
X射线光谱在生命起源和全球气候变化等若干重大科学问题中的研究进展
2021-03-09 10:36罗立强沈亚婷
光谱学与光谱分析 2021年3期
罗立强,沈亚婷
国家地质实验测试中心,北京 100037
引 言
生命从哪里来,到哪里去? 是自然科学探索的永恒主题。寻找生命起源,研究生命进化,分析影响人类命运的重大事件,是国际科学研究的前沿领域。
生命的诞生与进化,并非一帆风顺。一些重大事件的发生必然会产生显著性影响。发生在66百万年前的小行星撞击事件,直接导致了地球上76%的物种消失[1]; 全球气候变化,也正加剧自然生态恶化,增加人类患病风险,严重威胁着人类生存环境。控制和降低大气二氧化碳(CO2)浓度、减缓全球气候变暖趋势,是我们共同面对和关注的重大科学问题。
本文简要概述了国际上在生命起源、地球早期生命寻迹、全球气候变化等重大科学问题中的主要研究进展。本文并不对这些领域作全面评述,而主要关注以X射线荧光(XRF)和X射线吸收(XAS)为代表的X射线光谱分析技术在相关领域的研究与应用,以揭示其重要作用和潜在价值。将关键科学问题聚焦于: (1)生命起源和进化研究中的物质组成和结构形态; (2)地球早期生命碳成因研究中的元素、物相和生物功能团分析; (3)海洋碳循环及全球气候变化中的铁形态与科学意义。
1 生命起源与X射线光谱分析技术
生命起源有多种学说,如源于陨石或彗星等[2]。化学观点认为生命始于有机物合成[3],主要包括RNA有机合成[2]及深海热液新陈代谢[4]两种学说。这两种理论假说相互间既有差异也协同互补[5]。
1.1 RNA有机合成理论与XRS
RNA有机合成理论(有机汤理论)认为,地球初始大气组分富含CH4,NH3和H2,在紫外线辐射下产生氰化氢和乙醛等有机分子,经加氢反应后生成氨基酸,氨基酸的加氢聚合反应生成含核酸基团的腺嘌呤等物质[2,6]。再经进一步浓缩和聚合产生缩氨酸及蛋白质,产生具有自催化和自复制功能的分子[2-3,5],由此开始RNA、DNA和蛋白质等的复制、信息编码、序列转录、转移和突变等生命历程[6]。
目前,基于氰氨、羟乙醛、磷酸盐等的初始化学成分合成实验已取得显著性进展[3]、在太阳系外星系云盘中也发现了CH3CN和HCN等多样性氰化物[7]证据,支持并解释了RNA有机合成理论的合理性。但另一方面,RNA有机合成理论还有一些问题待解,例如关于基因编码的产生机制[6]、RNA合成反应效率及专属性影响因素等[2]。
随着生命诞生,地球大气从富CO2向富氧环境转变。在此过程中,气相组成与性质对RNA的生物分子结构也产生了重要影响。最近有研究运用X射线吸收近边结构(XANES)谱分析C形态[8],结合模拟温压热降解实验,发现在粘土矿Mg蒙脱石和富N2/O2条件下,有机质由富含N的芳香族杂环化合物组成; 而在富CO2条件下,则主要为脂肪族和氨基化合物,揭示冥古宙和太古宙时期的大气组成与性质是决定RNA结构的关键性因素。
水和核酸等有机聚合物是生命诞生的两个基本条件。作为核酸基本组成的嘌呤类化合物是RNA形成中的重要产物。但目前关于腺嘌呤、鸟嘌呤等的形成机制知之甚少[2]。最近有研究采用XANES技术测定水溶液条件下的氮、氧K边吸收谱[9],获得了RNA基本核苷酸单元——单磷酸鸟嘌呤核苷在溶液状态下的分子结构,揭示了形成机制[9-10]; 此外,还可利用XANES研究RNA大分子折叠机制及化学突变作用机理,探索RNA结构与核糖酶催化活性相关性规律及其反应机制[11]。在探索生命起源和RNA分子构效关系研究中,XAS技术已凸显出巨大价值,在未来探索RNA动态反应机制和规律研究中将会发挥更为重要的作用。
1.2 深海热液新陈代谢理论与XRS
46亿年(4.6 Ga)前的超新星大爆炸,诞生了太阳系。4.5 Ga期间,地球受系内行星撞击,产生了月球并致地球旋转和倾斜,带来日夜交替、季节变化、热力和能量分配[12]。早期地球处于缺氧还原环境,太阳光通量比现在大约低20%[13]。地球在4.550~4.033 Ga年间又受到大量小行星强烈轰击,使地球壳岩难以留存,生命难以诞生或延续。海洋约在4.3~4.4 Ga期间形成[14]。海洋的形成,为生命诞生提供了理想的温床。
深海热液新陈代谢理论认为,生命起源于深海热液系统硫化矿自养化学反应[5]。地球形成初期,中洋脊火山作用产生的热液喷口及其海床提供了约360~405 ℃的硫化物热液和生命所需能量,以及丰富的反应气体、溶解性元素、适宜的pH与温度条件和氧化还原梯度[4,12,15],这些物质和环境条件促生了单体有机物形成,并通过一系列自养反应和新陈代谢过程,形成了RNA分子和原始细胞,诞生了生命[4-5]。
两种学说的主要不同之处在于诞生生命所需的能量来源。有机分子合成理论认为RNA生物分子合成的能量源自空间紫外光照; 而深海热液学说认为生命依赖于H2和金属硫化物的催化作用,它通过乙酰辅酶A(AcCoA)路径还原CO2,由此生成微生物新陈代谢中的核心碳架主链硫酯,由于在H2和CO2合成乙酸盐和CH4时会释放能量,因此这一过程不需要来自空间的光照提供能量[15]。目前这两种学说都仍在不断地探索和研究发展中。
金属硫化物是深海热液新陈代谢理论中的关键化合物,它催化了生命前体化合物的合成[4-5]。热液中的一氧化碳和甲基硫通过FeS和NiS的催化反应形成了乙酰甲硫醚,同时也构建隔室内壁、防止反应产物扩散,形成了一个既可保有足够浓度,又具复制特性的RNA世界[4]。模拟实验和XANES分析证实,H2S和Fe(Ⅱ)在溶液条件下确可形成稳定的硫化矿物相[16]。广延X射线吸收精细结构(EXAFS)谱显示,四面体矿物相第一壳层含4个S原子、第二壳层含2个Fe原子,从而揭示了这种具有生命前体化合物催化性质的硫化矿物相的形成过程与转化机制,对深入理解生命起源与进化过程具有重要参考价值。
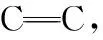
海洋热液体系中橄榄岩与水发生的蛇纹石化生氢作用,为产生初始非生物型甲烷及其碳氢化合物提供了所需H2源[15,21-22],是海洋热液生命体系中的重要地球化学反应。低温生氢模拟实验和XANES分析表明,在水岩作用早期阶段,含Fe(Ⅱ)矿物与溶液态Fe(Ⅱ)在100 ℃即可发生氧化反应,通过蛇纹石化作用产生溶解性H2、磁铁矿和低分子量有机酸[23]。在球粒状陨石硅酸盐样品脱氢反应研究中,通过XANES测定Fe3+/TFe(全铁)比值[24],发现微米与亚微米级层状硅酸盐呈现不同氧化还原态,亚微米级细粒层状硅酸盐含70%高比值Fe3+/TFe,微米级粗粒层状硅酸盐氧化度较小,仅为55%,揭示高速率生H2与存在高含量Fe3+密切相关,如以下化学反应所示:
3Fe+4H2O→Fe3O4+4H2
2FeO+H2O→Fe2O3+H2
1.3 生命进化过程与XRS
蛋白质信号传递是生命进化研究中的一个重要科学问题,研究表明,它与元素的氧化还原过程相关。
叶绿体传感激酶(CSK)是一种两组分信号传导变性感应组氨酸蛋白酶,可以为质体醌(PQ)与光合作用基因表达提供信号传递链,其响应和信号传递与激酶中的Fe形态转化相关。利用XAS研究CSK感知和监控PQ氧化还原过程[25],发现CSK中铁以Fe3+存在,EXAFS傅里叶变换后的主、次峰位和键长揭示CSK中存在Fe—S键,半胱氨酸中存在Fe—S配位和Fe—S—Fe键桥。这表明CSK是一种通过进化留存下来的氧化还原3Fe—4S簇响应体,其氧化还原态的改变导致了CSK蛋白构象变化,从而实现自激酶活性的响应调制和信号传递。
随着生物进化,生命与碳、氢、氮、氧、硫和磷六种基本元素及起酶催作用的过渡金属形成了紧密依存关系[26]。过渡金属在生物进化中常相互替代,但生命基本元素较少观察到此类现象。近期研究发现,盐单胞菌可在有砷而无磷条件下生长,出现生命基本元素P被As替代的现象,μXANES和μEXAFS分析显示,细胞中As为五价而非三价,第一壳层为四氧配位,第二壳层为As(Ⅴ)—O,As(Ⅴ)—C结构,而非As—Fe或As—S结构[26],表明盐单胞菌生长中不仅伴随着砷酸盐的吸收,而且还随之进入核酸、蛋白质、脂质及代谢物中,这一研究揭示了生物进化对环境的适应性和生物多样性。
在小行星撞击地球等重大事件后,地球及其承载生命的灾后恢复能力,是生物进化研究中的一个关键科学问题。基于IODP/ICDP国际科学钻探项目所获岩芯样品和XRF分析,对经历白垩纪小行星撞击后的生态恢复过程进行研究,发现表征生产力的Ba/Ti比在小行星撞击后30 kyr为1,撞击后100 kyr,Ba/Ti比增加到2.0[1],表明生产力恢复迅速,揭示生态进化过程本身才是生产力恢复的决定性因素。
动物组织结构、形态特征是生物进化和自然选择的结果。动物皮毛颜色可揭示年龄、雌雄、饮食特性,在自身伪装、交配、统建领地中发挥着重要作用。例如,生活在侏罗纪早期的狭翼鱼龙,与大多数当今仍栖息于远洋环境并呼吸空气的脊椎动物一样,一生多为黑色,它们的枝状载黑细胞可从生理上通过黑色素再分配调整肤色,过滤紫外线、隐蔽或在寒冷水温时保持体温稳定,这些特殊的形态和生理适应性调整,正是这些远洋四足动物为适应海洋生活而经历的必要生物进化过程[27]。通过XRF和XAS分析发现,这种褐黑素中存在Zn—S键[28],褐色素残余物由Zn有机硫络合物组成[29],两者都与真黑素形成内环配位络合物,与二醇或羧基功能团中的O/N配位[30],但真黑素不含硫基团,褐黑素需要含硫基半胱氨酸。
在3Ma前业已灭绝的哺乳动物化石中,μXRF显示Zn主要分布在毛发末梢,XAS揭示Zn与有机S形成了四面体络合物,构成了高浓度褐色素体,且Cu,Zn,S在动物表皮呈不均匀分布,在尾足部有机硫缺乏,这表明3 Ma年前,此类哺乳动物尚未进化到均匀着色阶段[31],且羽、毛的组分特征和不易降解特性,也应是自然界生物进化的结果。
采用μXRF和XANES分析晚侏罗纪150 Ma鸟类羽毛,在沉积物表层下鸟骨碎片中发现硫酸盐,与沉积物基质相比,硫醇在羽毛中浓度更高、硫酸盐更低; 羽毛中Cu与有机硫的非均匀分布相关,揭示两者间形成了具有杀菌特性的铜有机螯合物,从而抑制了羽毛角质蛋白中的有机硫降解,有效保存了羽毛[32]。羽毛中存在具抑菌作用的Cu有机螯合物,应是生物进化与自然选择的必然结果。这些研究也充分显现了XAS技术在发现并揭示生物进化机制与规律探索中的重要作用。
XAS在生命起源与进化研究中也面临一些挑战。例如在蛋白质信号传导机制EXAFS分析中,由于N、O散射特性相近,单一的EXAFS拟合不能区分O或N[25]。一些样品存在元素间干扰,如用XAS分析角质蛋白中Zn有机硫化物时,硫含量约为7%,而降解产物富含氧化性硫。随着埋葬降解,双硫键氧化、断裂,产生的硫K吸收谱背景会干扰原始外皮角质蛋白中较弱的杂环硫振荡谱,遮蔽了原生硫信号[28]。因此,在使用XAS技术开展生物进化过程与机制的研究中,还需进行深入的探索和多技术结合。
2 早期生命寻迹与X射线光谱分析技术
地球上早期生命可追溯至数十亿年前的远古时代。3.5 Ga是目前认为的最确切生命起点,3.8 Ga或更久远年代的发现亦有报道[33]。XAS技术在地球早期生命寻迹过程中也发挥了重要作用。
2.1 早期生命寻迹
地球上的早期生命迹象,主要在几处仍暴露于地表的古老稳定地块——太古宙克拉通地区发现,如澳大利亚和南非(3.2~3.5 Ga)、格林兰岛和加拿大(>3.5 Ga)等。
在澳大利亚,发现了2.97 Ga古生物膜和球型微生物[34],在3.20 Ga黑色硅质体和3.24 Ga黄铁矿中分别发现了铁厌氧菌[35]、丝状微生物[36]及有机质与元素碳、氮、硫等[37]。在3.47 Ga重晶石中报道发现了硫酸盐还原菌[38]和细胞壁降解产物[39]; 在南非,发现了3.20 Ga前一种由潮滩微生物蚀刻作用形成的微生物垫[40],在3.45 Ga沉积硅质岩中发现了与现代超嗜热菌相似的微生物,并可见细胞壁[41]; 在格林兰地区,发现了由光自养菌产生的3.7 Ga生物成因碳[42]及3.8 Ga层状石墨和磷灰石,石墨内含生物基本元素N,P和S,推测为生物变质作用产物[43]; 在加拿大东北部3.95 Ga变质沉积岩中,据报道发现了认为源于古生物有机质的石墨[44]。在魁北克3.77(U-Pb)~4.28(Sr-Nd)Ga NSB表壳带铁质层[33],发现了具有海水化学组分和重Fe同位素特征的微米级管状和丝状赤铁矿,与海洋热液沉积层微生物近似,认其为古微生物化石。
这些早期生命迹象的发现,对深入研究地球生命起源和进化具有重要意义。但由于年代久远,历经地质、环境巨变,使发现与确证面临重重困难。
2.2 早期生命质疑
对地球上已报道发现的早期生命迹象,目前的质疑主要包括三个方面:
首先,年代存疑。例如,关于加拿大东北部3.95 Ga泥岩石墨地质年代,有观点认为这些样品并没有作者声称的那么古老[45];
其次,成因存疑。早期生命以假定生命以碳基形式存在,且产生了碳同位素分馏为前提。相比13C,生物选择性地从化学键中捕获并释放轻同位素12C。在曾有微生物生存的地方,12C富集。例如3.95 Ga泥岩石墨δ13Corg=-28.2‰~-11.0‰,含较丰富轻碳同位素,并与层叠有机质近似; 碳酸盐δ13Ccarb与泥岩石墨δ13Corg相差达25%,呈显著分馏,故认为其源于有机质[44]。但碳同位素不是生物成因的唯一性判据,碳既可源于生物,也可源于非生物[46],受到了多种因素影响:
① 地质构造作用: 在澳大利亚4.252 Ga锆石中发现的钻石-石墨包裹体δ13低达-58‰,但其可能是早期地球构造循环作用的产物,而非生物成因[47];
② 变质流体作用: 格林兰3.8 Ga表壳铁质层石墨,结晶度不好,流体二次运移或高变质作用可导致非生物成因石墨生成[48-49];
③ 矿物分解作用: 菱铁矿在450 ℃可发生歧化反应,产生非生物成因石墨[46]:
6FeCO3→2Fe3O4+5CO2+C
④ 非生物合成作用: 实验室可合成非生物成因甲烷,所得碳同位素特征与生物成因相似[50],格林兰3.7 Ga石墨或是由非生物脱碳反应、或金属与CO的F-T合成反应产生[33,46]。
第三,方法存疑。例如,尽管激光拉曼可揭示保存指数,但非生命指标; 岩石粉碎、样品制备、非原位测定等,会引入污染,或致母岩缺失,较难获得准确年龄和成因[51]。
因此,不仅寻找早期生命的迹象十分困难,而且对早期生命的识别与确证也面临挑战。
2.3 早期生命质证与X射线光谱
碳成因所具有的双重性,导致任何单一方法都难以实现对地球上早期生命的确证,需要包括XRS在内的多学科、多技术、多方法的协同研究和多信息佐证。
2.3.1 生物成因识别
低δ13C值是早期生命存在的主要证据,但具有双重性而不具充要性; 铁、硫同位素生物和非生物成因分馏效应也与之相似。因此,在同位素分析的同时,探寻地质过程、伴生矿物、反应机制等的相互印证,就成为了辨识早期生命真伪的重要途径。在此过程中XAS技术显现了较显著的潜在应用价值。
(1)矿化机制。碳质进入沉积物,会经历氧化为碳酸盐、不溶性三价铁还原为可溶性亚铁等过程,并伴生含铁碳酸盐、亚铁硅酸盐和磁铁矿等矿物,生成莲花座型碳酸盐簇体:
这种特殊簇体可归因于微生物矿化作用[33]。μXRF和X射线计算机断层扫描(μXCT)既可准确测定碳酸盐组成,亦可测定岩石组构和空间结构[52],是生物矿化机制研究中的重要潜在技术手段和未来发展方向之一。
(2)物相特征。在加拿大东北部发现的3.95 Ga石墨尽管存在歧化反应可能,但并未发现歧化反应产物菱铁矿和磁铁矿; 同时,由共聚焦激光拉曼测得的物相结晶温度亦为(563±50) ℃,与母岩一致,表明石墨非源于后期污染; 再结合其低的δ13C值,故根据以上三点综合判断,此石墨为生物成因[44]。与此类似,格林兰3.7 Ga石墨与母岩结晶温度的一致性也成为了非后期变质作用产生的主要证据之一[53]。XRF可以定量测定菱铁矿[54],如结合XAS分析其中的Fe形态[55]、区分生物和非生物成因纳米磁铁矿[56],将可更准确厘定矿物反应过程与产物,佐证生物或非生物成因机制。
(3)矿物形貌。近现代热液生态体系下,沉积物中微生物可产生管状赤铁矿; 在加拿大3.77~4.28 Ga热液沉积物中观察到丝、柱状物。根据均变论,远古时期的管状和丝状赤铁矿也应是细菌包层和胞外细丝矿化残留,其氢氧化铁圆柱体由蓝藻菌细胞形成[33]。这种三维结构测定,正是μXRS技术的显著特点。例如,用μXRF扫描侏罗纪早期狭翼鱼龙化石[27],可获得早期矿化过程所保留的三维细胞形态,且元素微区分布表明,鱼龙表皮通过自生磷酸钙的部分复制形成了化石,促生了原位分子转化,有机分子转化为稳定大分子的地质聚合反应,使古生物获得了二次保存机会。μXRF不仅可以获得物质二维成分信息[57-58]、三维元素分布[59]及物质空间组构[52],还可以揭示古生物化石形成和保存机制[27],因此可为早期生命的识别与研究提供更丰富的多维数据信息。
(4)生命元素。磷等生命基本元素的存在,具有生物成因指示作用。海底碳质积累会通过生物代谢浓聚磷。在加拿大NSB表壳带含石墨碳酸盐中发现存在内生磷灰石,且与在古元古代晚期微生物生态系统中发现的磷灰石相似,表明该处石墨源于生物,揭示在3.77~4.28 Ga年前,生命即在海底热液喷口附近存在并栖息[33]。磷灰石和磷可采用XRF测定[60-62],EXAFS可揭示过渡金属与磷灰石纳米晶体的键合位与结构[63],如果我们将XRS应用于碳、磷等元素分布与形态分析,将有助于揭示生命元素间的相关性特征。
(5)地质构造。沉积物和碳酸盐间的不断键合和捕获形成的沉积叠层结构是早期生命存在的重要迹象。用XRF扫描格林兰3.7 Ga变质碳酸岩中圆拱和锥型部位,发现元素分布与沉积纹层结构相关,在特定层位K、Ti显著降低,Si升高,Ca不变; 稀土和镱元素分配模式与交叉沉积及波纹状角砾岩分布均揭示其形成于海洋沉积环境,推测为生物CO2埋存作用形成[64]。但这种沉积叠层结构是否由生物活动产生仍受到质疑。用μXRF扫描样品[65],确证元素呈梯度分布且Ca,K和Ti变化与前文[64]一致。但新观点认为[65],尽管元素呈叠层分布,但岩石从白云岩向石英质过渡时未发现分层残余物,岩石本身即由石英和富铁夹层组成; 同步辐射扫描显示,REE+Y分配模式并不只是源于白云岩,云母也是重要载体; 与碳酸盐相比,富含云母的硅酸盐样品REE+Y丰度更高,Eu正异常也更显著。综合主次元素二维分布和稀土元素分配模式,该研究认为所研究样品尽管源于海洋沉积环境,但碳并非生物成因。遗憾的是,该研究[65]尽管提及XANES,但未见详情报道。如能应用XAS原位分析元素形态,则应可更准确地质证其来源与成因。
2.3.2 生物功能团鉴定
有机微化石是早期生命的重要标识。采用聚焦离子束FIB获取亚微米无污染新鲜样本,通过微区XANES原位测定1.88 Ga年前冈弗林特有机微化石组成和结构[66],发现远古生命已具备了蛋白质合成能力,且历经1.88 Ga漫长地质过程仍可保存至今,极富挑战性:
(1)有机微化石中观察到丰富生物功能团: 在288.6,286.4和290.3 eV处观察到羧基[67-68]、C-N/S/O未饱和键和纳米OC-CaCO3峰[66,69-70],以及酰胺/亚胺/腈(401.4 eV)、芳香族N(399.8/398.8 eV)、钙/钾硝酸盐(401.7和405.4 eV)和吡咯(402.2 eV)等[71-72]。微化石中低含量N导致了XANES信噪比低[66],带来一定的分析困难;
(2)多样配位结构反应了复杂地质环境变化。不同N/C比和S-,N-,O-与芳香基结构,反应了不同的热成熟度、复杂的氧化还原过程及多样成岩与保存条件。热成熟度增加,XANES吸收谱峰由高能向低能漂移。高含量烯烃[73]和/或对苯醌等芳杂环功能团[74]将使芳香族和烯烃功能团C-XANES谱拓宽并由285.1向284.9 eV漂移; 酚基/羰基由286.7向286.4 eV漂移。研究表明,芳香结构中杂原子嵌入、氨基酸与酚基间的缩聚反应等,是出现这种漂移的主要原因[67-68];
(3)配体特征揭示了远古与现代藻类的相似性与进化变异。由XANES测得有机微化石N/C比为0.21和0.24,现代蓝藻和微藻为0.24和0.17,这表明存在源于蛋白质的酰胺基团,揭示其尽管经历了50度的成岩温度变化,但亚微米级化石保存了与现代蓝藻和微藻相似的C和N有机功能团。有机微化石中含有更高含量的芳香化合物和含氧基团,也揭示了生物进化前后的变异特性。
2.3.3 细胞形态分析
细胞空间结构和形态可揭示生物形成与地质过程的相互关系,从而为佐证生命迹象提供依据。法国阿尔卑斯变质沉积岩中225 Ma石松类大孢子化石,经历14 k bar和360 ℃高压变质作用及35 km俯冲造山运动,组构保存完好,孢壁呈有机质化学异质性。由FIB切得包含孢内壁和4~5 μm外壁的薄片,用μNEXAFS测定内、外壁C吸收谱[75-76],在285.1,292.8和286.7 eV处观察到芳香族/烯烃C、C—C键/芳香及酮基团; 在287.2,288.7和290.3 eV处,发现少量酚基、羧基、脂肪酸功能团; 研究表明外壁由芳香族化合物、烯烃及饱和烃组成,内壁为石墨型碳,有机基团与石墨共存,表明部分有机质经历变质作用而石墨化,但在经历高度变质作用后,生物特征并未完全抹去,其残存有机分子信息仍是生命存在的重要证据[76]。
在地球早期生命的研究中,尽管XRS已在生物成因识别、功能团鉴定及细胞形态分析中得到了成功应用,但更深入的开展XRS微区与形态原位分析,应是未来一段时期的发展方向。
3 全球气候变化与X射线光谱分析技术
全球气候变化是人类面临的重大科学问题,海洋碳循环是影响全球气候变化的重要因素。探索Fe的生物有效性和沉积碳汇机制,是提升海洋和内陆水系固碳能力的关键途径。
3.1 全球气候变化
随着工业化进程,大气CO2浓度一直呈上升趋势。与工业革命前相比,CO2已上升38%,到2100年,CO2还将增加20%[77],减少大气CO2刻不容缓。
3.1.1 海洋碳汇与海洋铁循环
海洋碳循环是影响全球气候变化的重要因素。海洋CO2储量是大气的60多倍[78],人类活动产生的四分之一CO2已被海洋吸收[77]。海洋浮游植物对大气CO2的光合吸收与沉淀作用,是海洋吸存大气CO2的主要机制,南大洋生物泵作用的强化导致了从间冰期向全冰期过渡时的CO2降低[79]。
海洋铁循环,是海洋碳循环的控制性因素,全球大洋50%的初级生产力受铁循环控制。铁在地壳中的丰度达5.6%,但除部分冷水海域外,Fe在海水中浓度极低(<10-15mol·L-1),导致在40%的海域,特别是在高硝酸盐、低叶绿素海域,Fe供给严重不足[80]。铁量不足,使通过光合作用大量吸收大气CO2的海洋浮游植物不能有效利用海洋中的氮和磷,导致海洋初级生产力下降,固碳能力减弱; 而海洋铁量上升,则可使冰期大气CO2浓度减少约25%[81]。海洋铁缺失,制约了海洋初级生产力和海洋碳汇能力[82]。
海洋中铁的存在形态,决定了生物可利用度和海洋初级生产力。海水中铁量不足的原因在于三价铁Fe(Ⅲ)的低溶解度[83]。海水中可溶性Fe(Ⅱ)浓度小于1 nmol[84]。以可溶态和胶体颗粒物存在的铁-有机络合物是海洋中的主要形态之一[85]。难溶性三价铁和铁有机络合物铁的存在,降低了海水中铁的生物可利用度,不利于海洋初级生产力的增长和碳汇能力的加强。
海洋铁源的时空分布与溶解性存在较大差异,对海洋铁循环产生了重要影响。铁既可源于海岸带和浅层沉积物、冰川、岛湖,也可源于火山、尘埃和生物燃烧。大气飘尘或气溶胶[83]是海洋中铁的主要来源之一[86]。在冰川尘粒、燃烧颗粒物和干旱土壤三类大气飘尘颗粒物中,铁的可溶性差异达三个数量级[87]。大气飘尘在长距离(~1 500 km)转运中,约有一半会沉积于海洋中,颗粒物中所含铁的矿物组成、物相和形态等,一定程度上控制了其生物可利用性和大洋初级生产力[82]。此外,除了非生物性化学反应影响,海洋铁循环还会受微生物调控[88],其在碳铁循环中的作用亦不可忽视。
3.1.2 内陆水系碳汇与铁有机络合物
内陆水系是主要碳源之一。目前内陆水系CO2已处于过饱和状态,每年向大气排放2.1 Pg C yr-1的CO2[89],河流释放的CO2比湖泊和水库总排放量高约5倍[90]。陆生碳库中约有一半通过内陆水系进入海洋[90]。
另一方面,内陆水系也发挥着重要碳汇作用。它会将CO2吸埋于水系沉积物中,且吸埋速率比海洋快。内陆湖泊、水库每年可吸埋有机碳约0.15 Pg C yr-1,其中水库吸埋占其中的40%[91]。
沉积有机碳汇是全球碳库的重要组成部分。全球海洋与水系沉积物中,超过15%的有机碳(OC)通过与活性铁结合形成稳定的铁-有机碳(OC-Fe)络合物,并在沉积物中长期封存。沉积型有机碳约占全球铁相关有机碳库的21.5%,其中,总有机碳中23%~27%保持着与活性铁键合,与铁相关的有机碳储量是大气碳量的2 900~6 800倍[92]。
3.1.3 多元素体系对碳铁循环的潜在影响
铁的存在形态受共存元素的影响。化石燃料产生的含铁尘埃对部分海洋表水溶解铁的贡献率为70%~85%,并与黑炭、V、Ni、Al等元素存在相互影响[93]。Fe与C、N、O、S的氧化还原反应在驱动全球生物地球化学循环的同时,也受到各种氧化物、氮化物、碳化物、硫化物及共存过渡金属元素的调控[88],例如:
2Fe2++MnO2+2H2O→Mn2++2FeOOH+2H+
与Fe相比,Cu的高氧化还原电位及生物可获性使其从碳代谢物中可提取更多能量[94]; Cu与有机配体的键合力也高于Fe[95]; 决定海洋初级生产力的海洋浮游植物生长需要有效利用海洋中的氮,而三种最重要的典型固氮酶在活性反应位除Fe外,还含有V和Mo[96]。因此,在海洋与内陆水系碳循环研究中,探索多元素体系下的相互关系与作用,十分必要。
3.2 全球气候变化研究中的科学问题
海洋浮游生物对大气颗粒物中Fe的吸收途径及其Fe形态转化机制,是全球气候变化研究中的一个关键科学问题。目前的主要观点包括:
①光解作用下,Fe(Ⅲ)与铁载体络合物分解,Fe(Ⅲ)还原为生物可吸收态Fe(Ⅱ)[97];
②还原酶作用下,Fe(Ⅲ)-铁载体在吸收前被还原为过渡态Fe(Ⅱ)s,实现跨植膜运输[98];
③生物调控作用下,异养和自养菌类原核生物从铁载体获取铁,硅藻类真核生物将溶液中溶解性Fe(Ⅲ)′还原为Fe(Ⅱ)后吸收[98],兼养浮游生物直接吸收胶体铁[85];
④矿物相作用下,大气飘尘中亚铁矿物含量决定了Fe的海洋浮游生物可利用度[99]。
因此,对海洋碳循环具有决定性影响作用的海洋铁的生物利用机制还需要深入探索。
铁-有机络合物精细结构是碳汇研究中的一个重要科学问题。有研究认为,OC-Fe与细粒粘土矿物量正相关; 但也有研究表明,反应铁相并未提供充足表面用于吸附,而是铁氧化物中的铁为溶解性有机大分子提供了具有内聚作用的共价键,通过粘土和金属氧化物吸附[92]、共沉淀或直接螯合[100],将碳封存于沉积物中,目前,关于OC-Fe的准确配位结构也还没有取得一致性清晰辨识[101]。
探索Fe-OC构效关系对厘清沉积碳汇机制具有重要科学意义。目前关于OC-Fe络合物的形成速率、固化过程和构效关系等还不十分明了[82]; OC-Fe在早期成岩和长期埋藏过程中的性质、演化和元素与矿物间的相互作用等科学问题,还需进一步研究[102]; 关于OC-Fe分解所导致的沉积有机碳汇向碳源转化的潜在可能风险、趋势与份额等,也亟待进行深入探索。
3.3 全球气候变化研究中的XRS分析技术
海洋铁含量极低,只有10-9~10-12mol·L-1 [82],甚至低至10-15mol·L-1 [80],在可以测定海洋低铁量之前,人们还不能认清海洋中碳循环本质。随着分析技术进步,分析灵敏度提高,人类才进行了海洋中nmol·L-1级铁量的测定,并由此发现了铁含量是海洋碳循环中控制因素这一重要科学规律,促进了海洋铁限学说和海洋碳铁循环理论的建立[82]。在此基础上进行的大洋施铁实验,显著提高了海洋初级生产力。没有分析科学技术的进步,就没有海洋铁限学说的形成。
3.3.1 海洋铁源生物有效性
大气飘尘是重要海洋铁源,铁形态是生物有效性的决定性因素。飘尘中Fe的生物有效性测定,如采用总体评估手段如淋滤实验,则所测溶解性Fe主要为胶体[103],而非真实溶解态Fe[99]。而采用μXAS直接测定大气飘尘中矿物和元素形态,则更为准确、可靠。μXAS研究发现,源自干旱地区飘尘中的可溶性铁量最小(<1%),主要由含Fe(Ⅲ)的水铁矿、针铁矿和赤铁矿组成; 冰川尘粒含可溶性铁量略大(2%~3%),主要为混合矿物相; 油品燃烧颗粒物中可溶性铁最高,可达77%~81%,且以易溶Fe(Ⅲ)存在[87]。
有研究认为,蓝藻通过细胞表面过程增加溶解速率来实现对颗粒物中铁及其氧化物的利用[104]。但也有研究发现[99],与非冰川物质相比,采用冰川颗粒物培养的海洋硅藻生长更迅速,并具有更高光合作用效率。XAS数据显示,冰川颗粒物中所含原生Fe(Ⅱ)硅酸盐矿物是产生这一结果的原因,大气飘尘的生物可利用度由其所含Fe(Ⅱ)原生矿物量控制。该研究也揭示,冷冰期大气飘尘给海洋带来了生物有效性更高的铁源,从而增加了冷冰期海洋碳汇能力。
3.3.2 铁有机络合物构效关系

铁-有机质(Fe-OM)络合物的形成过程和存在形式对结构稳定性具有显著影响。EXAFS分析表明,C/(C+Fe)比值变化会影响Fe—C结构和稳定性及Fe和C循环[106]; 因形成不溶性Fe(Ⅲ)-OM络合物,使得经共沉淀途径形成的Fe-OM可沉积更大量的碳,而吸附型沉积碳量较低。更为重要的是,与吸附机制相比,经共沉淀机制生成的Fe-OM解吸率更低[107],有利于碳储存。
3.3.3 多元素体系
XRS技术在碳铁生物地球化学循环多元素体系研究中也发挥了重要作用。例如,C,N和Fe的EXAFS谱表明,氨基酸糖作为细胞壁中的重要N源,在富有机质土壤条件下,C和N脂类化合物和蛋白质会在Fe氧化物或氢氧化物表面富集,揭示微生物C和N循环与羟基化Fe氧化物密切相关[108]。海洋泻湖是海洋潜在铁源之一,XAS分析发现,泻湖沉积物中有机碳浓度会影响Fe,Ni和S的存在形态和矿物相组成[109]。
海洋Fe和相关性元素及沉积型有机碳在影响全球碳循环和气候变化的同时,气候变化也会通过影响元素存在形态进而影响有机质的稳定性和碳循环。例如,采集夏威夷山坡底土分析其中有机质,结合XANES谱,发现Fe和Al可稳定富N有机质; 降雨量越多,还原态Fe越多; 在还原性气候条件下,Al对有机质的稳定性影响比Fe更具重要性; 从而揭示气候变化对有机碳稳定性也产生了显著性影响[110]。
4 结论与展望
自太阳系诞生、地球形成,生命即开始了从无机向有机分子合成并最终形成RNA世界的漫长旅程。在这之中,有机碳和海洋热液体系发挥了至关重要的温床孕育作用。而μXRS因其独特的元素微区与形态原位分析功能,在研究RNA有机合成、海洋热液体系硫化矿自养化学反应过程,探索酶活响应与蛋白质信号传递途径,揭示海洋生物地球化学循环机制,发现生命起源与进化中的氧化还原规律等方面,发挥了至关重要的支撑作用,呈现出广阔的应用前景。
地球上生命起源于何时,目前还没有取得共识,但对于生命起源与进化的探寻,一直是人类孜孜以求的科学目标。要寻找地球上的最早生命迹象,首先需要确定识别生命是否存在的依据。目前碳同位素比值是生物成因的主要判据,但非充要性条件。因此,寻找相关科学证据即成为必然。XRS不仅可以通过伴生矿物、共存元素、分配模式等的测定分析,提供生物成因证据,而且还可以通过测定元素存在形态、有机功能团结构和细胞壁组成等,提供早期生命的形成环境、保存条件和成因机理等关键信息,成为早期生命探寻研究中不可或缺的重要手段。
海洋中可被生物利用的铁量不足,导致海洋初级生产力下降,固碳能力减弱。目前海洋浮游植物作用下Fe的形态转化途径与利用机制、海藻扩散与接触界面间的元素氧化还原过程等还没有获得全面准确认识; 铁有机质结构稳定性是影响沉积碳汇能力的关键因素,但铁有机络合物的准确配位还没有清晰辨识。深入开展XRS微区、原位、活体分析技术与方法研究,准确测定铁源物相和元素赋存状态,揭示铁有机质构效关系及沉积碳储机制,应是未来的重点发展方向之一。
X射线光谱分析技术不仅在生命起源、地球早期生命、全球气候变化等重大科学问题的探索中取得了重大研究进展,在冶金、材料、地质、文物、工矿、生态环境与生命科学等领域中,XRS也得到了广泛应用,其无损、原位、活体分析特性,可获得物质组分三维微区分布与元素形态信息的巨大应用潜力,在未来科学技术的探索研究中必将发挥更加重要的作用。
致谢:2019年《光谱学与光谱分析》编委会期间,受孟广政社长之托,组织国内学者撰写X射线光谱分析研究进展评述专辑。经过一年多的努力,专辑终于得以完稿。在此特别感谢孟社长对我国X射线光谱分析一直以来的关心和支持! 感谢广大科研工作者的持久关注,同时也特别感谢参与本专辑撰写的各位学者。
猜你喜欢
今日农业(2022年14期)2022-09-15
天天爱科学(2022年9期)2022-09-15
天天爱科学(2022年4期)2022-05-23
当代水产(2022年3期)2022-04-26
航空世界(2020年10期)2020-01-19
当代陕西(2019年23期)2020-01-06
琴童(2017年7期)2017-07-31
小学科学(2017年5期)2017-05-26
山东青年(2016年2期)2016-02-28
环境与生活(2016年6期)2016-02-27
