
饲粮氮硫比对藏羔羊瘤胃发育及微生物菌群结构和功能的影响
2021-03-03 03:30闫小兰李鹏翔侯生珍贾建磊
动物营养学报 2021年2期
闫小兰 李鹏翔 侯生珍 魏 涛 贾建磊*
(1.青海大学农牧学院,西宁 810016;2.青海省畜牧兽医科学院,西宁 810016;3.山东省烟台市蓬莱市畜牧兽医站,烟台 265600)
青海省有着全国45%的藏羊资源,藏羊肉作为优质的肉品具有蛋白质含量高、脂肪含量低、矿物质丰富、口感鲜嫩、膻味少等优点。随着我国居民生活水平的提高,对藏羊肉需求量增加,因此,探索提高藏羊生产性能的方法尤为重要。蛋氨酸是牛、羊等动物的限制性氨基酸,在瘤胃微生物的作用下的净降解高达60%[1]。硫作为蛋氨酸的合成元素,同时也是自然界中较丰富的矿物质元素之一,与动物体内碳水物的化合合成、脂肪代谢及蛋白质代谢等活动密切相关[2]。动物缺硫则出现食欲减退、毛品质下降、体质虚弱、甚至死亡;但硫过量也会降低动物的采食量、饲粮转化率和生产性能,并干扰其他矿物元素的吸收利用[3-5]。不同动物在不同的生长时期补充一定的硫能够提高动物的生产性能[6-7]。当硫不过量饲喂时,有机硫和无机硫的消化相似,有机硫还原成硫化物的可能性较小,更容易直接被瘤胃微生物利用转化为微生物蛋白质[8]。研究表明,反刍动物硫需要量通常以氮硫比的形式表示。适宜的氮硫比可提高瘤胃纤维物质的消化以及纤维消化酶的活性,促进瘤胃微生物蛋白质的合成,促进氮的沉积,从而促进反刍动物的生长[9-10]。我国目前藏羊饲养仍以粗秸秆类主,其中含硫氨基酸缺乏、非蛋白氮比例大,因此,研究藏羊饲粮中适宜的氮硫比对促进氮代谢以及提高饲粮利用率和藏羊生产水平具有重要意义。
NRC(1988)推荐反刍动物氮硫比为10∶1;但根据目前研究发现该值无法满足不同动物在不同时期的需求[11]。前苏联标准[12]推荐毛肉兼用型绵羊饲粮适宜氮硫比为5∶1~6∶1,肉毛兼用型绵羊为6∶1~7∶1,生长期绵羊为8∶1~9∶1。研究发现,安哥拉山羊产毛期的饲粮适宜氮硫比为7.2∶1.0[13];内蒙古白绒山羊在绒生长旺盛期的饲粮适宜氮硫比为7.11∶1.00,在生长缓慢期的饲粮适宜氮硫比为7.80∶1.00[14]。冯媛[10]通过体外模拟发现,饲粮氮硫比为7∶1~8∶1的绵羊的营养物质利用率最佳。
瘤胃是反刍动物特有的消化器官,其内定居着大量的微生物,能将固体饲料消化分解生成挥发性脂肪酸(VFA)、甲烷和微生物蛋白质,进而被胃肠道所吸收。适宜氮硫比能改善瘤胃的发酵环境,进而影响瘤胃上皮的发育和瘤胃微生物的活性。各国的饲养标准对反刍动物饲粮的适宜氮硫比的规定存在差异。已有的研究结果也不尽一致,而且多集中在产毛动物上,并且多以无机硫为添加源,尚未进行有机硫为添加源方面的研究。因此,本研究以青海省当地的特色物种——藏羊为研究对象,以生物学效率为100%的蛋氨酸为硫添加源,探究不同氮硫比对藏羔羊瘤胃组织形态、瘤胃液VFA含量以及微生物菌群结构和功能的影响,以期为藏羊的高效养殖提供数据支持。
1 材料与方法
1.1 试验设计与试验饲粮
本试验采用单因素随机试验设计,选择健康状况良好、体重相近的60日龄断奶藏羔羊150只,随机分5组,每组30只。由于尿素不适用于在幼龄反刍动物饲粮中应用,因此本试验以蛋氨酸和赖氨酸替代精料补充料中的麸皮,以此调节各组饲粮的氮硫比分别为10.5∶1.0(A组)、9.0∶1.0(B组)、7.5∶1.0(C组)、6.0∶1.0(D组)、4.5∶1.0(E组)。据青海省当地饲料原料营养成分[10]配制能量和蛋白质水平相似的5组饲粮,试验饲粮组成及营养水平见表1。

表1 试验饲粮组成及营养水平(干物质基础)
1.2 饲养管理与样品采集
各组分别以氮硫比不同的饲粮进行限量饲喂。试验动物采用全舍饲的饲养方式,按组分栏饲养,分别在每天08:00、16:00进行饲喂,自由饮水。60日龄开始补饲(干物质采食量为羔羊体重的3%,羔羊随机采食),预试期5 d,正试期120 d。羔羊饲养至180日龄后,每组随机选择6只羔羊早上空腹、颈动脉放血屠宰后,解剖,采集瘤胃腹囊部位瘤胃组织用生理盐水清洗后,放入4%的多聚甲醛中固定。另外采集瘤胃液50 mL,于4层灭菌纱布过滤后将滤液装于5 mL冻存管中,放入液氮中带回实验室于-80 ℃冷冻保存,用于后期瘤胃微生物多样性的测定。
1.3 指标测定
1.3.1 饲粮中营养成分的测定
采用Foss半自动凯氏定氮仪(Kjeltec 2100)测定粗蛋白质(CP)含量,具体方法参考GB/T 6435—2006;粗脂肪(EE)含量使用SER148脂肪测定仪(索氏提取仪)进行测定,具体方法参照GB/T 6433—2006;酸性洗涤纤维(ADF)、中性洗涤纤维(NDF)含量使用ANKOM A200i型半自动纤维分析仪测定,具体方法参考袁翠林等[16];粗灰分(Ash)含量采用茂福炉高温灼烧法测定;钙(Ca)、磷(P)、硫(S)含量的测定分别参考GB/T 6436—2002、GB/T 6437—2002、GB/T 17776—1999的方法测定。
1.3.2 瘤胃组织形态的测定
瘤胃组织在固定液中固定3 d,然后在不同浓度的葡萄糖下进行梯度脱水,包埋剂冷冻包埋,切5~7 μm连续切片,进行苏木精-伊红(HE)染色,中性树脂封片,显微镜下观察瘤胃组织形态。使用Image-Pro Plus 5.1图像分析系统测量瘤胃乳头长度、乳头宽度、乳头角化层厚度、黏膜层厚度、黏膜下层厚度及肌层厚度。每个样本进行3张非连续性切片的观察,每张切片选取3个视野,每个视野中测定5组试验数据。
瘤胃乳头密度的测定:使用解剖显微镜对1 cm2的新鲜瘤胃组织样品进行直接计数,每只羊选取3个组织样品计数,取平均值。
1.3.3 瘤胃液VFA含量测定
利用气相色谱仪(Thermo Scientific,美国)测定瘤胃液VFA含量。具体参考Liang等[17]方法。
1.3.4 微生物多样性的测定
将采集的瘤胃液送到北京百迈客科技有限公司利用Illumina Hiseq 2500高通量测序平台进行测序。主要操作流程:提取瘤胃液样品中的总DNA,经1%琼脂糖凝胶电泳检测DNA质量合格后,根据保守区设计得到引物,在引物末端加上测序接头,进行PCR扩增并对其产物进行纯化、定量和均一形成测序文库,建好的文库先进行文库质检,质检合格的文库用Illumina Hiseq 2500进行测序。
1.4 生物信息分析
将测序后得到的原始测序序列经过PE reads拼接、Tags过滤、去除嵌合体等最终得到的有效序列,使用QIIME软件对Tags在97%的相似度水平下进行聚类划分操作分类单元(OTU)。将OTU的代表序列与微生物参考数据库进行比对可得到每个OTU对应的物种分类信息,进而在各水平统计各样品群落组成,利用QIIME软件生成不同分类水平上的物种分类表,再利用R语言工具绘制成样品各分类水平下的群落结构图。使用Mothur软件对样品α多样性指数进行评估;采用Binary-Jaccard距离算法在OTU水平上比较各样品间微生物类群的组成差异;使用PICRUSt软件通过比对16SRNA测序数据获得的物种组成信息,推测样本中的功能基因组成,从而分析不同分组之间在功能上的差异。
1.5 数据分析
通过Excel 2013对数据进行初步整理,应用SAS 9.0软件进行单因素方差分析(one-way ANOVA),试验数据以平均值和均值标准误(SEM)表示,P<0.05表示差异显著。
2 结 果
2.1 饲粮氮硫比对藏羔羊瘤胃组织形态的影响
由表2可知,B组瘤胃乳头长度、黏膜下层厚度以及肌层厚度显著高于其他4组(P<0.05),C、D、E组瘤胃乳头长度显著高于A组(P<0.05);B、C、D组瘤胃乳头宽度显著高于A、E组(P<0.05);B、E组瘤胃乳头密度显著高于其他3组(P<0.05);C组瘤胃角化层厚度显著高于其他4组(P<0.05),其他4组间差异不显著(P>0.05)。
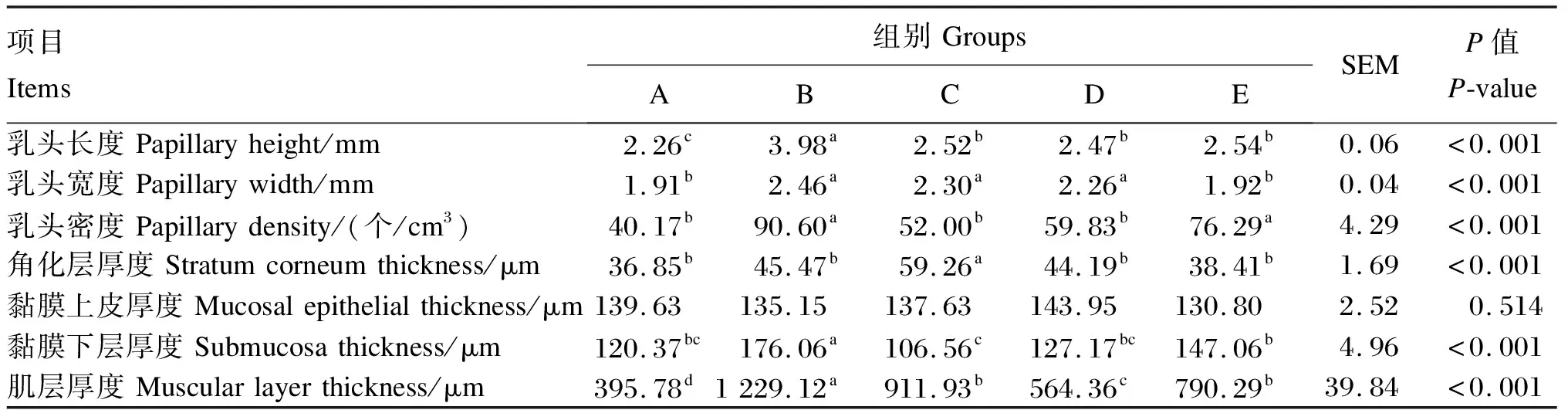
表2 饲粮氮硫比对藏羔羊瘤胃组织形态的影响
2.2 饲粮氮硫比对藏羔羊瘤胃液VFA含量的影响
由表3可知,B组瘤胃液总挥发性脂肪酸(TVFA)、乙酸、丙酸、丁酸、异丁酸含量显著高于其他4组(P<0.05)。
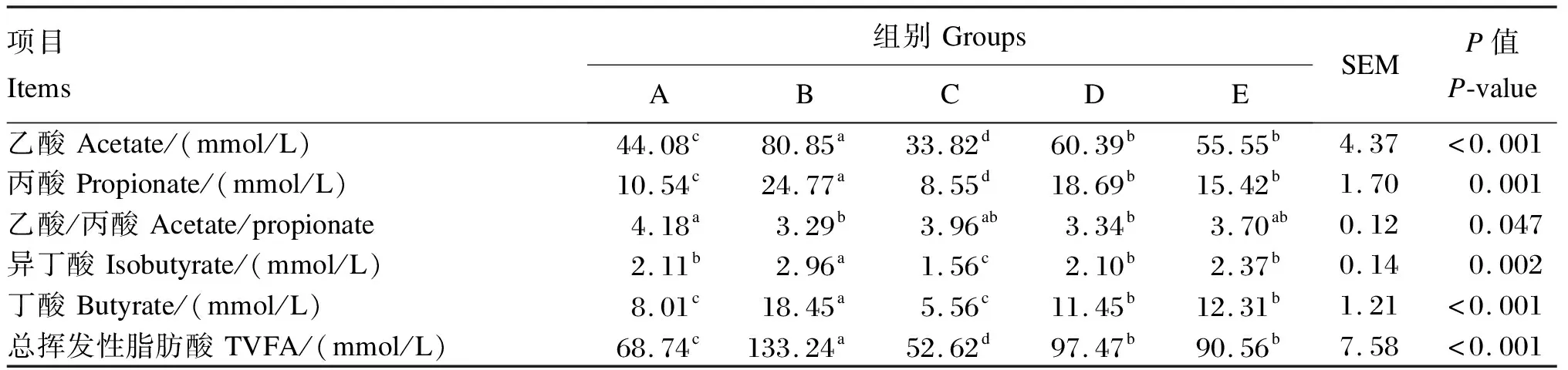
表3 饲粮不同氮硫比对藏羔羊瘤胃液VFA含量的影响
2.3 饲粮氮硫比对藏羔羊瘤胃微生物多样性的影响
对30只藏羔羊瘤胃液样品进行高通量测序,将得到的原始序列双端Read拼接、过滤后共产生3 433 220条优质序列,基于相似度大于97%的原则,每个样品至少产生87 770条优质序列,将测序获得的有效序列进行聚类,共获得1 022个OTU,其中B组OTU数量显著高于其他组(P<0.05)。由图1可知,5组间共有632个OTU,A、B、C、D和E组的独有OTU数量分别为13、23、22、8、8个。

图1 藏羔羊瘤胃微生物Venn图
应用Binary-Jaccard算法在OTU水平上对所有样品进行主成分分析(PCoA),由图1可知,PC1轴的可信度为34.20%,PC2轴的可信度为10.06%,饲喂同一饲粮的藏羔羊瘤胃样点相对聚集,饲喂不同饲粮的藏羔羊瘤胃样点相对分散。
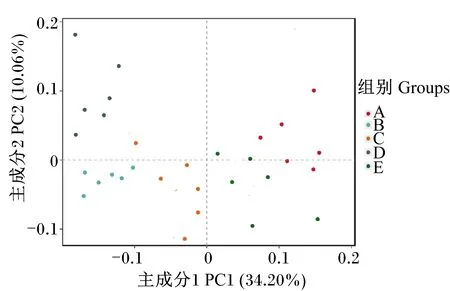
图2 主成分分析图
α多样性指数可以反映瘤胃微生物的丰富度和均匀度。在同一测序深度下,比较5组的α多样性指数,由表4可知,B、C组Shannon指数显著高于A组(P<0.05),其余各组间差异不显著(P>0.05);A组Simpson指数显著高于其他4组(P<0.05);B组Chao1指数和ACE指数显著高于其他4组(P<0.05);基因文库覆盖率均大于99%,各组之间差异不显著(P>0.05)。
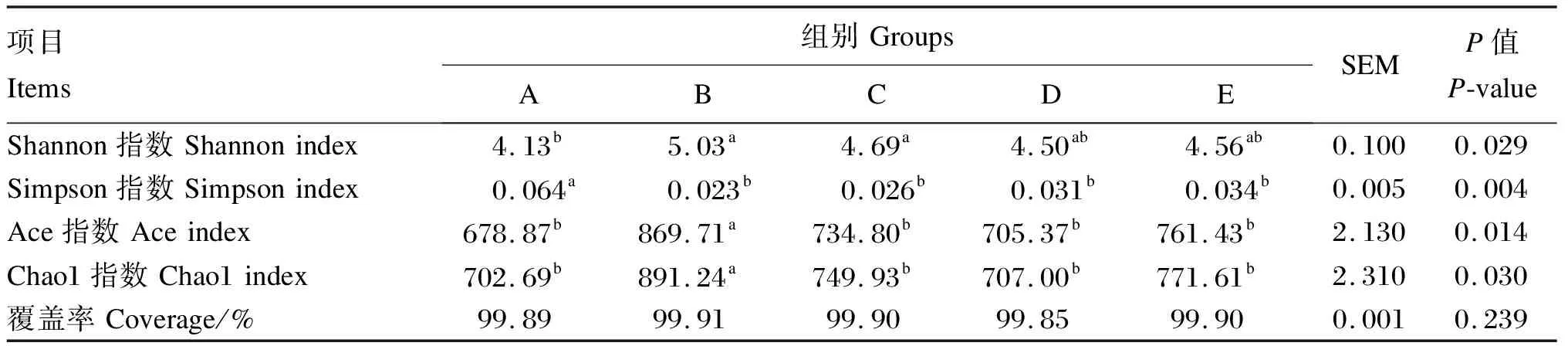
表4 α多样性指数
2.4 饲粮氮硫比对藏羔羊瘤胃菌群结构的影响
各样本进行I11uminaHiSeq测序平台双末端测序,所得有效序列在不同分类水平上进行物种注释和统计共得到19个门、94个科、228个属。由图3和表5可知,在门水平下相对丰度大于0.1%的前10个菌门中厚壁菌门(Firmicutes)为5组的优势菌门,A组厚壁菌门相对丰度显著高于C组(P<0.05)。

表5 饲粮氮硫比饲粮对藏羔羊瘤胃细菌门水平相对丰度的影响

Others:其他;Cyanobacteria:蓝藻菌门;Chloroflexi:绿弯菌门;Proteobacteria:变形菌门;Actinobacteria:放线菌门;Synergistetes:互养菌门;Bacteroidetes:拟杆菌门;Firmicutes厚壁菌门。
由图4和表6可知,在科水平下,5组中的优势菌科均为毛螺菌科(Lachnospiraceae)、克里斯滕森氏菌科(Christensenellaceae)、韦荣氏球菌科(Veillonellaceae)、瘤胃球菌科(Ruminococcaceae);D组毛螺菌科相对丰度显著高于A、B、E组(P<0.05);A组韦荣氏球菌科相对丰度显著高于C、D组(P<0.05),其他各组间无显著差异(P>0.05);各组克里斯滕森氏菌科(Christensenellaceae)和瘤胃球菌科(Ruminococcaceae)相对丰度差异不显著(P>0.05)。
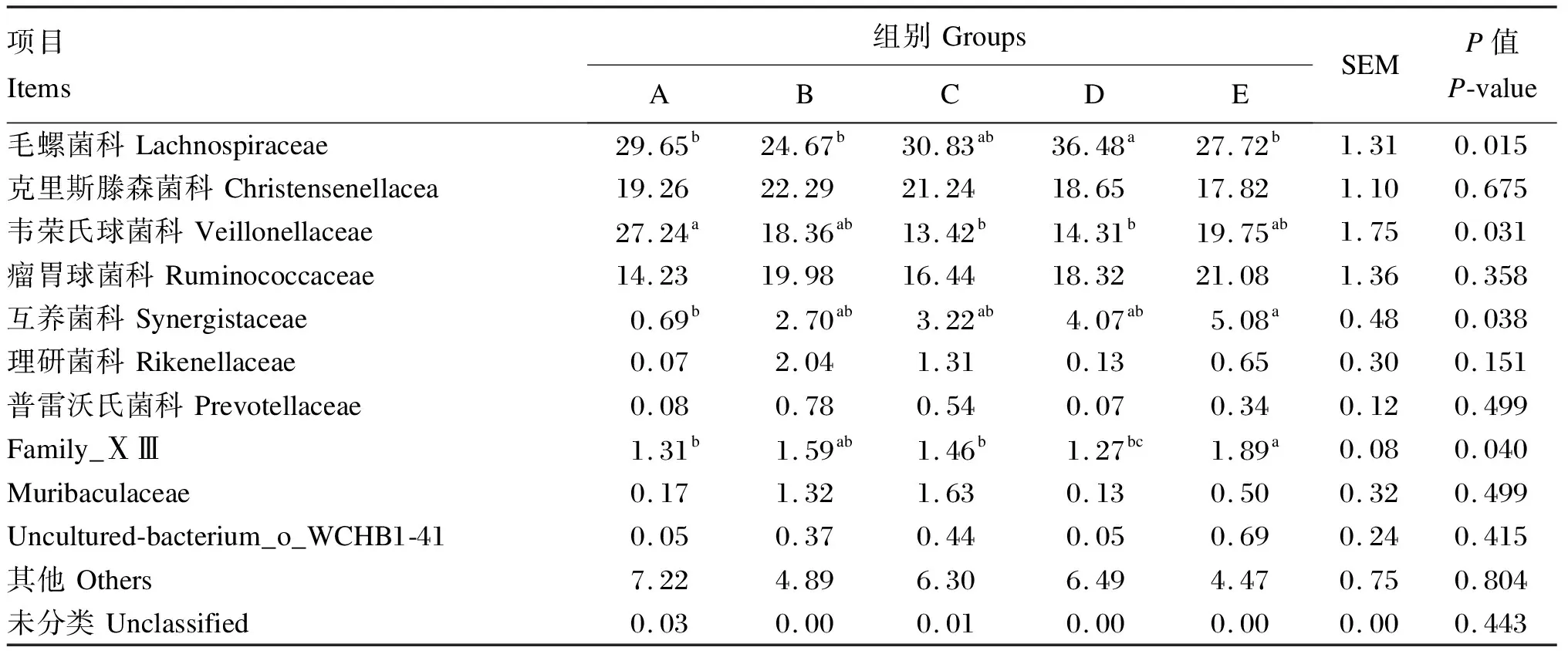
表6 饲粮氮硫比饲粮对藏羔羊瘤胃细菌科水平相对丰度的影响
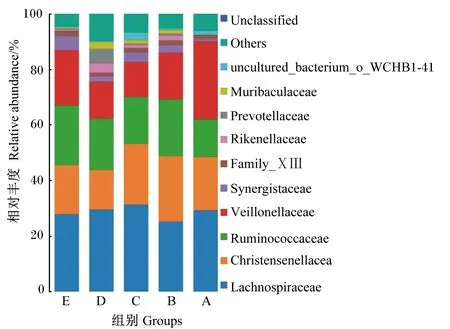
Unclassified:未分类;Others:其他;Prevotellaceae:普雷沃氏菌科;Rikenellaceae:理研菌科;Synergistaceae:互养菌科;Ruminococcaceae:瘤胃球菌科;Veillonellaceae:韦荣氏球菌科;Christensenellacea:克里斯滕森菌科;Lachnospiraceae:毛螺菌科。
2.5 功能预测分析
利用PICRUSt软件对已测得的基因数据进行预测,将得到的功能基因中的信息参考KEGG数据库2级功能层基因类别划分归类,5组藏羔羊瘤胃细菌在二级功能层共包含39种基因功能家族,其中碳水化合物代谢、氨基酸代谢、能量代谢、辅酶和维生素代谢以及核苷酸代谢的功能基因相对丰度在5组中较高。
3 讨 论
反刍动物瘤胃组织形态结构的变化可以反映其消化功能,瘤胃上皮组织与瘤胃内容物直接接触,其组织形态可依据不同的发酵水平及外部环境变化发生相应形态学变化以满足机体正常生理代谢需要。瘤胃乳头增加了瘤胃上皮与内容物的接触面积,有利于瘤胃上皮对养分的吸收和离子转运[18]。瘤胃乳头长度、宽度和瘤胃壁的厚度是评价瘤胃发育的重要指标[19]。刺激瘤胃乳头发育的主要原因是有机酸的存在,尤其是VFA,其刺激强度顺序为丁酸>丙酸>乙酸,这一顺序与VFA被瘤胃壁组织的代谢程度相一致[20]。反刍动物对饲料中碳水化合物的消化吸收是以在瘤胃内生成VFA为主,VFA是反刍动物主要的能量来源,主要由乙酸、丙酸和丁酸组成,能促进反刍动物肠胃运动,抑制肠胃中病原微生物的生长[21]。饲粮中的纤维在纤维分解菌的作用下生成乙酸,淀粉分解菌能够促进丙酸的沉积,厚壁菌门是瘤胃内重要的纤维消化菌[22]。张桂国等[23]研究发现,饲粮氮硫比7.2∶1.0可提高丙酸在TVFA中的比例,降低乙酸/丙酸。殷云浩等[24]研究发现,饲粮氮硫比为7.5∶1.0和9.0∶1.0可显著降低氨态氮及乙酸、丙酸的比例。本试验结果显示,饲粮氮硫比为9.0∶1.0组的瘤胃内的乙酸、丙酸、丁酸以及TVFA含量显著高于其他各组,且乙酸/丙酸最低。乙酸/丙酸常作为评价瘤胃发酵类型的标志,一般认为乙酸/丙酸小于3属于丙酸发酵类型,而大于3为乙酸发酵类型[25]。本试验瘤胃的乙酸/丙酸均在3以上,说明不同氮硫比没有改变瘤胃发酵类型。饲粮氮硫比为9.0∶1.0时瘤胃乳头长度、宽度、密度及肌层厚度较高,这与瘤胃液VFA含量的变化相吻合。由于研究对象与前人不同,因此藏羊饲粮适宜氮硫比与前人研究结果不一致。
反刍动物瘤胃内聚集着大量依靠发酵来消化饲料的微生物,瘤胃微生物的组成与结构是影响瘤胃发酵功能的重要因素。诸多要素影响瘤胃微生物区系的结构,饲粮中的各元素也是其中的重要一项。硫作为动物体内常量矿物质元素之一,是瘤胃微生物细胞结构成分或作为酶、辅酶的成分,能够影响瘤胃微生物的活性。矿物元素缺乏会降低瘤胃微生物的生长效率,降低饲料消化率,使微生物群体数量减少,最终也会降低动物对饲料的采食量[26]。分析瘤胃液样品的OTU数量和α多样性,其结果可以反映微生物群落的丰度和多样性。本研究通过HiSeq测序发现氮硫比为9.0∶1.0组的瘤胃OTU数量及ACE、Chao1、Shannon指数高于其他各组,表明该氮硫比下藏羊瘤胃微生物种类较丰富。另外,本试验中各组覆盖率均高于99%,覆盖率高于97%说明测序样品取样充分[27],因此,本次测序结果能真实地反映藏羔羊瘤胃细菌群落种类和结构。

Amino acid metabolism:氨基酸代谢;Biosynthesis of other secondary metabolites;其他次生代谢产物的生物合成;Cancers: Overview:癌症:概述;Cancers: Specifictypes:癌症:特定类型;Carbohydrate metabolism:碳水化合物代谢;Cardiovascular diseases:心血管疾病;Cell growth and death:细胞生长与死亡;Cell motility:细胞运动;Cellular community:细胞群落;Circulatory system:循环系统;Development:发育成长;Digestive system:消化系统;Drug resistance:耐药性;Endocrine and metabolic diseases:内分泌代谢疾病;Endocrine system:内分泌系统;Energy metabolism:能量代谢;Environmental adaptation:环境适应;Excretory system:排泄系统;Folding, sorting and degradation:折叠、分类和降解;Global and overview maps:全球和概览地图;Glycan biosynthesis and metabolism:聚糖生物合成与代谢;Immune diseases:免疫性疾病;Immune system:免疫系统;Infectious diseases: Bacterial:传染病:细菌性;Infectious diseases: Parasitic:传染病:寄生虫病;Infectious diseases: Viral:传染病:病毒性;Lipid metabolism:脂质代谢;Membrane transport:膜传输;Metabolism of cofactors and vitamins:辅酶和维生素代谢;Metabolism of other amino acids:其他氨基酸代谢;Metabolism of terpenoids and polyketides:萜类化合物和聚酮类化合物的代谢;Nervous system:神经系统;Neurodegenerative diseases:神经变性疾病;Nucleotide metabolism:核苷酸代谢;Replication and repair:复制和修复;Sensory system:感觉系统;Signal transduction:信号转导;Signaling molecules and interaction:信号分子与相互作用;Substance dependence物质依赖性:Transcription转录;Translation:翻译;Transport and catabolism:运输和分解代谢;Xenobiotics biodegradation and metabolism:外源生物降解与代谢。
微生物主要可分为细菌、原虫和真菌三大类[28]。其中细菌是瘤胃中多样化最高的群体,能够利用纤维、淀粉、蛋白质和糖,在动物健康和生产性能中具有重要作用。研究发现,厚壁菌门和拟杆菌门是反刍动物瘤胃内的优势菌门[29-20]。本研究通过高通量测序技术对饲喂不同氮硫比饲粮的藏羔羊瘤胃液进行菌群结构分析,发现各组的优势菌门均为厚壁菌门,其他菌门的相对丰度较少。曾燕等[31]报道,绵羊瘤胃内相对丰度最高的为厚壁菌门(44.37%),蒙古羊瘤胃内相对丰度最高的为拟杆菌门(67.00%)。韩旭峰[32]研究发现,80~110日龄陕北绒山羊瘤胃微生物以厚壁菌门为主。以上结果表明,瘤胃细菌菌体结构受反刍动物品种、日龄等因素的影响,其群落结构发生变化[33]。厚壁菌门中包含的瘤胃球菌属、假丁酸弧菌属、丁酸弧菌属、颤螺旋菌属以及真杆菌属等菌属主要参与纤维物质的分解[34]。本研究中,各组厚壁菌门的相对丰度均高达84%以上,氮硫比为10.5∶1.0时厚壁菌门的丰度最高(93.96%),显著高于氮硫比为6.0∶1.0时,而在非纤维物质的降解中发挥重要作用的拟杆菌门相对丰度最低(0.38%),不利于营养物质的消化。研究表明,有机硫制剂能选择性的刺激瘤胃纤维分解菌和真菌[35]。因此,氮硫比为10.5∶1.0时比较适于厚壁菌门的繁殖,不利于拟杆菌门的生长。本试验进一步在科水平上的研究发现,厚壁菌门下的毛螺菌科、克里斯滕森氏菌科、韦荣氏球菌科、瘤胃球菌科为各组的优势菌科。毛螺菌在消化纤维过程中与产琥珀酸丝状杆菌具有协同作用,并且对病原菌具有拮抗作用[36-37]。韦荣氏球菌科与氨基酸、辅酶、能量和维生素的代谢,糖苷键代谢以次级代谢产物呈正相关[38]。本试验中,氮硫比为6.0∶1.0时毛螺菌科相对丰度显著高于氮硫比为10.5∶1.0、9.0∶1.0、4.5∶1.0时,氮硫水平为10.5∶1.0时韦荣氏球菌科相对丰度显著高于氮硫比为7.5∶1.0和6.0∶1.0时。研究发现,细菌种类对底物的偏好不同,由于毛螺菌科不易体外分离培养,对毛螺菌的生理特性理解不够深入[39]。因此,这一变化机制有待进一步研究。
瘤胃微生物菌群发酵饲料为反刍动物提供蛋白质和能量,是一个庞大的生物资源库,因此积极挖掘一些与营养生理功能密切相关的瘤胃菌群功能基因是非常重要的,如碳水化合物转运及代谢、氨基酸运输和代谢、VFA的生成等[40]。Abhauer等[41]研究结果证实蛋白质和碳水化合物的代谢是微生物生存必需的一项基本功能。本试验通过利用PICRUSt软件对藏羔羊瘤胃细菌测序数据进行KEGG数据库对比分析后发现,碳水化合物代谢、氨基酸代谢、能量代谢、辅酶和维生素代谢以及核苷酸代谢的功能基因相对丰度在5组中较高。由此可知,藏羔羊瘤胃细菌在提高宿主纤维与蛋白质的利用率至关重要。
4 结 论
① 饲粮氮硫比为9.0∶1.0时瘤胃组织形态发育较好,瘤胃液中VFA含量较高,瘤胃微生物的相对丰度及多样性较高。
② 通过KEGG数据库对比分析发现,羔羊瘤胃细菌基因功能主要集中在碳水化合物代谢和氨基酸代谢。
③ 饲粮氮硫比为9.0∶1.0更能经济有效地促进瘤胃发育和改善羔羊瘤胃微生物区系结构。
猜你喜欢
文萃报·周二版(2022年25期)2022-06-23
科学(2020年2期)2020-08-24
中成药(2018年10期)2018-10-26
纤维复合材料(2018年4期)2018-04-28
现代检验医学杂志(2016年1期)2016-11-12
中国塑料(2016年12期)2016-06-15
湖南畜牧兽医(2016年3期)2016-06-05
大型铸锻件(2015年4期)2016-01-12
大型铸锻件(2015年1期)2016-01-12
动物营养学报(2015年10期)2015-12-01
