
乳酸锌对断奶肉兔生长性能、肠道发育和血清生化指标的影响
2021-03-03 03:30俞成浩郭志强李丛艳任永军梅秀丽谢晓红邝良德
动物营养学报 2021年2期
俞成浩 郭志强 雷 岷 李丛艳 任永军 郑 洁 梅秀丽 杨 锐 谢晓红 邝良德*
(1.四川省畜牧科学研究院,成都 610066;2.西南民族大学生命科学与技术学院,成都 610041;3.四川省动物遗传育种重点实验室,成都 610066)
锌作为所有动物必需的微量元素,广泛参与包括生长发育、能量代谢和蛋白质合成等各种生物过程,是动物体内300多种酶的组成成分或激活剂。由于锌的重要作用,锌缺乏会影响细胞分裂,抑制动物的生长发育,因此,在饲粮中添加锌用以提高生长性能已成为畜牧业中的普遍做法[1]。随着中国兔养殖业的迅速发展,对家兔饲粮营养需求的研究也日益深入。目前家兔饲粮中应用较为广泛的锌源依旧是无机锌(如硫酸锌等),但无机锌吸收利用率低,对环境污染严重。有机锌源由于其消化率和生物利用度均优于无机锌源,并且提高家兔生长性能和减少向环境排泄的锌量,而被越来越多的使用[2]。尽管近年来众多国内外学者对于有机锌的研究报道很多,但由于研究中有机锌的种类、添加水平、试验饲粮组成、动物生理阶段和品种等都不尽相同,从而得出的结论也有很大差异。杨翠军等[3]研究表明,饲粮中添加60 mg/kg硫酸锌能提高生长期獭兔的生长性能、血清总蛋白含量及谷丙转氨酶、谷草转氨酶和碱性磷酸酶活性等。杨国雨等[4]研究表明,饲粮添加40~120 mg/kg硫酸锌对生长獭兔的被毛长度、氮利用、皮张面积等有显著影响,但是对平均日增重、料重比和全净膛率等均无显著影响。任战军等[5]研究表明,饲粮中添加70~140 mg/kg蛋氨酸锌能提高幼龄艾哥兔的生长性能与组织锌沉积。因此,关于有机锌的作用效果仍有待于进一步讨论。目前关于乳酸锌的研究多集中在猪、鸡和鱼等动物上,其在兔上的研究还鲜有报道。本试验旨在研究乳酸锌对断奶新西兰兔生长性能、肠道发育和血清生化指标的影响,以期为乳酸锌在肉兔生产实践中的合理使用提供理论依据。
1 材料与方法
1.1 试验动物
试验于2019年6月—2019年7月在四川省畜牧科学研究院肉兔科研基地进行。选用健康、体重相近的28日龄同期断奶新西兰兔。
1.2 试验设计及饲养管理
选择28日龄同期断奶新西兰兔420只,随机分为5组:对照组(饲喂基础饲粮,饲粮中添加硫酸锌形式的锌0 mg/kg)、硫酸锌组(在基础饲粮中添加硫酸锌形式的锌80 mg/kg)以及3个乳酸锌组(在基础饲粮中分别添加乳酸锌形式的锌20、40、80 mg/kg),每组14个重复,每个重复6只(公母各占1/2)。试验期共49 d,其中预试期7 d,正试期42 d。试验采用3层阶梯式笼养方式,每笼2只兔(1公1母),颗粒料投喂,早、晚各饲喂1次,自然光照、自由采食、自由饮水。
1.3 试验饲粮
参照NRC(1977)标准配制肉兔基础饲粮,其组成及营养水平见表1。
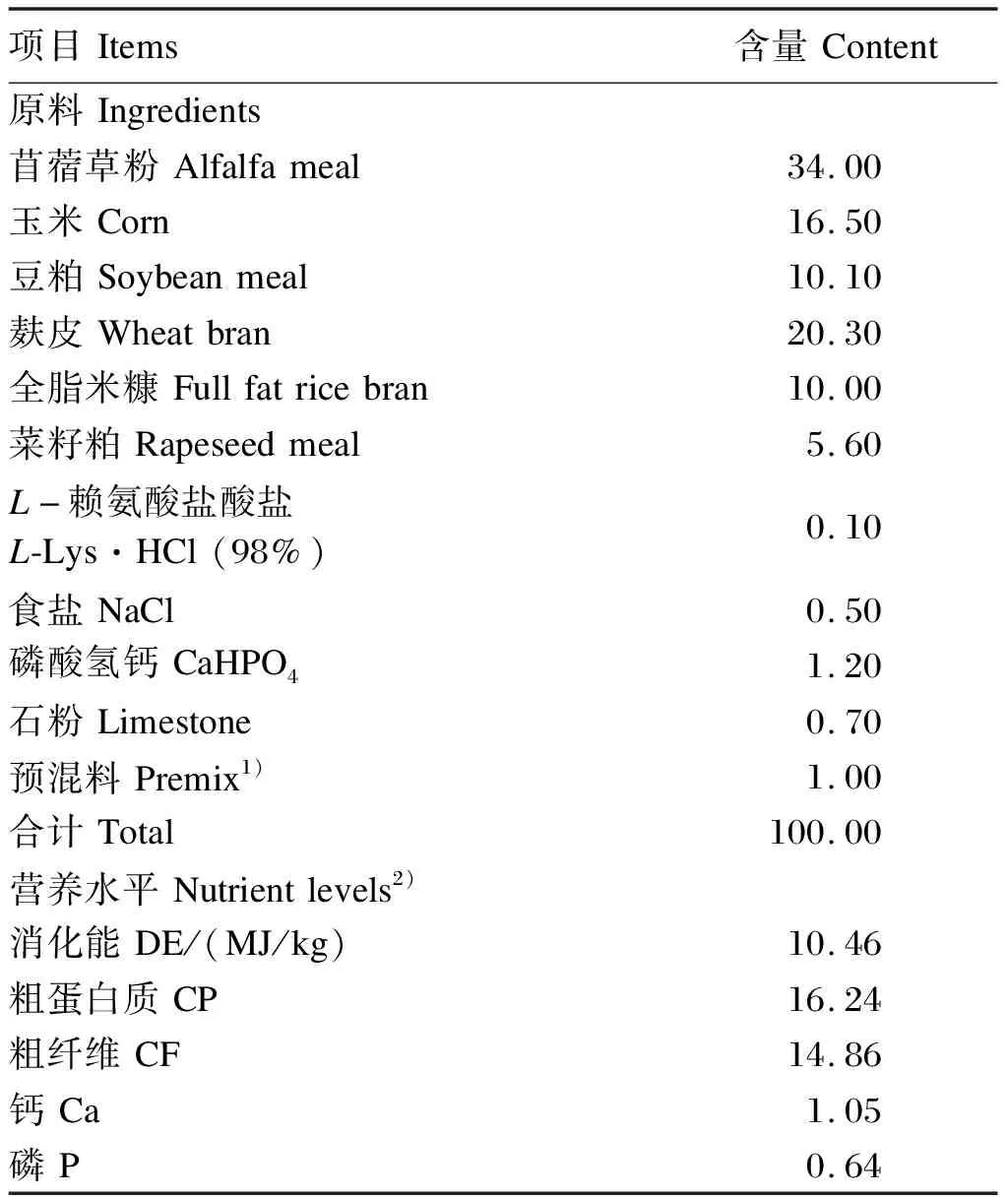
表1 基础饲粮组成及营养水平(风干基础)
1.4 样品采集
在试验正试期结束,于屠宰前采集血液样品。每组选取6只(公母各占1/2)接近平均体重的兔,进行耳缘静脉采血5 mL。采血后静置10 min,待血液凝固后,经3 000 r/min离心20 min,然后分离血清并分装于1.5 mL的EP管中,放置在-80 ℃冰箱中冷冻保存,用于各项血清生化指标的检测。将采完血的肉兔进行屠宰,各取十二指肠、空肠和回肠中段3 cm的组织样品,用生理盐水冲洗干净并迅速固定于10%福尔马林缓冲液中24 h。
1.5 测定指标及方法
1.5.1 生长性能指标
于正式试验开始和结束时空腹称重,计算平均日增重;准确记录试验兔的喂料量和余料量,计算平均日采食量;根据试验兔平均日采食量和平均日增重,计算料重比;记录发生过腹泻兔数和死亡兔数,计算腹泻率和死亡率,计算公式如下:
腹泻率(%)=(腹泻兔数/试验兔数)×100;死亡率(%)=(死亡兔数/试验兔数)×100。
1.5.2 肠道发育指标
固定的样品经洗涤、脱水、透明、浸蜡、包埋等处理后,切片(5 μm)置于载玻片上,苏木精-伊红染色,封片。选择完整且走向良好的绒毛来测量绒毛高度和隐窝深度。绒毛高度是从绒毛顶端到绒毛隐窝交界处的距离,隐窝深度定义为相邻绒毛间的内陷深度,用Nikon Eclipse TS100显微镜和NIS-ELEMENT F3.2成像软件对组织进行组织学检查并拍照。使用Imageproplus 5.1图像分析软件分析每个样品的绒毛高度和隐窝深度,每个样品测量10次,获得每个指标的平均值,用于统计分析。
1.5.3 血清生化指标
血清铜锌超氧化物歧化酶、总超氧化物歧化酶、谷胱甘肽过氧化物酶活性及丙二醛、生长激素和胰岛素样生长因子-Ⅰ含量均采用南京建成生物工程研究所试剂盒测定,根据说明书要求进行操作。
1.6 数据分析
试验数据先由Excel 2010进行汇总,再利用SPSS 19.0软件进行方差和卡方分析,差异显著时采用Duncan氏法进行多重比较,数据结果用平均值±标准差表示,P<0.05为差异显著。腹泻率和死亡率的显著性采用卡方检验。
2 结 果
2.1 乳酸锌对断奶肉兔生长性能的影响
由表2可知,与对照组相比,饲粮中添加40和80 mg/kg乳酸锌对断奶肉兔试验末重有显著影响(P<0.05);乳酸锌各添加水平均可以显著提高断奶肉兔平均日增重(P<0.05),显著降低料重比、腹泻率和死亡率(P<0.05)。随着乳酸锌添加水平的增加,断奶肉兔平均日增重更高,料重比更低,但对平均日采食量无显著影响(P>0.05);饲粮添加80 mg/kg乳酸锌的效果表现优于添加同水平硫酸锌,但是除试验末重和腹泻率外,其他生长性能指标差异不显著(P>0.05);40 mg/kg乳酸锌组与硫酸锌组在试验末重、平均日采食量、平均日增重和料重比等指标上差异均不显著(P>0.05),但其腹泻率和死亡率显著低于硫酸锌组(P<0.05)。

表2 饲粮乳酸锌添加水平对断奶肉兔生长性能的影响
2.2 乳酸锌对断奶肉兔肠道发育的影响
由表3可知,与对照组相比,饲粮中添加40和80 mg/kg乳酸锌对断奶肉兔小肠各段绒毛高度、绒毛高度/隐窝深度和十二指肠隐窝深度有显著影响(P<0.05);乳酸锌各添加水平对空肠和回肠隐窝深度均无显著影响(P>0.05)。小肠各段绒毛高度随着乳酸锌添加水平的增加而增加(P<0.05),饲粮添加80 mg/kg乳酸锌效果显著优于添加同水平硫酸锌(P<0.05);饲粮添加40和80 mg/kg乳酸锌可以显著降低十二指肠的隐窝深度,提高十二指肠和空肠绒毛高度/隐窝深度(P<0.05)。添加40 mg/kg乳酸锌与添加80 mg/kg硫酸锌在提高肉兔小肠绒毛高度、隐窝深度及其比值方面差异不显著(P>0.05),改善肠道发育效果接近。

表3 饲粮乳酸锌添加水平对断奶肉兔肠道发育的影响
2.3 乳酸锌对断奶肉兔血清生化指标的影响
由表4可知,乳酸锌各添加水平对断奶肉兔血清铜锌超氧化物歧化酶、总超氧化物歧化酶、谷胱甘肽过氧化物酶活性及生长激素和胰岛素样生长因子-Ⅰ含量等指标有显著影响(P<0.05),且随着乳酸锌添加水平的增加,血清铜锌超氧化物歧化酶活性和生长激素含量有升高的趋势(P>0.05)。40 mg/kg乳酸锌组肉兔血清酶活性和激素含量与硫酸锌组差异不显著(P>0.05);而饲粮添加80 mg/kg乳酸锌在提高肉兔血清铜锌超氧化歧化酶活性和生长激素含量方面显著高于添加同水平硫酸锌(P<0.05)。

表4 饲粮乳酸锌添加水平对断奶肉兔血清生化指标的影响
3 讨 论
3.1 乳酸锌对断奶肉兔生长性能的影响
锌对动物的生长性能有很大影响。现有的研究表明,补锌能促进动物的生长发育。Wang等[6]研究发现,在23日龄断奶仔猪基础饲粮中添加3 000 mg/kg氧化锌能显著提高仔猪的终末体重和平均日增重,降低料重比。Zhang等[7]在28饲龄断奶仔猪基础饲粮中分别添加不同水平氧化锌,结果发现,随着氧化锌添加水平的增加,断奶仔猪的终末体重和平均日增重逐渐增加,料重比逐渐降低。通常来说,动物对锌的需要量受到年龄、生理阶段、饲粮组成和气候环境等多方面影响。目前养兔生产中,肉兔饲粮锌的添加水平为70~140 mg/kg[8]。而在本试验中,饲粮中添加40和80 mg/kg乳酸锌显著提高了断奶肉兔的试验末重、平均日增重,降低了料重比,随着乳酸锌添加水平的增加,平均日增重更高,料重比更低,这与上述研究结果基本一致。现代畜牧生产中,常在饲粮中添加锌来控制幼龄动物的腹泻,提高动物生长性能。Wang等[6]研究发现,断奶仔猪饲粮中添加3 000 mg/kg氧化锌可以显著降低仔猪腹泻率。在本试验中,乳酸锌各添加水平均可以显著降低断奶肉兔腹泻率与死亡率,其中添加40 mg/kg乳酸锌组肉兔的腹泻率和死亡率最低。
已有研究表明,饲粮中添加有机锌对动物生长性能的作用效果要优于无机锌。Cui等[9]研究发现,在雌性水貂饲粮中添加相同水平的甘氨酸锌和硫酸锌,甘氨酸锌组水貂的平均日增重和料重比显著优于硫酸锌组。任战军等[5]研究了蛋氨酸锌和硫酸锌对幼龄艾哥兔生长性能的影响,结果表明蛋氨酸锌对改善艾哥兔生长性能的效果优于硫酸锌。本试验中,80 mg/kg乳酸锌组肉兔的试验末重显著高于同水平硫酸锌组,表明乳酸锌对肉兔生长性能的提高效果要好于相同添加水平的硫酸锌,这与上述研究结果一致。同时,本试验中40 mg/kg乳酸锌组与硫酸锌组在肉兔试验末重、平均日增重和料重比等指标方面差异不显著,表明饲粮添加40 mg/kg乳酸锌的饲喂效果与添加80 mg/kg硫酸锌相当。
3.2 乳酸锌对断奶肉兔肠道发育的影响
锌参与肠道结构和功能的维持,在肠道免疫功能中起着至关重要的作用。缺锌会导致空肠绒毛萎缩和扁平化,这种形态上的变化会减少肠表面积的吸收,并且单位面积的绒毛数量会大大减少,而锌的补充会导致肠黏膜的加速再生和刷状边缘酶水平的增加[10]。动物体内锌的吸收主要部位是小肠,其中十二指肠远端和空肠近端起关键作用。锌吸收通过2种方式进行:主动和被动运输。主动运输是由特定转运蛋白进行的,其有效性随着食物中锌摄入量的增加而增加;被动运输是通过扩散机制进行的,其有效性与肠道腔内锌的浓度呈正比[11]。
小肠绒毛高度和隐窝深度是经常被用作监测肠黏膜形态的可靠指标,在肠道发育和营养吸收中起着关键作用[12]。Zhu等[13]研究发现,饲粮中添加锌显著增加了猪的小肠绒毛高度,降低了小肠隐窝深度。Wang等[6]研究结果表明,饲粮中添加氧化锌(3 000 mg/kg)可显著提高断奶仔猪的回肠绒毛高度、宽度和表面积。Hu等[14]研究发现,饲粮中添加2 000 mg/kg氧化锌可显著增加断奶仔猪空肠黏膜的绒毛高度和绒毛高度/隐窝深度。Shao等[15]在鸡上研究也表明,玉米豆粕饲粮中添加锌能显著提高鸡小肠的绒毛高度。在本试验中,乳酸锌添加水平对肉兔小肠各段绒毛高度、绒毛高度/隐窝深度和十二指肠隐窝深度有显著影响,但对空肠和回肠隐窝深度无显著影响,这可能是由于与上述研究相比,本试验中锌的添加水平较低,不足以引起空肠和回肠隐窝深度指标的变化。40 mg/kg乳酸锌组与硫酸锌组在提高小肠绒毛高度、隐窝深度及其两者比值方面差异不显著,这表明饲粮添加低水平乳酸锌对于改善肠道发育的效果与高水平硫酸锌相当。
3.3 乳酸锌对断奶肉兔血清生化指标的影响
锌作为动物体内不可缺少的微量元素,参与减少由于细胞中自由基的形成而引起的氧化损伤,在抗氧化防御系统中起着重要的作用,影响着多种抗氧化酶的活性[16-17]。Ma等[18]研究发现,饲粮中添加90 mg/kg锌能显著提高肉鸡肝脏中超氧化物歧化酶和谷胱甘肽过氧化物酶的活性,且甘氨酸锌的添加效果比硫酸锌更好。张彩英等[19]研究发现,饲粮中添加高剂量氧化锌(3 000 mg/kg)能显著提高断奶仔猪血清铜锌超氧化物歧化酶活性。丙二醛是动物体内脂质过氧化的最终产物,锌可以防止脂质过氧化,从而减少动物体内丙二醛含量[20]。Nagalakshmi等[21]研究发现,补锌显著降低了大鼠血清丙二醛含量。在本试验中,乳酸锌和硫酸锌均可以显著提高断奶肉兔血清抗氧化酶活性,随着乳酸锌添加水平的增加有提高血清抗氧化酶活性的趋势,40 mg/kg乳酸锌组与硫酸锌组在血清铜锌超氧化物歧化酶、总超氧化物歧化酶、谷胱甘肽过氧化物歧化酶活性和丙二醛含量上差异不显著,表明低水平乳酸锌与高水平硫酸锌的抗氧化作用相同。
研究发现,锌不仅影响动物体内多种酶的活性,而且还与许多激素的合成和分泌有关。锌是胰岛素样生长因子-Ⅰ诱导细胞增殖、激活胰岛素样生长因子-Ⅰ受体酪氨酸激酶和肝脏胰岛素样生长因子-Ⅰ基因表达的必要条件[22]。因此,锌缺乏会严重影响胰岛素样生长因子-Ⅰ的合成、分泌和功能。Macdonald[23]研究表明,在缺锌的情况下,动物体内的胰岛素样生长因子-Ⅰ水平持续降低。Cesur等[24]研究发现,缺锌会降低动物血液中的胰岛素样生长因子-Ⅰ含量,但在口服补锌后会显著恢复。Ninh等[25]对锌与动物体内胰岛素样生长因子-Ⅰ含量之间的关系研究表明,生长期严重缺锌动物的血液胰岛素样生长因子-Ⅰ含量显著降低。生长激素在动物体内起着多种生理作用,其分泌依赖于锌离子。朱宇旌等[26]研究发现,在断奶仔猪饲粮中添加半胱胺螯合锌能显著提高仔猪血清生长激素含量。在本试验中,乳酸锌和硫酸锌均可以显著提高断奶肉兔血清生长激素和胰岛素样生长因子-Ⅰ含量,且随着乳酸锌添加水平的增加,血清生长激素和胰岛素样生长因子-Ⅰ含量有显著升高的趋势,这与上述研究结果一致。饲粮添加40 mg/kg乳酸锌效果与添加80 mg/kg硫酸锌差异不显著,提示饲粮中添加低水平的乳酸锌可代替高水平的硫酸锌。
4 结 论
综合分析表明,乳酸锌的添加效果显著优于相同水平的硫酸锌;添加40 mg/kg乳酸锌在肉兔促生长、降低腹泻死亡率、改善肠道发育、提高血清酶活性和激素含量等方面与80 mg/kg硫酸锌效果相当,从生态环保、资源有效利用、食品安全和国家法律法规的角度考虑,在养兔生产中建议使用40 mg/kg乳酸锌代替80 mg/kg硫酸锌。
猜你喜欢
作文小学中年级(2022年5期)2022-06-02
今日农业(2021年12期)2021-10-14
中国民间疗法(2021年9期)2021-07-22
河南畜牧兽医(2020年1期)2020-01-11
河南畜牧兽医(2020年13期)2020-01-10
中成药(2018年10期)2018-10-26
国际妇产科学杂志(2016年2期)2016-06-16
中国资源综合利用(2016年7期)2016-02-03
河北医学(2015年8期)2015-06-23
中国实用医药(2014年7期)2014-04-26
