
白藜芦醇的生理功能及其在猪生产中的应用研究进展
2021-03-15 05:40:02李岩利胡睿智谭继君伍小松
中国畜牧杂志 2021年3期
李岩利,胡睿智,谭继君,伍小松,2*
(1.湖南农业大学动物科学技术学院,湖南长沙 410128;2.湖南农业大学动物医学院,湖南长沙 410128)
白藜芦醇(RES)的化学名称为反式-3,4´,5-三羟基二苯乙烯,由2 个芳香环组成且通过亚甲基桥连接。RES 最早于1939 年被 Takaoka 从白叶藜芦的根部分离出而被发现[1],是一种天然的植物多酚,在松树、花生和大豆等植物中广泛存在[2]。目前研究已证实RES 具有抗氧化[3]、抗炎[4]、抑菌[5]、抗病毒[6]、改善脂肪沉积[7]、能量和脂质代谢调节[8]的作用,RES 与氧化损伤和心血管疾病的发病机制密切相关[9-10]。本文就RES的生理功能及其潜在作用机理进行总结,并结合其在猪生产方面的研究报道,探讨其作为功能性饲料添加剂改善猪生产性能的应用前景。
1 RES 的来源、合成和提取工艺
RES 有顺式和反式2 种同分异构体,顺式RES 在自然界中不稳定,极易转化为其反式异构体,因此,通常所说的RES 往往是其反式结构。RES 的分子式为C14H12O3,相对分子质量为228,为无色针状晶体,不溶于水,易溶于甲醇、乙醚、氯仿等有机溶剂,熔点为256~257℃[11]。RES 的2 种异构体如图1 所示。

图1 RES 的结构式
当植物应对外界伤害或真菌攻击时,会产生RES来抵抗外界伤害带来的影响[12]。RES 在植物体内的合成通过苯丙烷途径完成[13]。如图2 所示,以苯丙氨酸作为合成RES 的起始底物,苯丙氨酸解氨酶(PAL)为催化剂,经过非氧化脱氨转化为反式肉桂酸,反式肉桂酸又经过肉桂酸-4-羟化酶(C4H)的催化,生成对香豆酸(p-CA),再经4-香豆酰辅酶A 连接酶(4CL)的催化作用,将p-CA 与辅酶A(CoA)的泛酸基团连接,生成4-香豆酰CoA,最后在RES 合酶(STS)的催化下,4-香豆酰CoA 和丙二酰CoA 结合生成RES。对于某些植物含酪氨酸裂解酶(TAL),可直接催化酪氨酸(Tyr)而产生p-CA,再经4CL、STS 催化过程最终生成RES[14]。其中,STS 是整个RES 合成途径的关键酶,底物丙二酰CoA 是其限制性因素[15]。当植物处于逆境威胁的情况下,才会激活STS 催化的这一合成途径[16]。

图2 白藜芦醇的生物合成途径[13-14]
提取RES 时因溶剂不同而得到的提取率不同。单辉君等[17]研究表明,不同溶剂提取RES 的效率为甲醇>95%无水乙醇>乙酸乙酯>水。提取RES 的方法包括有机溶剂提取、碱提取、酶解提取、超声提取、微波提取、超临界CO2提取等。姚庆收等[18]用有机溶剂提取法、碱提取法和超声提取法从花生根部及其愈伤组织中提取RES,其中超声提取法的提取率最高,且操作简单、用时短。张青松[19]用有机溶剂提取、酶解、超声提取、超临界CO2提取RES,其中超临界CO2提取的纯度最高,但提取率最低。瞿卫林等[20]用酶辅助提取RES,提取率较有机溶剂提取、碱提取和超声提取的常规提取方法提高3~5 倍。因此,生产中可选择使用超临界CO2提取,加酶辅助提取的方法获得的RES 纯度和提取率较高。
微生物合成和化学合成RES 也是得到RES 较常用的方法。微生物合成RES 主要是通过将4CL和STS在微生物中表达,把香豆酸添加到培养基中作为前体物质,来实现RES 的微生物合成[21],目前所利用的微生物主要是大肠杆菌。董良媛[22]通过用大肠杆菌BL21 将葡萄RS基因构建pET22b-RS 表达载体,来转化大肠杆菌BL21(E.coliBL21,DE3),再通过添加p-CA 和丙二酰CoA 后获得产量为25.2 mg/L 的RES。Watts 等[23]利用转化有阿拉伯拟南芥的4CL基因和落花生的STS基因的大肠杆菌JM109 菌株,以p-CA 为底物,得到了100 mg/L 的RES。RES 的人工合成方法也是合成RES的可行办法且早已受到各国学者的关注。目前化学合成RES 的主要方法:①Witting 及Witting-Homor 路线合成,此方法所需条件温和,产物易分离,研究人员多利用此种方法来合成二苯乙烯类化合物,已被广泛应用[24]。②Perkin 反应,此方法可得到单一的白藜芦醇产物,很好地保持其立体构型[25]。③Herk 反应,此方法的关键步骤是通过Herk 反应在2 个苯环之间形成碳碳双键,产物为单一的RES[26]。
2 RES 的生理功能
2.1 抗氧化和清除自由基作用 多酚物质中酚羟基的数目与其抗氧化能力有关[27],RES 分子中的3 个酚羟基均能清除自由基。活性氧(ROS)含有未成对的电子或基团,RES 的酚羟基可失去一个氢质子,将其转移给ROS,ROS 接受质子后失去了氧化性质,同样失去质子的2 个白藜芦醇分子又可以互相共用电子对,发生结构重排,从而起到清除自由基的作用。RES 分子中的4'-羟基基团与苯环形成共振效应连接对二苯乙烯骨架,使得4'-羟基基团有很强的反应活性,是RES 发挥抗氧化作用的重要基团[28]。RES 能够清除氧自由基和羟自由基,且有体外试验表明其清除过氧化亚硝酸离子(ONOO-)的能力强于维生素C[29]。体外试验中,通过将官能团逐次引入苯环,发现RES 的3 个酚羟基均具有抗氧化作用,且酚羟基数量减少会降低其抗氧化活性[30]。潘润天[31]通过对比RES、2,6-二叔丁基-4-甲基苯酚(BHT)和维生素C 在体外清除羟基自由基、ABTS 自由基、DPPH 自由基和抑制亚油酸过氧化的能力,结果发现RES 清除羟基自由基和抑制亚油酸过氧化的能力最强,且显著高于BHT 和维生素C。
当生物机体受到有害刺激时,会产生过量ROS,使原本存在的氧自由基动态平衡被打破,导致机体组织损伤[32]。机体内存在2 种抗氧化系统:①非酶参与的抗氧化系统,通过维生素C、维生素E、谷胱甘肽(GSH)等直接作用清除自由基。②需酶参与的抗氧化系统,通过激活Ⅱ相解毒酶等清除自由基。通过雄性小鼠和缺血再灌注损伤大鼠的试验表明,RES 能上调抗氧化酶的活性,如过氧化氢酶(CAT)、超氧化物歧化酶(SOD)、谷胱甘肽过氧化物酶(GSH-Px),同样可使机体丙二醛(MDA)含量有所降低[33-34]。
RES 主要通过上调核转录因子E2 相关因子2/ 抗氧化反应元件(Nrf2-ARE)信号通路来实现其在体内的抗氧化作用。Nrf2 是细胞抵御氧化应激时发挥调节作用的一个重要因子[35]。在正常机体状态下,Nrf2 被胞浆蛋白伴侣分子(Keap1)抑制[36]。当细胞处于氧化应激状态时,Keap1 的抑制作用解除,Nrf2 进入细胞核,结合小分子蛋白Maf 后,再结合抗氧化反应元件(ARE),进而启动Nrf2 下游抗氧化蛋白的转录,如醌氧化还原酶1(NQO1)、血红素加氧酶1(HO-1)、SOD、GSH-Px 等,发挥抗氧化作用[36]。RES 可通过催化还原型烟酰胺腺嘌呤二核苷酸氧化酶(NADPH oxidase,Nox)亚基蛋白水平的下调抑制其亚基Rac1膜转位和p47phox 磷酸化从而抑制NADPH 氧化酶介导的ROS 生成,对于某些特定底物,RES 可以直接激活沉默信息转录调节因子1(SIRT1),也可以通过磷酸二酯酶(PDE)抑制的通路间接激活SIRT1,SIRT1 有助于使叉头转录因子(FOXO)上调SOD2、CAT 和内皮型一氧化氮合酶(eNOS)的水平,从而发挥抗氧化作用[30,37]。RES 还可通过激活过氧化物酶活化受体Y辅助活化因子1α(PGC-1α),上调线粒体抗氧化酶表达,从而降低ROS 生成[38]。此外,通过大鼠心肌细胞和小鼠膝关节软骨细胞模型试验表明,RES 可通过上调磷脂酰肌醇-3 激酶/ 蛋白激酶(PI3K-Akt)信号通路激活Nrf2,发挥抗氧化作用[39-40]。但同时也有研究表明,RES 在人U251 胶质瘤细胞模型中可抑制PI3KAkt 信号通路使细胞免受应激损伤[41],所以对于不同的细胞类型和刺激因素,RES 对PI3K-Akt 信号通路的调控存在偏差。RES 还可通过激活丝裂原活化蛋白激酶(MAPK)通路中的胞外信号调控激酶(ERK1/2)、c-Jun氨基末端激酶(JNK)、p38 来影响细胞的抗氧化反应[42]。
2.2 抗炎作用 当机体受到炎症细胞因子和促炎细胞因子等有害物质的刺激时,机体内一系列的防御性反应会被引发,从而引起炎症的发生。机体内适度的炎症可以清除外源或自身变异的抗原,修复机体损伤[43],但炎症因子的过度激活会导致机体组织器官的损伤[44]。有研究表明,肝细胞内脂肪沉积产生的大量ROS,会导致溶酶体破裂,引起溶酶体酶的释放,进而激活核转录因子-κB(NF-κB)炎症通路[45]。NF-κB 是炎症反应中重要的转录调节因子,正常情况下受抑制性亚单位IκB 的抑制作用[46]。当外界刺激时,会激活NF-κB 信号通路,NF-κB 进入细胞核,促进促炎细胞因子的表达,如肿瘤坏死因子α(TNF-α)、白介素1β(IL-1β)、白介素6(IL-6)等。同时,TNF-α和IL-1β可反过来激活NF-κB 通路[47]。此外,脂多糖、病毒、紫外线和ROS 均能激活NF-κB 通路[48]。RES 可抑制IκBα激酶的降解,阻止NF-κB 进入细胞核来发挥其抗炎作用[49]。Manna 等[50]研究表明,RES 能抑制不同种类细胞中NF-κB 的活化,包括U-937、HeLa 和H4,且RES 可以剂量和时间依赖性下调因TNF-α诱导的NF-κB 信号通路。Estrov 等[51]研究表明RES 能阻断IL-1β诱导的NF-κB 的活化。Adhami 等[52]研究表明,RES 可阻断IκB 的活化、磷酸化和降解,进而抑制NF-κB 通路。RES 还能通过激活AMPK 信号通路和SIRT1 使NF-κB 去乙酰化,减少其转录[53],在敲除SIRT 表达的角质细胞中,RES 可逆转SIRT1 的表达从而抑制紫外线诱导的JNK 活化,缓解细胞炎症损伤[54]。在以白癜风白斑病灶周围皮肤的角质形成细胞模型中的研究发现,RES 可通过抑制JNK 和p38的磷酸化来缓解炎症[55]。此外,RES 还可通过抑制环氧合酶(COX)和过氧化物酶体增殖物激活受体(PPAR)等炎性介质的表达缓解炎症[53]。RES 抗氧化和抗炎的潜在作用机理如图3 所示。
2.3 抑菌作用 RES 作为一种天然的植物抗毒素,对很多细菌和真菌都具有抑菌作用。李巍伟[56]研究发现,经RES 体外作用后的葡萄球菌细胞壁层次不清,细胞器结构模糊、细胞质内含物消失、出现空泡的现象,表明RES 可破坏葡萄球菌,同样RES 作用于皮肤藓菌后也出现核膜断裂、线粒体内有空泡、线粒体呈致密化和髓样化的特征,表明RES 对皮肤藓菌有抑菌作用,但对革兰阴性的大肠杆菌无明显的抑制作用。RES 可增强抗生素的抗菌活性,如加强氟喹诺酮类抗生素对金黄色葡萄球菌和大肠杆菌的活性[57]。彭如洁[58]通过RES对幽门螺杆菌(Hp)的体外抗菌试验发现,31.25 mg/L RES 的单独抑菌率与8 mg/L 甲硝锉的抑菌率无显著差异,RES 与甲硝锉的联合抑菌率显著高于甲硝锉的单独抑菌率,且与甲锉联合作用时提高了耐药Hp 对抗生素的敏感性。RES 对一些菌的抑菌效果见表1。但目前来说,RES 抗菌方面的试验研究还有限,其具体机制尚不清楚。根据已有的报道得知其可能的抑菌机制:①RES与细菌ATP 酶可逆结合,进而抑制细菌ATP 水解和合成,从而影响细菌在体内代谢,RES 还可抑制细菌对糖的代谢而导致细菌生长抑制[59-60]。②RES 可诱导细菌DNA 片段化和SOS 应激反应[61]。③RES 可抑制细菌生物膜稳定性和细菌鞭毛的合成,降低细菌的集群效应和运动特性[62]。
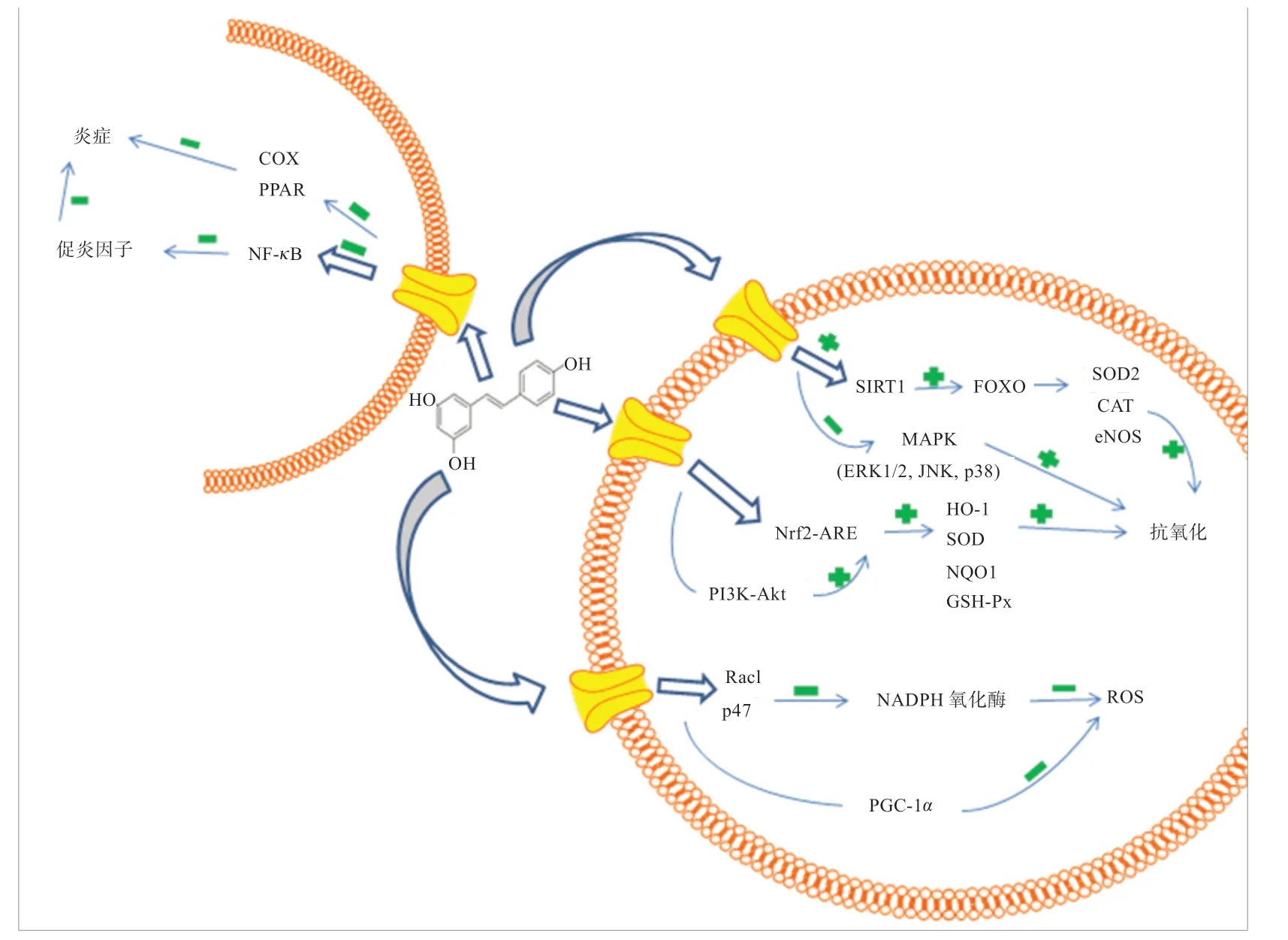
图3 RES 抗氧化和抗炎的潜在作用机理[36-42,45-47]

表1 RES 对不同细菌的最低抑菌浓度和抑菌效果
3 RES 在猪生产上的应用
RES 有助于改善动物体内的脂质过氧化表达,增强机体内的抗氧化系统。猪精子细胞膜含有多不饱和脂肪酸,在保存过程中会受ROS 影响,使细胞膜脂质过氧化,从而引起精子质膜和顶体的破坏,降低精子的受精能力[65]。范晓腾[66]将RES 应用于猪精液的常温保存中,发现第5 天的精液能达到与鲜精ROS 水平一致的效果,而正常保存精液的精子ROS 水平与鲜精相比却显著降低。在仔猪上,Zhang 等[67]研究发现,与未做处理的IUGI 仔猪相比,在患有宫内生长迟缓(IUGI)哺乳仔猪的饲粮中添加RES 提高了IUGI 仔猪的饲料利用效率,有效减轻了IUGI 引起的线粒体肿胀和超氧自由基的生成,有效提高了肝脏线粒体DNA 含量、ATP生成量。Ahmed 等[68]研究发现日粮中添加RES 显著提高了灌服大肠杆菌和沙门氏菌的仔猪干物质消化率和血清免疫球蛋白G(IgG)水平,对饲料转化率有改善作用,降低了TNF 水平,增强了仔猪对沙门氏菌和大肠杆菌的防护作用。在育肥猪上,Zhang 等[69]通过在日粮中添加300 mg/kg 或600 mg/kg RES 显著提高了肌肉总抗氧化能力(T-AOC)和GSH-Px 的mRNA 水平,降低了MDA 含量。Zhao 等[7]研究发现,RES 能有效抑制猪伪狂犬病毒引起的NF-κB 通路的活化。庞卫军等[70]研究发现RES 可显著抑制猪前提脂肪细胞增殖和分化,减少细胞内脂肪的合成。
综上所述,RES 在养猪生产中具有以下潜在应用价值:①可作为猪精液常温保存中较好的抗氧化剂和抗凋亡剂,作为猪精液稀释液中的营养物质添加使用,以增加精子的保存时间和保存效果,提高母猪受精率和仔猪整体质量。②作为辅助剂同抗生素共同使用来部分替代抗生素,增强抗生素效果,从而达到减抗的目的。③商品猪饲粮中可添加适宜比例的RES 来提高其饲料转化率和生长性能等指标,增强商品猪的抗病性和肉品质,提高经济效益。
4 小结与展望
饲用抗生素滥用不仅造成环境污染,严重威胁食品安全与人类健康,从某种意义上来说也增加了养殖的附加成本。无抗养殖是未来畜牧业的重要趋势。RES 具有抗氧化、抗炎、抑菌等多种生物学功能,可有效改善猪的免疫性能,提高猪的繁殖和生长性能,预防疾病,改善猪肉品质,因此可作为饲用抗生素的潜在替代品应用于养猪生产中。但RES 本体在体内的生物利用度相对较低,在人类和不同动物模型中均可被迅速吸收、代谢和排泄。因此,利用最新的代谢组学技术探究RES 在动物体内的代谢,并阐明相关代谢产物的生理功能对合理利用RES 至关重要。在当前非洲猪瘟背景下,基于RES 及其代谢产物的生理功能及作用机理将其合理应用于提高猪的免疫和抗病力,为养猪生产提供新的选择与思路。
猜你喜欢
当代水产(2021年6期)2021-08-13 08:17:10
当代水产(2020年4期)2020-06-16 03:23:22
当代水产(2020年3期)2020-06-15 12:02:52
科学中国人(2018年8期)2018-07-23 02:26:46
中成药(2018年2期)2018-05-09 07:20:05
新乡学院学报(2016年6期)2016-12-01 05:21:38
当代化工研究(2016年9期)2016-03-20 16:22:11
特产研究(2015年1期)2015-04-12 06:36:09
食品工业科技(2014年15期)2014-03-11 18:17:20
食品工业科技(2014年15期)2014-03-11 18:17:15
