数、一次枝梗数、二次枝梗数、单株产量等8个性状存在正向超优亲优势和竞争优势,而结实率与千粒质量表现出负向超亲优势和竞争优势。
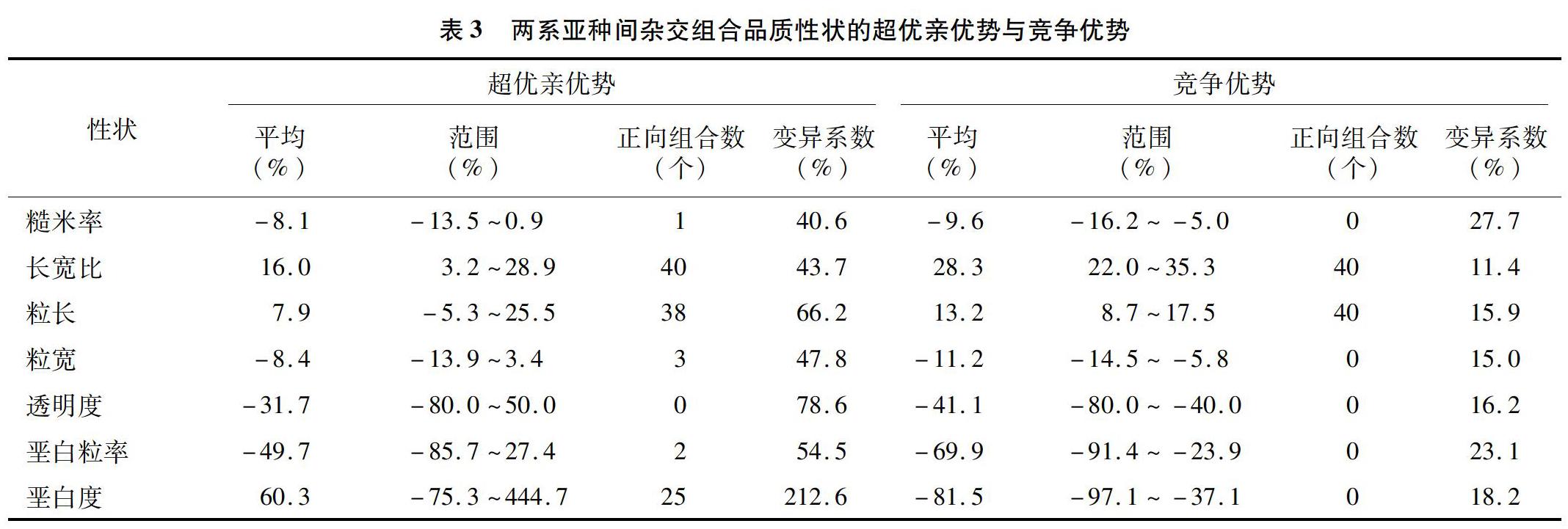
2.3兩系亚种间杂交组合品质性状的超优亲优势与竞争优势
由表3可知,7个品质性状超优亲变异系数变化范围为40.6%~212.6%,竞争优势变异系数变化范围为11.4%~27.7%,表明7个品质性状的超优亲优势不同性状间差异较大,而竞争优势在不同组合间差异较小。垩白度的超优亲优势变异系数最大,为212.6%,糙米率的变异系数最小,为40.6%;糙米率的竞争优势变异系数最大,为27.7%,而长宽比的竞争优势变异系数最小,为11.4%;长宽比、粒长和垩白度的正向超优亲优势出现频率分别为100%、95%和62.5%,其中垩白度的优势最强,超过低值亲本60.3%,这是一个不利的超亲优势表现;糙米率、粒宽、透明度、垩白粒率表现出明显的负向超优亲优势;品质性状的竞争优势与超优亲优势不同,只有长宽比和粒长2个性状的表现出正向的杂种优势,且优势出现频率均为100%,垩白粒率和垩白度负向竞争优势,这是2个有利的竞争优势表现。在籼粳间杂交稻选育时要考虑垩白度超亲的问题。总的来说,品质性状中长宽比、粒长和垩白粒率存在正向超优亲优势,而糙米率、粒宽、透明度、垩白度等4个性状表现出负向的超优亲优势。长宽比、粒长存在正向竞争优势,糙米率、粒宽、透明度、垩白粒率、垩白度表现出负向竞争优势。
2.4F1与父本相应性状的相关与回归
考察播始天数、株高、穗长、单株有效穗、每穗实粒数、每穗总粒数、结实率、一次枝梗数、二次枝梗数、千粒质量、单株产量、糙米率、长宽比、粒长、粒宽、透明度、垩白粒率、垩白度等18个性状与父本对应性状进行相关和回归分析,结果(表4)表明,18个性状都呈正相关,其中千粒质量、穗长、垩白度、一次枝梗数、二次枝梗数、长宽比、单株产量、每穗实粒数、每穗总粒数、株高等10个性状显著或极显著相关。从相关系数的大小看,F1代与父本性状的相关程度为千粒质量>穗长>垩白度>一次枝梗数>二次枝梗数>长宽比>单株产量>每穗实粒数>每穗总粒数>株高。对与父本相关达显著的性状做进一步分析,千粒质量的回归系数为0.569,即父本总粒数每提高1百分点,F1就相应提高0.569百分点,因此选择千粒质量大的品种作父本,可以增加F1的千粒质量,进而提升杂种产量。
3讨论与结论
籼粳亚种间具有强大的杂种优势,利用亚种间杂种优势是提高产量潜力的主要途径,两系法有利于亚种间杂种优势利用[11]。本试验选用的广亲和两系不育系9311S的广亲和基因源自培矮64S。由于广亲和基因的作用,9311S与典型粳稻品种配组表现出良好的亲和性。研究两系不育系与粳稻父本配制杂交种的优势表现,对于利用亚种间杂种优势具有重要的意义。
籼粳杂交稻的特点是产量优势显著、高秆大穗、有效穗中等或较少,生育期较长,并且主要表现为灌浆期的延长[8]。本研究中农艺性状和产量性状大多数性状存在正向超亲或竞争优势。本研究中F1比父本(常规中熟中粳稻)播始天数迟16.6d,超亲优势和竞争优势的平均值分别为17.1%和15.5%。由于F1超亲晚熟,导致与本研究中组配的杂交组合不适宜在淮北地区种植。这与王才林等研究典型杂交粳稻抽穗期多表现为负向优势,有利于选育早熟组合研究结果[12]不同。生物产量是水稻高产的基础,高产品种需要合适的株高,但是株高超过一定限度就与抗倒伏能力存在负相关关系。吕川根等提出长江中下游杂交稻理想株型的株高为110~120cm[13]。本研究F1株高范围是103.8~122cm,符合生产上的需求。40个组合株高全部超过父本和徐稻3号,表现出籼粳亚种间杂交稻株高超亲的一般规律。F1与父本株高的相关系数为0.319,达到了显著相关水平,因此选择株高合适的恢复系能选育理想的杂交种。张宏根等认为结实率低是制约杂交粳稻产量优势发挥的主要限制因素,提高结实率和千粒质量是恢复系产量性状改良的重点[14]。虽然9311S含有广亲和基因,但是本研究表明中结实率比父本低16%,千粒质量比父本低3.7g,40个组合的结实率和千粒质量超亲优势和竞争优势值均分负值,表现出明显的负向超亲优势和竞争优势。结实率和千粒质量偏低是制约两系亚种间杂交稻利用的关键因素。F1与父本千粒质量的相关系数为0.604,达极显著相关,因此提升恢复系的千粒质量,可以提升杂种F1的产量。
亚种间杂交种的穗形大、着粒密、灌浆慢,导致品质下降,是限制两系杂交稻利用的另一因素。吕建群等认为大部分品质主要受基因加性效应影响,糙米率受恢复系的影响较大[15]。F1的糙米率与父本的相关系数为0.173,不显著相关,这与吕建群等的研究结果不同。戴正元等研究认为垩白粒率和垩白度主要受恢复系影响[16]。本研究结果表明两系亚种间杂交种的品质多介于双亲之间,优于父本和对照品种(徐稻3号)。F1的垩白度与父本的相关系数为0.389,达显著相关,改良恢复系的垩白度可以改善亚种间杂交种的品质。
参考文献:
[1]万建民.水稻籼粳交杂种优势利用研究[J].杂交水稻,2010,25(增刊1):3-6.
[2]黄群策.中国两系法籼粳杂交水稻育种的研究进展[J].科技导报,1995,13(11):46-48.
[3]邹江石,吕川根,王才林,等.两系杂交稻“两优培九”的选育及其栽培特性[J].中国农业科学,2003,36(8):869-872.
[4]杨建君,刘志坚,谭鹏飞.杂交水稻‘Y两优1号特征特性及高产栽培技术[J].南方农业,2013,7(2):81-82.
[5]李建武,邓启云,吴俊,等.超级杂交稻新组合Y两优2号特征特性及高产栽培技术[J].杂交水稻,2013,28(1):49-51.
[6]魏中伟,马国辉.超高产杂交水稻超优1000的生物学特性及抗倒性研究[J].杂交水稻,2015,30(1):58-63.
[7]蔡文平,郑智明,狄小江,等.两系籼粳杂交稻培两优2859特征特性及栽培技术[J].现代农业科技,2008,(16):206.
[8]宋昕蔚,林建荣,吴明国.水稻籼粳亚种间杂种优势利用研究进展与展望[J].科学通报,2016,61(35):3778-3786.
[9]祁玉良,鲁伟林,石守设,等.水稻两系亚种间杂种优势分析及其亲本选配的研究[J].河南农业科学,2005(10):35-38.
[10]张新忠,吕亮杰,吕超,等.大麦农艺性状与产量性状的杂种优势分析[J].麥类作物学报,2016,33(1):39-43.
[11]吕川根,姚克敏,李霞,等.两系法亚种间杂交稻育种的若干思考[J].中国农业科技导报,2007,9(2):38-43.
[12]王才林,汤玉庚.杂交粳稻三系育种的基础研究Ⅰ.优势表现及其与亲本的关系[J].杂交水稻,1987(1):16-21.
[13]吕川根,邹江石.两个超级杂交稻与汕优63光合株型的比较分析[J].中国农业科学,2003,36(6):633-639.
[14]张宏根,孔宪旺,朱正斌,等.粳稻三系亲本的性状特征与杂种优势分析[J].作物学报,2010,36(5):801-809.
[15]吕建群,刘光春,任鄄胜,等.5个籼型不育系和5个新恢复系的品质性状配合力及遗传力分析[J].西南农业学报,2014,27(1):12-17.
[16]戴正元,李爱宏,刘广青,等.几个优良籼稻亲本品质性状的配合力和杂种优势分析[J].植物遗传资源学报,2006,7(2):234-238.
猜你喜欢
矿山安全信息(2022年40期)2022-04-07
当代水产(2021年10期)2021-12-05
今日农业(2020年20期)2020-11-26
聚氯乙烯(2018年9期)2018-02-18
现代园艺(2016年17期)2016-10-17
现代园艺(2016年17期)2016-10-17
今传媒(2016年9期)2016-10-15
投资者报(2016年33期)2016-09-30
