
高能饲粮对育成期蛋鸡肝脏miRNA表达谱的影响
2021-03-03 03:30王星果关树勇孙平江胡玉萍窦套存李永峰王克华
动物营养学报 2021年2期
王星果 卢 建,2* 关树勇 曲 亮 孙平江 郭 军 胡玉萍 窦套存 马 猛 李永峰 王克华**
(1.江苏省家禽科学研究所,扬州 225125;2.南京农业大学,南京 210095;3.江苏峪口禽业有限公司,宿迁 223733)
随着我国蛋鸡市场日趋饱和,蛋鸡业正在从数量增长模式向质量提高模式转变,这就需要对蛋鸡的生产性能作进一步改善。影响蛋鸡生产性能的因素复杂,其中营养水平是一个很重要的因素,营养水平过低或过高都会影响蛋鸡的健康,降低蛋鸡的生产效益[1]。对蛋鸡的营养调控可以从能量[2]、蛋白质[3]、维生素[4]和矿物质[5]水平等方面进行,其中能量水平的调控以往不够重视,但随着科学研究的深入,人们越来越认识到能量水平对蛋鸡的生长发育及生产性能有着重要影响[6]。再加上对节约饲粮成本的考虑,人们更加重视能量水平对蛋鸡生长和生产的调控[7],其中对育成期蛋鸡能量水平进行调控是其中的一个方向。
肝脏作为鸡物质代谢特别是脂质代谢的主要器官,摄入的能量物质主要在肝脏中进行代谢,肝脏对育成期蛋鸡的能量调控起着关键作用。miRNA是近年来研究较热的一类小分子物质,其通过靶向调控功能基因的表达进而调控各种生物过程,在肝脏中也发挥了重要作用,特别是对肝脏代谢[8-10]。因此,研究鸡肝脏miRNA特别是育成期蛋鸡能量调控后肝脏miRNA具有重要意义,且目前尚无其表达谱的相关研究,对此进行研究很有必要。有关育成期蛋鸡能量调控的研究集中在对调控后蛋鸡的表观特征进行阐述,如吕秋凤等[11]用4种不同能量水平的饲粮饲喂育成期蛋鸡,结果发现高能组可显著提高蛋鸡的体重;张李俊等[12]用3种不同能量水平的饲粮饲喂育成期蛋鸡,结果发现中高能组蛋鸡的体重显著高于低能组,产蛋率也高于低能组;李娜等[13]用5种不同能量水平的饲粮饲喂育成期蛋鸡,结果发现次高能组蛋鸡的体重和屠宰性能均显著高于低能组。研究也发现,miRNA在鸡肝脏代谢中发挥了重要作用,如miR-122通过负调控靶基因脂肪酸结合蛋白5(FABP5)、脯氨酰羟化酶alpha亚基(P4HA1)和泛酰巯基乙胺酶1(VNN1)等对肝脏代谢起调控作用[14-16];miR-33通过靶向抑制基因脂肪量和肥胖相关蛋白(FTO)、肉毒碱氧辛基转移酶(CROT)等的表达进而对肝脏代谢起调控作用[17-18]。有关鸡miRNA表达谱的研究很少,如性成熟启动前后下丘脑miRNA表达谱、皮质酮处理的雏鸡法氏囊miRNA表达谱、经选育的肉鸡胸肌miRNA表达谱研究[19-21]。
本课题组前期研究了育成期能量摄入量对蛋鸡体重和产蛋性能的影响,使用4种不同能量水平饲粮饲喂育成期蛋鸡,发现12~21周龄时不同能量组之间蛋鸡的体重始终保持极显著差异,高能组>中高能组>中低能组>低能组;能量水平越高,开产日龄越早,低能组、中低能组和中高能组集中在22周龄开产,高能组在21周龄开产,且高能组的开产日龄极显著早于其他各组;不同能量组43周龄时的产蛋数也有差异,高能组显著高于其他各组;但尚未进行miRNA相关研究。本研究选取高能组和低能组饲粮饲喂育成期蛋鸡,并对其肝脏miRNA进行高通量测序,分析其表达谱,对其靶基因进行预测和功能研究,为探明肝脏miRNA在高能饲粮对蛋鸡生产性状的调控中所起的作用,进而为改善蛋鸡生产性状打下基础。
1 材料与方法
1.1 试验设计与饲养管理
以江苏省家禽科学研究所选育的如皋黄鸡蛋鸡为试验动物。选取体重相近的9周龄母鸡160只,随机分成2个组,每组8个重复,每个重复10只鸡,分别定量饲喂代谢能水平为10.88(低能组)和12.14 MJ/kg(高能组)的饲粮。前8周常规饲养,18周龄后自由采食相同营养水平的标准饲粮,试验饲粮组成及营养水平见表1,以粉料形式分9~18周龄和19~20周龄2个阶段配制及饲喂。蛋鸡的采食量、能量摄入量及体重见表2,以平均值表示,使用SPSS 19.0软件中的t检验进行显著性分析。9~16周龄在育成鸡舍4层阶梯笼内饲养(5只/笼),17周龄起在产蛋鸡舍3层阶梯笼内饲养(1只/笼)。整个试验期间,所有鸡只自由饮水,常规光照和免疫。

表1 试验饲粮组成及营养水平(风干基础)
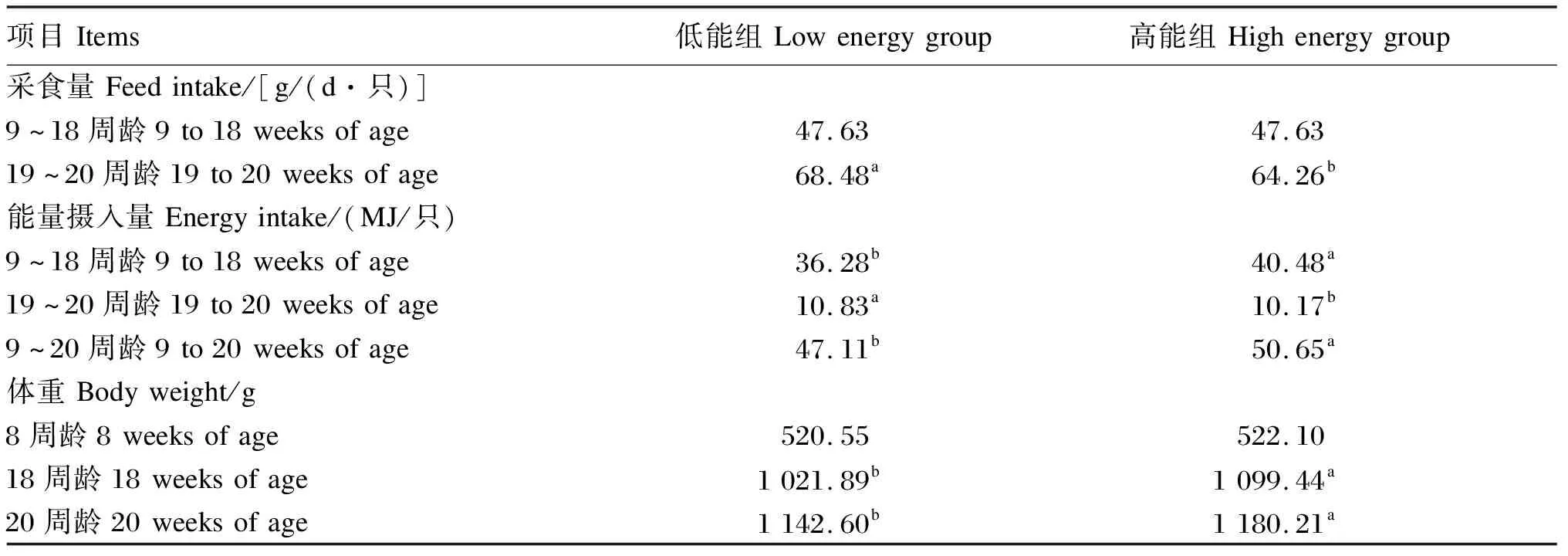
表2 蛋鸡的采食量、能量摄入量及体重
1.2 样品采集与small RNA测序
于20周龄时,每个重复随机选取1只蛋鸡,称重后每组选取最接近平均体重的3只,屠宰后取肝脏组织,放入液氮保存。
采用TRIzol提取肝脏组织总RNA,使用Agilent 2100毛细管电泳检测RNA质量,采用Illumina的Truseq small RNA样品准备试剂盒构建small RNA文库,PCR扩增富集,加上测序接头和Index部分,并通过凝胶电泳纯化,利用Quant-iT PicoGreen dsDNA Assay Kit对文库进行定量,Illumina Hiseq平台上机测序。
1.3 miRNA序列分析
测序得到的原始数据去除接头后进行质量过滤,得到纯净序列,对序列长度在18 nt以上的纯净序列进行去重处理后,得到唯一序列,使用Bowtie程序将纯净序列和唯一序列与鸡基因组比对,能比对上的序列进行后续分析。
将后续分析序列与miRBase中的已知miRNA成熟序列和前体序列进行比对,对比对上的序列进行注释并以各相应成熟miRNA为单位统计表达量。剩余序列与鸡基因组中其余非编码RNA(rRNA、snRNA、snoRNA、tRNA)进行比对并注释。
对注释到已知miRNA的序列表达量进行标准化,采用DESeq分析表达差异miRNA,按照表达量倍数差异(fold change)>2.0或<0.5和表达差异显著性P<0.05筛选出差异表达miRNA。根据前期鉴定的高能组和低能组肝脏差异表达基因,以高能组上调miRNA对应下调基因,下调miRNA对应上调基因,以差异表达基因mRNA的3′UTR序列为目标序列,应用miRanda软件对差异表达miRNA进行靶基因和靶位点预测。采用超几何分布对差异表达miRNA的靶基因进行基因本体论(gene ontology,GO)功能和京都基因与基因组百科全书(kyoto encyclopedia of genes and genomes,KEGG)信号通路富集分析。
2 结 果
2.1 蛋鸡肝脏small RNA测序质量分析
2组蛋鸡肝脏small RNA经过高通量测序,得到原始序列,结果见表3。从表中可以看出,6个样品测得的原始序列均在千万以上,而纯净序列/原始序列很高,集中在83%~89%,说明低质量的序列较少,测序效果较好。高能组与低能组相比,原始序列和纯净序列的数量均相当,说明2组测序质量相近。

表3 测序数据初步分析
2.2 蛋鸡肝脏miRNA鉴定
将各样品纯净序列去重后的唯一序列与鸡参考基因组比对,比对上的序列再与miRBase中的miRNA及前体作比对,对其进行注释,得到每个样品注释到的miRNA及前体,结果见表4。从表中可以看出,6个样品能注释到的miRNA均超过300个,占miRBase中已鉴定的鸡miRNA的近1/3。从前体结果可见每个样品鉴定出的miRNA有少部分定位在同一个前体上。各样品注释到miRNA的纯净序列占全部纯净序列的比例都在50%左右。表5显示了注释到各非编码RNA的纯净序列情况,几种主要的非编码RNA中,miRNA比其他非编码RNA多1~3个数量级。以上结果表明,miRNA在鸡肝脏中数量及含量都很多,说明其具有重要的作用。

表4 miRNA数据统计

表5 非编码RNA纯净序列统计
2.3 高能饲粮对蛋鸡肝脏miRNA表达量的影响
将高能组和低能组样品中鉴定出的miRNA表达量进行比较分析,筛选出差异表达miRNA,结果见表6。从表中可以看出,与低能组相比,高能组有2个上调miRNA(miR-1684a-3p和miR-1682),6个下调miRNA(miR-15a、miR-203a、miR-144-5p、miR-19b-3p、miR-29c-3p和miR-29a-3p)。其中miR-1684a-3p差异最明显,相差了200多倍,而miR-1682在低能组中未检测出,其可能在高能组具有特殊作用。
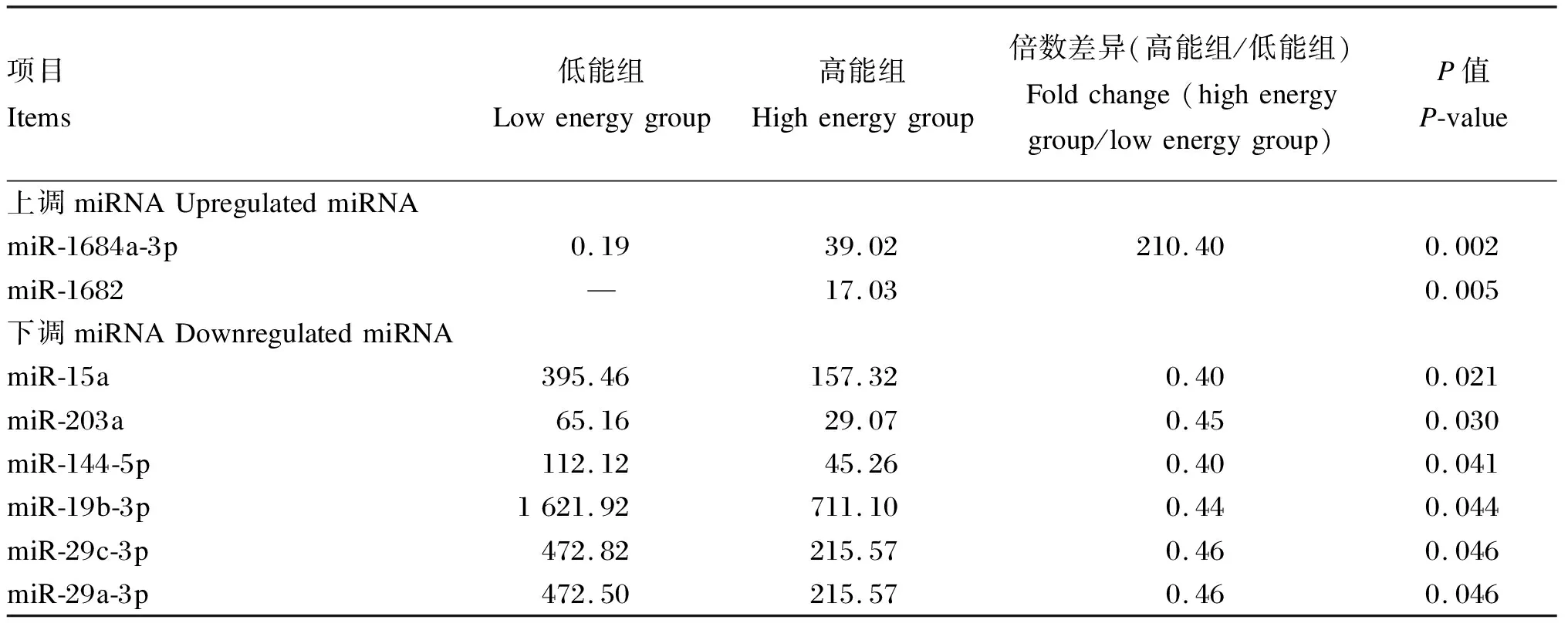
表6 差异表达miRNA
2.4 肝脏差异表达miRNA靶基因预测
课题组前期鉴定出高能组和低能组肝脏差异表达基因188个,其中高能组上调82个,下调106个,对高能组和低能组差异表达的8个miRNA运用miRanda进行差异表达基因的靶基因预测,共预测出41个潜在靶基因,结果见表7。由表可以看出,其中miR-15a预测出的靶基因最多,为26个,而miR-1682预测出的靶基因最少,为3个。miR-29c-3p与miR-29a-3p由于只有末尾差1个碱基,其种子序列相同,故其预测靶基因也相同。
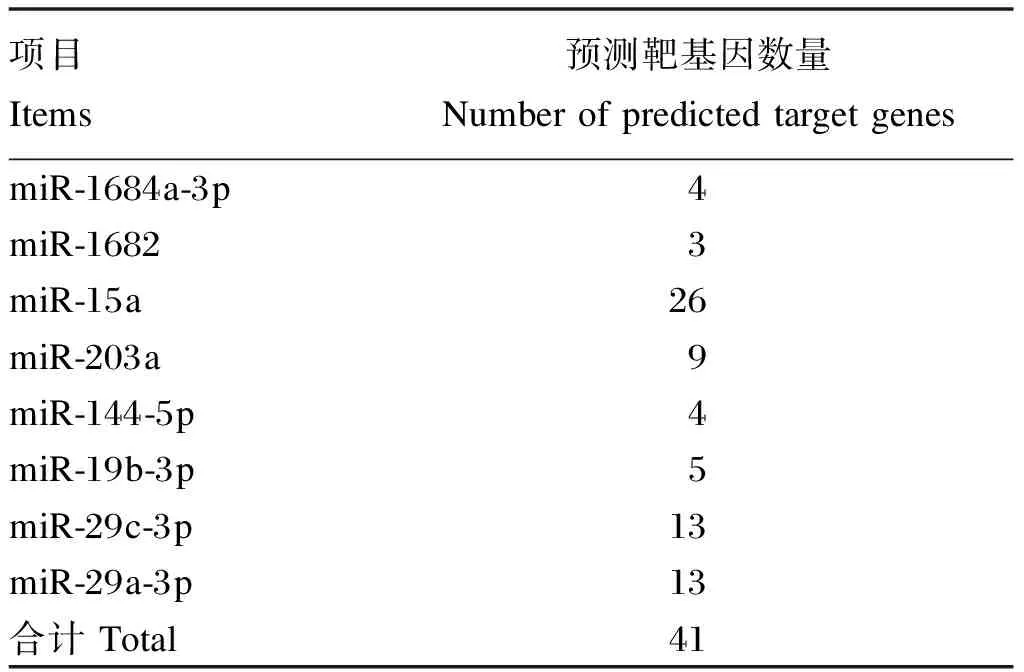
表7 差异表达miRNA靶基因预测
2.5 肝脏差异表达miRNA靶基因功能分析
2.5.1 GO功能富集分析
对高能组和低能组差异表达miRNA的预测差异表达靶基因进行GO功能富集分析,发现有192个显著富集的GO功能,其中147个富集在生物过程(biological process, BP),11个富集在细胞组分(cellular component, CC),36个富集在分子功能(molecular function, MF)。图1显示了富集最显著的20个GO功能,其中富集较多的是免疫系统过程正调控和分子功能正调控,涉及的基因有整合素亚基beta3(ITGB3)、RAB7B RAS癌基因家族成员(RAB7B)、甘露糖结合凝集素(MBL)、胰岛素样生长因子结合蛋白2(IGFBP2)、淋巴细胞抗原96(LY96)(免疫系统过程正调控)和RAB7B、鸟苷酸环化酶激活物1A(GUCA1A)、RasGEF结构域家族成员1C(RASGEF1C)、G蛋白信号调节因子6(RGS6)、印度hedgehog信号分子(IHH)、ARFGEF家族成员3(ARFGEF3)、高迁移率组核小体结合结构域3(HMGN3)、谷氧还蛋白(GLRX)(分子功能正调控),分别是5和8个,占总差异表达基因的12.2%和19.5%,说明它们是较为重要的GO功能。
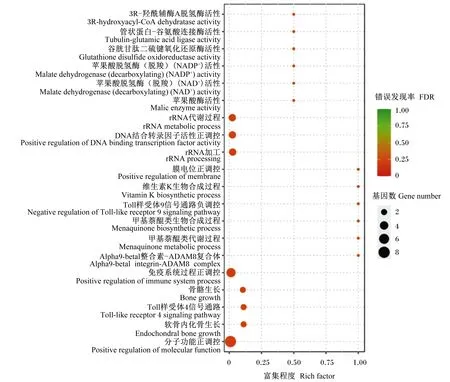
图1 差异表达miRNA预测差异表达靶基因GO富集分析:富集最显著的20个功能
2.5.2 KEGG信号通路富集分析
对高能组和低能组差异表达miRNA的预测差异表达靶基因进行KEGG信号通路富集分析,发现有4个显著富集的KEGG信号通路。表8显示了显著富集的KEGG信号通路及其涉及的基因,其中脂肪酸延长、非不饱和脂肪酸生物合成与脂质代谢相关,吞噬体与免疫相关,光转导与视觉相关,提示差异表达miRNA可能影响这几个信号通路。

表8 差异表达miRNA预测差异表达靶基因KEGG富集分析
3 讨 论
测序是现今运用广泛的组学技术,且随着技术的不断进步,数据的容量和质量都有了很大提高,现在常用的small RNA测序数据量一般都在一千万以上原始序列[19,22],本研究small RNA测序所得原始序列就在10 000 000以上。同时本研究的测序数据质量较高,能比对上miRNA的序列比例较高,且比其他非编码RNA所占比例多1~3个数量级。结果表明,本研究miRNA测序所得数据质量较高,后期分析结果较准确,置信度更高。
在对差异表达miRNA进行分析的过程中,本试验发现上调的2个miRNA(miR-1682和miR-1684a-3p)在低能组无表达或痕量表达,提示它们在能量摄入量对肝脏的影响中发挥了很重要的作用,而这2个miRNA暂无功能研究报道,因此有必要进行更深入的研究。
使用本课题组前期鉴定的低能组和高能组肝脏差异表达基因作为候选基因进行差异表达miRNA的靶基因预测,可将有效靶基因的范围大大缩小,阳性率也更高,其他研究者也有运用此方法的良好例子(差异表达miRNA与差异表达基因联合分析)[23-26]。miR-15a的预测靶基因数最多,提示其可能在能量摄入量对肝脏的影响中作用更明显。
在GO功能富集分析中,富集较多的主要是与免疫和分子功能调控相关的功能,说明miRNA是通过调控免疫和分子功能相关的基因进而调控这些过程的。之前本课题组分析育成期高能和低能饲粮饲喂的蛋鸡肝脏中差异表达基因的GO功能,也发现富集较多的有免疫相关功能,提示免疫相关过程受miRNA调控更多。由此表明,能量摄入量能直接或间接影响蛋鸡的免疫功能。
在KEGG信号通路富集分析中,也发现了与免疫相关的信号通路显著富集——吞噬体,吞噬体在动物细胞内起到抗原呈递、降解以及激活免疫的作用,是免疫过程中十分重要的环节[27-28]。吞噬体也能受到外界环境的影响,如洋葱伯克氏菌感染能延迟吞噬体成熟[29]。推测能量摄入量的差异通过影响miRNA的表达进而影响蛋鸡肝脏的吞噬体等免疫相关信号通路。除免疫相关信号通路,本试验还发现脂质代谢相关信号通路显著富集,这与Agrawal等[30]使用高能饲粮饲喂奶牛,KEGG分析发现血液白细胞中差异表达基因显著富集在不饱和脂肪酸氧化磷酸化以及Toll样受体信号通路的结果类似。这提示miRNA主要影响免疫和脂质代谢相关信号通路。
4 结 论
综上所述,对育成期蛋鸡进行高能饲粮饲喂可通过调控miR-15a等几个miRNA上调或下调肝脏中免疫和脂质代谢相关靶基因的表达,最终调控蛋鸡的免疫和脂质代谢,并影响生产性能。
猜你喜欢
今日农业(2022年2期)2022-11-16
动物营养学报(2022年8期)2022-09-01
小哥白尼(军事科学)(2022年4期)2022-07-08
今日农业(2021年7期)2021-11-27
中国饲料(2021年17期)2021-11-02
意林彩版(2020年3期)2020-11-22
大众电视(蓝天下)(2018年8期)2018-10-26
动物营养学报(2015年9期)2016-01-07
少儿科学周刊·少年版(2015年11期)2015-12-17
少儿科学周刊·儿童版(2015年11期)2015-12-17
