
红树林生态修复固碳效果的主要影响因素分析
2021-03-02 06:57陈顺洋安文硕陈光程
应用海洋学学报 2021年1期
陈顺洋,安文硕,陈 彬,陈光程*
(1.自然资源部第三海洋研究所,福建 厦门 361005; 2.福建省海洋生态保护与修复重点实验室,福建 厦门 361005;3.自然资源部海峡西岸海岛海岸带生态系统野外科学观测研究站,福建 厦门 361005)
红树林、海草床和盐沼湿地是重要海洋“蓝碳”生态系统,它们的面积虽不及全球海洋面积的2%,但每年埋藏到沉积物中的有机碳分别约占到全球海岸带和海洋碳埋藏量的46.9%和45.7%[1-2],在减缓全球气候变暖中发挥着重要的碳汇功能。由于红树林在固碳方面的重要作用,其生态保护和修复被认为是应对气候变化中“基于自然的解决方案”的重要内容[3]。自本世纪以来,我国在加强红树林湿地保护的同时也大力开展红树林种植等修复工程,红树林面积较本世纪初有了显著提高[4],在提高湿地生物多样性、消浪促淤和固碳增汇等方面也发挥着重要的作用。
在红树林生态修复中,适宜的生境条件(滩涂高程、水文动力和气候等)是红树林生态修复成功与否的关键因素,决定了修复后红树植物是否成活和定植[5],以及湿地的有机碳累积、底栖动物分布、初级生产等过程,最终决定了红树林生态系统的固碳能力[6-9]。此外,红树植物物种和种植方式等也影响着修复后生态系统结构和功能的发展[9-10]。本研究综述了红树林的生境条件、物种和种植措施等因素对生态系统固碳效果的影响,以期在发挥红树林生态效益的同时提高红树林固碳增汇的生态功能,为开展红树林生态修复提供指导。
1 红树林生态修复后生态系统固碳的特点
红树林位于海陆交接的潮间带,具有高的净初级生产力,可以吸收大气中的二氧化碳并储存在植被和土壤中[11-13]。在潮汐过程中,红树林复杂的植被地上结构(如地表支柱/呼吸根、茂密的植株等)发挥的消浪作用有利于促进潮水中颗粒有机碳的沉降[13-14],植物凋落物和死亡的根系分解后部分也能埋藏到沉积物中[11,15]。随着红树林发育,土壤pH降低、温度降低、红树酚类的输入等变化都有利于红树植物有机碳在土壤中累积[16-18]。同时,红树林中食草性底栖动物对红树林凋落物的储存和摄食作用也促进了红树植物有机碳埋藏进入土壤[11,19-20]。
红树林固碳主要反映在红树林植被和沉积物碳库中的有机碳累积,而凋落物和死木等碳库的累积量相对较低[21-22]。不同物种红树植物的生长速率存在差异[9],乔木或速生种红树植物生长速率较高[23]。一般认为种植20 a左右的红树林植被可达到成熟林的状态[24-25]。Chen等(2021)分析了国内外不同修复时间红树林的植被碳储量(图1),进一步发现人工红树林的植被碳库在种植20 a左右可达到全球红树林植被碳储量的平均水平[22],与植物群落的发育规律一致。
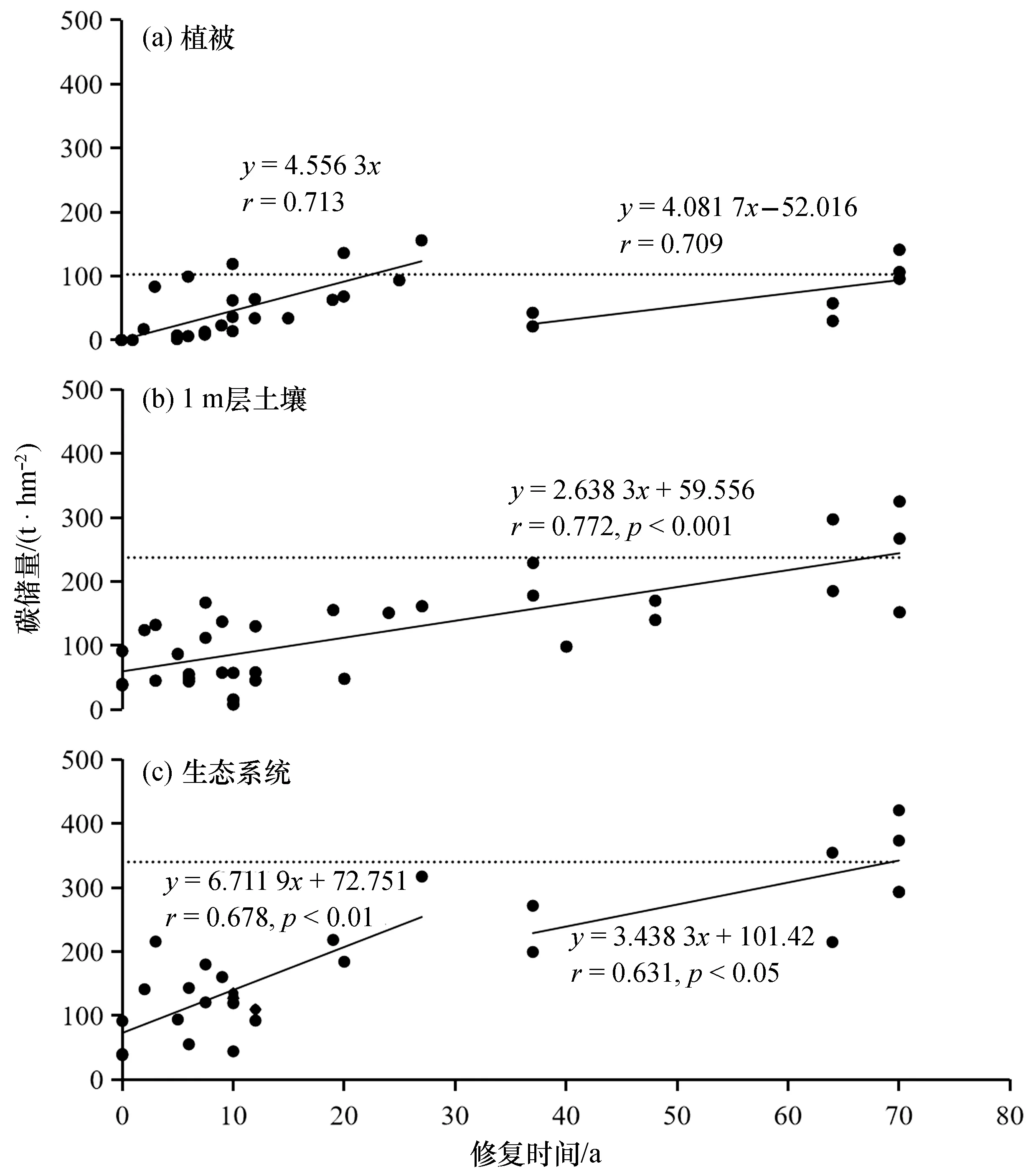
图1 不同修复时间红树林碳储量Fig. 1 Carbon stocks of mangroves in different restoration times图中资料引自文献[22],虚线为全球红树林碳储量平均值。
与植被相比,红树林土壤发育相对缓慢,尤其在种植初期,土壤中的有机碳累积量要明显低于植被[20]。相对于植被而言,红树林土壤中有机碳储量随恢复时间的变化是稳定增加的,以碳计大约为2.64 t/(hm2·a)[22],因此在植被达到成熟状态后土壤碳库对湿地生态系统固碳的贡献会逐渐增加。红树植物凋落物和死亡根系是土壤有机碳的主要来源[11,26-27]。红树林恢复初期土壤中累积的有机碳主要集中在表层,随着恢复时间的推移,植物对土壤有机碳的贡献才影响到底层土壤[6]。土壤中有机碳含量随深度逐渐降低,这种空间变化与植物根系的分布特征类似,因此学者也认为红树植物根系是土壤有机碳的主要来源[27]。
2 生境条件对红树林固碳的影响
根据地形、地貌和风浪等生境特点可将红树林划分为前缘浪击型(包括海岛、沙洲及碳酸盐生境等)、内湾型(包括内湾、潟湖等)和河流型(包括河口、三角洲等)[28-29]。前缘浪击型区域靠近大海,受风浪潮汐影响较大,滩面窄、底质粗,氮、磷等营养输入少,不利于红树植物生长[30-31];内湾型和河流型区域分布于隐蔽的河口港湾及沿河流上溯的河岸淤积浅滩,尤其是河流型区域的大量营养物质和淡水输入,有利于红树植物生长[31-32]。同时,河流型区域大量外源沉积物输入有利于形成较厚的土壤碳库层[33],内湾型区域有利于红树植物有机碳埋藏累积于土壤[13,16]。不同生境类型红树林碳库调查发现,河口内湾区域红树林的碳储量高于开阔海岸和河口边缘区域[13]。因此,在河口、河流或三角洲、内湾、潟湖等区域开展的红树林生态修复有利于红树植物生长和土壤有机碳的累积。
盐度是影响红树植物生长分布的主要环境条件之一[5]。红树植物对盐度有一定的适应范围,他们可分布在低盐的河口地区,也可生长在高盐的海洋环境中[34]。但红树植物的生长受到海水盐度的调控,通常在盐度为2.2~34.5范围的河口海岸生长较好,在淡水和高盐海水中生长不良[34-35],影响红树植物的生物量累积。如拉关木(Lagunculariaracemosa)在盐度为40时的生物量只有盐度为10时的43%[36],盐度为30时秋茄(Kandeliaobovata)的生物量较盐度为5时降低了50%[37]。
通常高盐环境能抑制微生物活性,降低土壤有机碳的分解与转化,有利于红树林土壤有机碳保持稳定[38]。当盐度高于一定条件时,红树林沉积物中腐殖质以絮凝形式存在,不会随水流进入外部系统;而海水盐度下降时,高达三分之二的腐殖质从絮凝形式中溶解释放出来进入水体[39]。高盐环境下硫酸盐还原菌与产甲烷菌竞争限制了土壤中的产甲烷作用和甲烷通量[40-41],减少了土壤中甲烷形式的有机碳损失。
潮水浸淹的程度决定了红树植物能否成活和生长,由潮汐和滩涂高程共同决定,在修复实践中则主要考虑滩涂高程的限制作用[37,42]。一般认为,红树林分布于平均海平面(或稍上)与大潮平均高潮位(或最高潮水位)之间的潮滩面[43-45]。红树植物物种对于滩涂高程具有一定的适应范围(表1),但在不同高程下生长表现不同[30,37,60]。实验结果表明,北部湾全日潮海域不同红树植物抗淹水能力表现为:白骨壤(Avicenniamarina)>桐花树(Aegicerascorniculatum)>秋茄>红海榄(Rhizophorastylosa)>木榄(Bruguieragymnorrhiza)[46]。

表1 中国红树物种的环境适宜性

续表1
淹水时长的增加会降低有机物的分解速率,减少碳的损失[61]。泰国废弃虾塘红树林修复6 a后,虾塘底部区域(每天淹水2 h)土壤碳储量较恢复前增加了45%,而在虾塘围堰等高地土壤的碳储量则减少一半,表明适宜的淹水条件(滩涂高程)让土壤保持湿润条件,有利于红树植物的生长和土壤有机碳的保存[62]。但这种差异也可能与虾塘不同区域水文动力条件有关,虾塘围堰的微地形条件不利于植物碎屑的保留。在红树林中,通常表现为内滩和中滩沉积物有机碳含量高于外滩[63],内滩和中滩较弱的水动力条件促进了潮水中颗粒态有机质的沉降,同时降低了植物碎屑被潮汐带走的机会[64-65],这种作用可能比低的分解速率对土壤固碳的贡献更大。
尽管土壤条件被视为决定红树林分布的重要因子[30],但有关土壤环境和红树植物生长之间仍缺少定量的关系,总体上红树植物以泥质潮滩为宜(表1),但也能生长在砂质、玄武岩铁盘层甚至砾石潮滩上[66]。一般认为,红树林土壤较为贫瘠,氮和磷是限制红树植物生长的主要营养元素[67]。如在外源营养输入较多的河口区域,土壤氮、磷含量较高有利于红树植物生长和生物量的累积[31],而土壤中氮、磷限制会增加红树植物地下生物量的比例,相对有利于红树林土壤有机碳含量的增加[68-69]。在植被发育成熟前,红树植物固碳的能力高于土壤,较高的土壤营养水平整体有利于生态系统的固碳。土壤低 pH会限制微生物的物种多样性,降低红树林土壤中有机碳的大量分解,促进土壤碳累积[16]。红树植物虽然普遍适应酸性土壤,但中性偏酸性的土壤较为有利于红树植物生长[70]。因此,中性偏酸性、泥质、营养丰富的土壤有利于维持和提高红树林生态系统的固碳作用。
3 红树物种对红树林固碳能力的影响
物种被认为是影响人工红树林植被生长和生物量累积的最重要因素[9]。各类型红树植物的生长速率有明显差异,通常情况下,生长速率从高到低为速生乔木、乔木、灌木/丛[23,71-73]。不同红树植物对海水盐度、滩涂高程和底质类型等生境因子都呈现出不同的适应性(表1)。所以在红树生态修复过程中,满足生境适宜的条件下,可优先选择生长速率高的物种以提高植被的固碳作用。相同物种在不同区域也可能有不同的快速生长期,如在浙江引种的秋茄,其植被固碳速率以3 a期最高,5 a和10 a期的固碳速率明显降低[74],而福田的秋茄林则是20 a左右接近生长峰值[24]。所以,有必要对红树植物生长进行长期监测,根据其生长规律加以抚育管理,才能有效的提高红树植物的固碳能力。
红树林沉积物中有机碳含量和储量因红树植物群落类型而异,这种差异是植物物种和滩面高程等因素共同作用的结果[11-12]。而通过比较种植于相同生境条件下不同物种的林地沉积物的碳储量则反映出物种在对沉积物固碳影响上的差异[14]。这种差异一方面可归因于不同物种的植被对消浪和网罗有机颗粒物的能力与其形态密切相关;例如,洛阳江口桐花树(灌木型)样地中沉积物的固碳速率高于同龄的秋茄(乔木型)样地[22],地表凋落物现存量也高于秋茄样地,这可能与桐花树群落密集的地上结构有利于有机碎屑的沉降埋藏和减少凋落物的输出有关。另一方面,灌木型和小乔木型的植物群落通常地下根密度高于稀疏的乔木型群落,也导致它们的沉积物有机碳累积速率较高[27]。
4 种植措施对红树林固碳能力的影响
由于地理隔离造成的红树植物种群的适应性差异,选择优良种苗来源有利于植物形成稳定健康的群落[75]。我国红树植物可分为抗低温广布性、嗜热广布性和嗜热窄布性等3种生态类群[30]。研究发现嗜热广布性红树木榄种源对幼苗保存率、株高、基径和单株生物量的影响显著[76-77],生长受种苗原产地纬度高低的影响不大,主要与母树林的状况有关,来自于天然成片生长良好母树林的种苗具有较宽的遗传物质基础和较强的适应性[76]。抗低温广布性红树的种源研究则有不同的结论,本地的种苗通常是修复的最优种源[78]。浙江等高纬度地区开展的秋茄种植显示种源地纬度越高幼苗成活率越高[79-80],这主要与不同地区秋茄耐寒适应能力的不同有关[79]。可见,嗜热性物种选择修复种源时应考虑母树林的状况,应选择天然成片生长良好的红树林作为种源地;抗低温广布性红树植物选择修复种源时应选择纬度相近的区域同时兼顾母树林现状。
采用不同物种搭配的种植方式已被证实比单一物种种植可产生更高的单位面积生物量固碳效果[81-82],这种方式下植物可高效利用生长空间。例如,我国东寨港秋茄和海桑的混交林初级生产力分别为两种纯林的1.93倍和1.55倍[82]。全球红树林土壤碳库分析发现,红树混交林的单位面积土壤碳储量比红树纯林高20%,土壤碳储量与红树植物物种丰富度之间存在相关性,认为在生态修复中应当注意物种的搭配[83]。但如果种植的物种之间生态位存在重叠,则物种间的竞争会导致对植物生长不利。有研究发现海桑和无瓣海桑的混交林平均初级生产力小于两种纯林,可能是因为海桑和无瓣海桑都是速生高大的建群种,在生存资源空间上竞争激烈[84]。而红树物种之间的化感作用也是需要注意的因素。木榄+秋茄群落的化感物质总体上有利于木榄的天然更新[85],无瓣海桑的直接引入有利于提高海莲和秋茄种群的生物量,但对桐花树和白骨壤种群不利[37]。因此在红树林修复中可采用建群种与非建群种、乔木与灌木、阳生物种与耐荫物种进行搭配。
红树林幼林期植被生物量随种植密度的增加而增高(表2),在生长初期阶段个体间的竞争可能还不足以抵消高密度对植被生物量累积的积极作用[9]。高密度种植时植被地上部分阻尼系数的增大可减缓潮汐的运动,促进有机颗粒物沉降[86]。斯里兰卡一项研究发现,沉积速率随种植密度的增加而增加,种植密度达到6.96 株/m2时样地的沉积速率可达光滩的两倍之多[10]。而随着种植时间的推移,种植密度过小,单位面积植物利用光能效率降低,种植密度过大则植株间形成光能和空间的竞争。在福建平潭种植9 a后,1.00株/m2种植密度的秋茄林生物量大于种植密度为0.67株/m2和2.00株/m2的样地[87]。植物生长存在自然稀疏的过程,即由于密度的抑制作用种群内个体会逐渐死亡,个体数量逐渐减少至稳定状态[88]。高密度种植的红树林也存在明显的自疏过程,并符合Yoda 的-3/2自然稀疏法则[89-90]。因此,适当密植保证植被修复的同时实现较高的固碳能力,后期则可通过其自疏过程或人工疏伐的方式来提高植物群落的固碳能力。
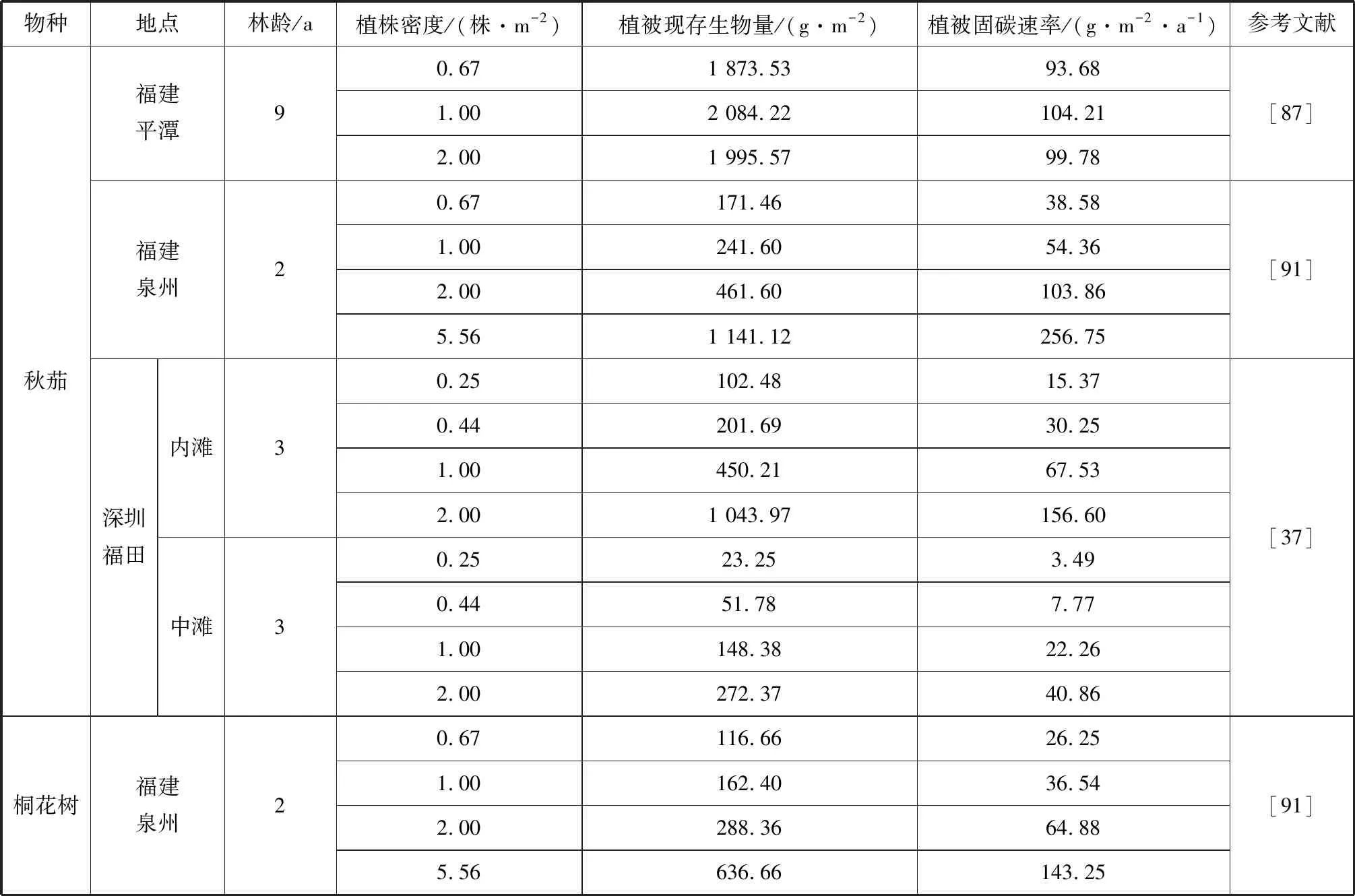
表2 不同种植密度红树植物群落的固碳速率
5 结论及展望
综上所述,红树林生态系统固碳作用主要包括了植被光合固碳和土壤碳沉积(包括红树植物有机碳的埋藏和外源有机碳的输入)。水文动力条件、海水盐度、滩涂高程及底质条件是影响红树林植被成活的关键因子,也影响红树植物的生长、颗粒态有机物的沉降、有机物的输出和保留、有机质的分解等过程,进而影响红树林生态系统固碳能力。因地制宜选择、搭配红树物种及适当密植等措施有利于红树林修复形成稳定的植物群落,提高红树林植被的初级生产力,促进湿地的有机碳累积。这些认识对我国即将开展的大规模红树林生态修复具有十分重要的指导意义。
但对于红树林生态修复的固碳过程和相关机制的认识还不足。如红树林修复后的碳累积/沉积过程、生态系统的固碳潜力、生境条件和种植技术对红树林固碳过程和固碳能力的影响,以及底栖动物群落恢复等其他生态学过程/功能的协同效应等都有待深入研究。需要强调的是,红树林的生态修复是恢复红树林生态系统的结构和功能[91],应在发挥生态修复整体生态效益的同时提高红树林固碳增汇的生态功能,但不能只注重提升湿地固碳功能而忽略其他生态过程和功能的恢复。
猜你喜欢
河北地质(2022年2期)2022-08-22
小猕猴学习画刊(2022年4期)2022-05-05
儿童故事画报·自然探秘(2022年3期)2022-04-27
老年博览·上半月(2021年10期)2021-11-02
海洋通报(2020年5期)2021-01-14
疯狂英语·初中天地(2018年6期)2018-11-24
歌海(2018年4期)2018-05-14
现代园艺(2017年23期)2018-01-18
天然产物研究与开发(2016年6期)2016-06-05
应用海洋学学报(2015年2期)2015-11-22
