
多年生稻稻瘟病抗性评价
2021-03-03 09:28何奕霏秦世雯张石来黄光福张静杨勤忠胡凤益
中国稻米 2021年1期
何奕霏 秦世雯 张石来 黄光福 张静 杨勤忠 胡凤益*
(1 云南大学农学院/云南省多年生稻工程技术研究中心,昆明650504;2 云南省农业科学院农业环境资源研究所,昆明650205;第一作者:heyifei@mail.ynu.edu.cn;*通讯作者:hfengyi@ynu.edu.cn)
稻瘟病是由稻梨孢菌(Magnaporthe oryzae)引起的水稻真菌病害之一,具有发病急、破坏力强、发病地区广泛、发病时期及发病部位类型多、造成经济损失严重等病害特征[1]。利用抗病基因改良品种是目前防治该病害最有效的措施,至今已定位了100 多个稻瘟病抗性主效基因,并克隆了30 多个稻瘟病抗性主效基因[2]。利用已开发的功能分子标记,可准确地检测不同水稻品种中存在的稻瘟病抗性基因[3]。
利用长雄野生稻地下茎无性繁殖特性培育的多年生稻已在生产上推广应用[4-6]。多年生稻不仅能实现免耕、减缓耕作带来的水土流失等问题,同时还能减少人力和农资投入[7]。在多年生稻试验示范和推广过程中发现,其对稻瘟病具有一定抗性,但具体的抗性水平和抗病基因还未明晰。虽然已发现长雄野生稻第12 号染色体上具有一定稻瘟病广谱抗性基因Pi57[8],但该基因是否导入多年生稻品种(系)并引起相应抗性反应还未知。因此,本研究对5 个多年生稻品种(系)PR23、PR24、PR25、PR101、PR107,及其父本长雄野生稻、母本RD23 及(RD23/O. longistaminata)F1进行田间病情调查、接种抗性鉴定,以及分析已知的多个R 基因在多年生稻品种(系)中有无,初步明确多年生稻对稻瘟病的抗性情况,为多年生稻的稻瘟病抗病育种和生产布局提供参考。
1 材料与方法
1.1 试验材料
以多年生稻品种(系)PR23、PR24、PR25、PR101、PR107、长雄野生稻(多年生稻父本)、泰国优质籼稻RD23(多年生稻母本)、(RD23/O. longistaminata)F1及本地感稻瘟病品种丽江新团黑谷(LTH)为供试品种。
供试稻瘟病菌为强致病力CH091C 菌株,由云南省农业科学院农业环境资源研究所保存和提供。
1.2 田间病情调查
2019 年6 月,在云南省西双版纳州多年生稻示范推广田块进行多年生稻品种(系)的叶瘟田间病情情况调查。采用五点取样法,各品种(系)取样数量为100株,参照国际水稻研究所叶瘟抗性评价分级标准[9]进行病级分级,按照病情指数=∑(各级病株数×该病级值)/(调查总株数×最高级值)×100 进行病情指数计算。病情指数为0 表示高抗(HR),0~5%表示抗(R), 5.1%~15%表示中抗(MR),15.1%~25%为中感(MS),25.1%~50%为感(S),>50%为高感(HS)。水稻苗瘟、叶瘟抗性分级标准见表1。
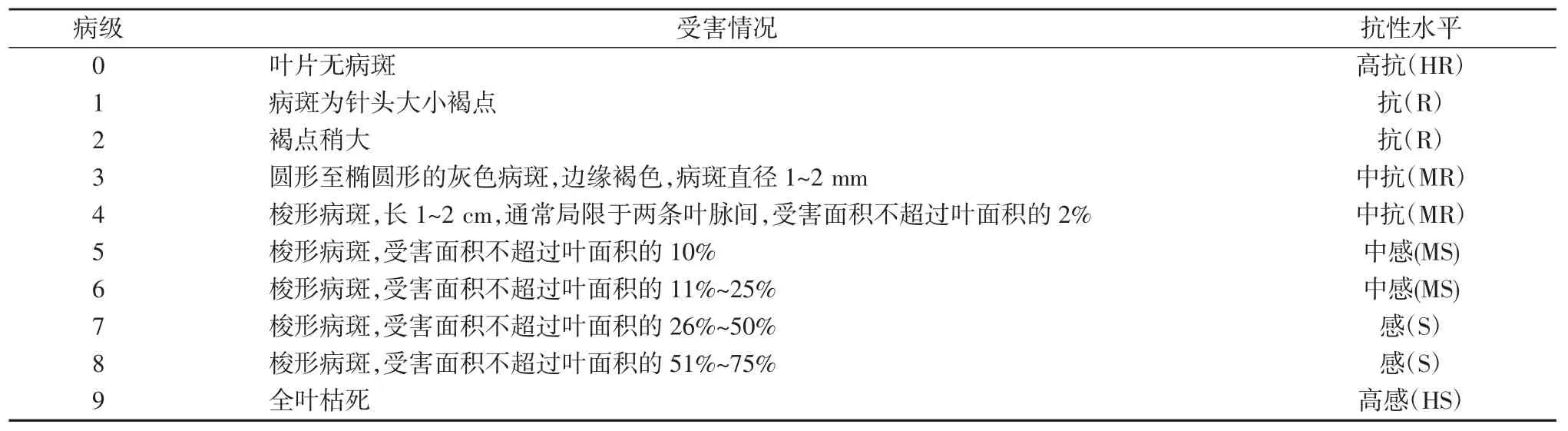
表1 水稻苗瘟叶瘟抗性分级标准(IRRI)

表2 稻瘟病抗性基因检测的引物序列及相关信息
1.3 苗瘟抗性水平评价
以LTH 为感病对照,利用5%次氯酸钠对RD23、PR23、PR24、PR25、PR101、PR107 种子以及长雄野生稻地下茎尖组织进行消毒后催芽,每个材料在播种盘(规格为30 cm×10 cm)中播2 行,每行10 株,共20 株,待长至3 叶1 心时接种稻瘟病菌。稻瘟病菌CH091C菌株于燕麦培养基(燕麦10.0 g、琼脂4.0 g、蔗糖5.0 g,加水至300 mL,120℃高温高压灭菌)上进行活化培养,培养7 d 后将菌丝洗净放置于光照培养箱中,产孢培养2 d 后配置浓度约1×105个/mL 孢子悬浮液。用喷壶均匀喷洒于水稻叶片上,放置于培养箱中25℃保湿培养24 h。接种7 d 后,参照国际水稻研究所苗瘟抗性鉴定分级标准进行抗性水平鉴定。
1.4 稻瘟病抗性基因检测
利用已知的多个稻瘟病抗性基因功能标记和紧密连锁SSR 标记对多年生稻材料进行PCR 扩增(表2),扩增产物利用1.0%(目标产物>500bp)或2.0%的琼脂糖凝胶和8%聚丙烯酰胺凝胶电泳进行检测(目标产物<500 bp)。
2 结果与分析
2.1 稻瘟病田间病情调查结果
通过调查多年生稻示范推广田的稻瘟病发病情况,发现多年生稻品种(系)和父本长雄野生稻无明显叶瘟症状,呈现较强的稻瘟病抗性,PR23、PR24、PR25、PR101、PR107、长 雄 野 生 稻、F1病 情 指 数 分 别 为1.44%、1.67%、1.56%、1.11%、1.33%、0、0.74%(表3)。而母本RD23 具有典型稻瘟病斑,调查病株最轻病级达3级(叶片出现椭圆灰白病斑),大部分病株病级分布于5~7 级(受害面积10%~50%),最高病级可达8 级(叶部受害面积为51%~75%),病情指数为63.33%(表3)。
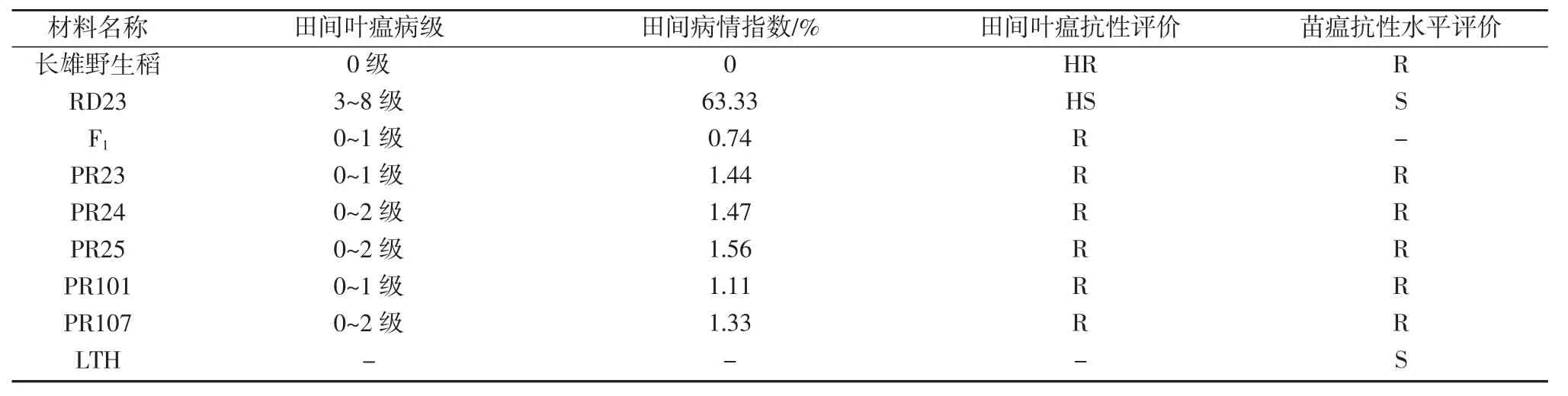
表3 多年生稻品系(种)田间病情调查和苗瘟抗性评价结果
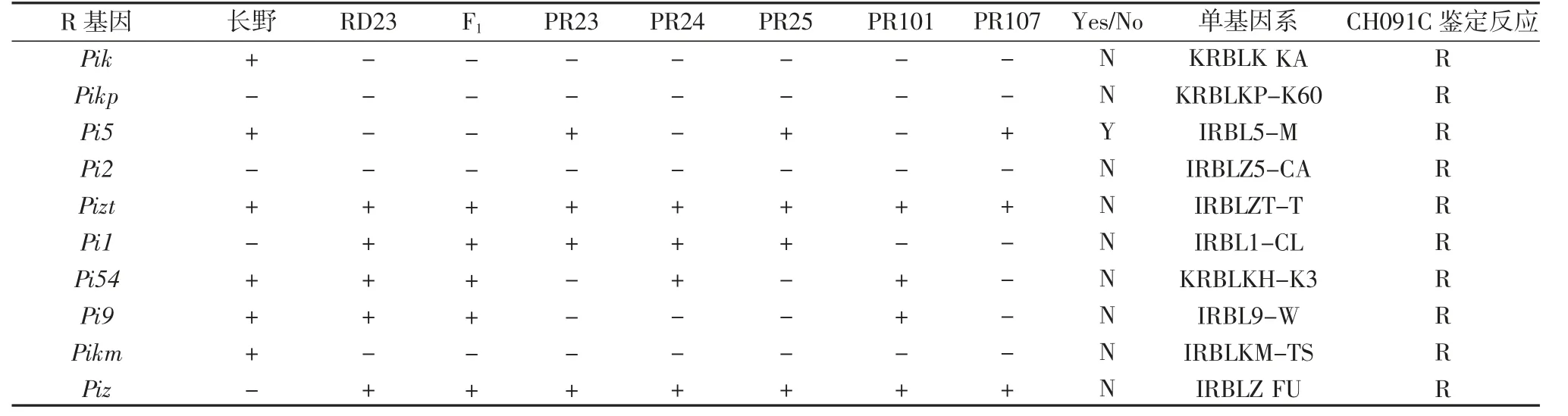
表4 10 个R 基因在多年生稻品系(种)及亲本中的分布检测
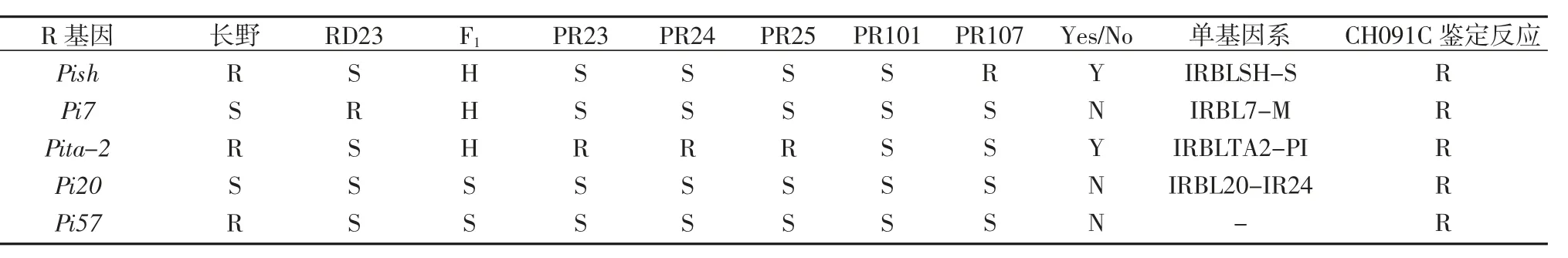
表5 5 个R 基因在多年生稻及其亲本中的多态性检测
2.2 苗瘟抗性水平评价
接种强致病力稻瘟病菌CH091C 菌株7 d 后,PR23、PR24、PR25、PR101、PR107 和长雄野生稻仅有针尖大小病斑,表现为抗病。而感病对照丽江新团黑谷(LTH)和多年生稻品种(系)母本RD23 均具有典型叶瘟症状,表现为感病(表3)。多年生稻品种(系)苗期接种稻瘟病菌进行稻瘟病抗性水平鉴定与田间病情调查结果较为一致,说明多年生稻品系(种)具有较强的抗瘟性,同时该特性可能来源于父本长雄野生稻。
2.3 稻瘟病抗性基因检测
利用功能标记检测10 个已知的R 基因在多年生稻中的分布,在PR23 和PR25 中均检测到抗性基因Pizt、Pi5、Pi1、Piz,但只有Pi5 在长雄野生稻以及PR23、PR25 中检测得到,且在RD23 中缺失,所以PR23、PR25 所携带的Pi5 基因可能来源于长雄野生稻(表4)。
PR107 携带Pizt、Piz、Pi5 稻瘟病抗病基因,仅Pi5在长雄野生稻检测到,而RD23 中缺失,因此来源于长雄野生稻的Pi5 可能为PR107 的稻瘟病抗病基因(表5)。
利用与Pish、Pita-2、Pi7、Pi20、Pi57(t)基因分别连锁的SSR 标记RM212(Pish)、RM6905(Pita-2)、RM273 84/RM27386(Pi7)、OSR32(Pi20)、RM5364/RM7102(Pi 57(t))对5 个多年生稻品系(种)进行基因型评价,发现PR23、PR24、PR25 中携带来自长雄野生稻的Pita-2位点,而PR107 中携带来自长雄野生稻的Pish 位点(表5)。推测PR23、PR24、PR25 的稻瘟病抗性基因可能来源于长雄野生稻的Pita-2 位点,而PR107 的稻瘟病抗性基因可能来源于长雄野生稻的Pish 位点。
因此,PR23 和PR25 的稻瘟病抗性基因可能来源于长雄野生稻的Pi5 基因和Pita-2 位点,PR24 稻瘟病抗性基因可能来源于长雄野生稻的Pita-2 位点,PR107 稻瘟病抗性基因可能来源于长雄野生稻的Pi5基因和Pish 位点,本文涉及到的稻瘟病抗性基因或位点PR101 中均不存在,推测其稻瘟病抗性来自长雄野生稻未知的稻瘟病抗性基因。
3 结论与讨论
稻瘟病是危害最严重的水稻病害之一,培育和推广抗病品种是目前最科学的防治对策。由于稻瘟病菌变异频率较高,大部分抗病品种的抗性会随着应用时间的增长而逐渐降低直至消失[17]。因此,需要不断挖掘、鉴定新的抗病品种进行推广或作为抗源应用于未来的稻瘟病抗性育种中。本文针对具有抗稻瘟病潜力的5 个主栽多年生稻品种(系)的抗性水平进行了初步探究。在田间病情调查中,这5 个多年生稻品种(系)的病情指数均未达到感病系数,抗性鉴定中,这5 个多年生稻品种(系)对接种稻瘟病菌为完全免疫反应。因此,可以认为这些多年生稻抗稻瘟病。此次多年生稻田间抗性的评价是针对其自然状态下的发病情况来评定,反应了多年生稻在田间的自然抗性的差异,今后可以进一步建立病圃对多年生稻的抗病水平进行更精准的评定。除此之外,对多年生稻抗性水平的全面解析还需进一步利用更多的优势生理小种对其抗谱进行测定,并结合田间鉴定。
多年生稻亲本的田间病情调查结果和抗性鉴定均显示,长雄野生稻为抗性品种,RD23 为感病品种,多年生稻的稻瘟病抗性可能来源于父本长雄野生稻。且有前人报道,基于长雄野生稻转录组中发现大量导致栽培稻感稻瘟病的基因在长雄野生稻中没有检测到相应的基因信号,长雄野生稻可能是稻瘟病抗性基因库[18]。本文利用功能性分子标记和紧密连锁SSR 标记对多个稻瘟病抗性基因在多年生稻品种(系)的分布及遗传背景进行分析,发现3 个抗性基因位点Pish、Pita-2、Pi5可能为多年生稻中来源于长雄野生稻的抗性基因。另外,本研究中还发现部分多年生稻的稻瘟病抗性可能来源于长雄野生稻中其余已知基因或未知基因,这些未知基因很可能是长雄野生稻中未被挖掘的稻瘟病抗性新基因,需进行更深入的研究。
无论是田间自然发病、接种鉴定,还是鉴定部分已知的稻瘟病抗性基因或位点,均表明已经育成的多年生稻23、云大24、云大25、云大101、云大107 具有较好的稻瘟病抗性,这对多年生稻的示范推广、品种布局、植保技术等示范推广具有实际指导意义。
猜你喜欢
智慧农业导刊(2022年21期)2022-11-03
当代水产(2022年7期)2022-09-20
今日农业(2022年4期)2022-06-01
作物学报(2022年6期)2022-04-08
中国饲料(2021年17期)2021-11-02
农民致富之友(2020年15期)2020-05-25
农民致富之友(2019年24期)2019-08-20
发明与创新·大科技(2019年5期)2019-07-31
食品工业科技(2014年13期)2014-03-11
现代农业科技(2009年19期)2009-03-20
