
水保措施对柑橘果园土壤细菌群落结构的影响
2021-03-02 03:05:04吴波波肖胜生余小芳丁明军
环境科学研究 2021年2期
吴波波, 王 鹏*, 肖胜生, 余小芳, 舒 旺, 张 华, 丁明军
1.江西师范大学地理与环境学院, 江西 南昌 330022 2.江西师范大学, 鄱阳湖湿地与流域研究教育部重点实验室, 江西 南昌 330022 3.江西省水土保持科学研究院, 江西 南昌 330029
土壤细菌丰度高、种类多、代谢活性丰富,能迅速对环境扰动做出反应,是土壤生态系统重要的活性部分[1],其群落在调节养分循环、影响植物生产力和生态系统稳定性方面起着重要作用[2]. 已有研究证实,影响土壤条件的农业实践会改变土壤微生物群落,尤其是细菌群落[3]. 不同学者在许多研究中也已探索了水保措施对土壤细菌群落结构的影响,大多数研究表明,与轮作和净耕相比,作物覆盖增加了土壤有机质含量和细菌多样性,是改善土壤微环境的有效策略[4-6];轮作增加了土壤细菌的数量,但并没有影响细菌的丰富度或多样性[7];净耕容易导致土壤有机质含量锐减、细菌多样性降低、树体早衰、果实品质下降[8]. 也有研究得出相反结论,例如,Sanaullah等[9]研究了农业管理系统对陆地生态系统功能的影响,发现豆科植物轮作降低了病虫害压力,增加了土壤中的氮含量,从而导致土壤微生物活性增强,细菌多样性提高;DONG等[10]认为,净耕可以增加土壤碳储量和养分含量,稳定土壤结构,从而增强土壤微生物活性. 可见,不同水保措施对土壤细菌群落结构的影响较为复杂,它们之间的相互作用及其作用机理尚未明确,水保措施对土壤特性及细菌群落影响的争论仍然存在. 此外,尽管已有学者开展了水保措施对土壤菌群群落结构的影响研究[6],但对亚热带环境下不同水保措施导致的柑橘果园土壤细菌群落结构响应尚不明确,特别是细菌群落交互作用的网络对亚热带柑橘果园土壤环境变化的响应还鲜有报道.
柑橘是中国乃至世界上最具经济价值的果树之一[11]. 随着人口快速增长及农户追求经济利益最大化等因素的影响,中国南方越来越多的坡地次生林转变为果园[12],柑橘果园也因其高经济收益在该地区广泛分布[13]. 规模机械化开挖对果园土壤的强烈干扰易导致土壤养分循环失衡及农作物生产力下降[14],探索新的水保措施已成为该区域柑橘产业可持续发展的重要途径. 在土壤中,细菌数量、群落结构质量变化(如多样性指数)是土壤健康重要且敏感的指标[15],对土壤细菌群落结构的深入认知,将有助于制定合理有效的水保措施. 此外,土壤细菌群落的分布模式和驱动因素可能在不同水保措施间有所不同,而不同水保措施下的土壤细菌群落在土壤养分利用效率之间的差异会严重影响植物的生长,因此,了解土壤细菌的多样性、群落组成及其交互作用对于通过调控细菌群落以实现作物的可持续生产至关重要.
鉴于此,为研究不同水保措施对柑橘果园土壤细菌群落结构的影响,该研究选取了江西省德安县柑橘果园的4种水土保持试验小区作为研究对象,通过Shannon-Wiener指数和Chao1指数探究了不同水保措施下柑橘果园土壤细菌的多样性,利用高通量测序技术分析了不同水保措施下柑橘果园土壤细菌的群落组成,并通过构建其分子生态网络,探索了不同水保措施对柑橘果园土壤细菌群落交互作用的影响,以期为柑橘果园土壤生态系统研究提供理论基础.
1 材料与方法
1.1 样地概况
试验地位于江西省德安县燕沟小流域内的江西水土保持生态科技园(115°42′38″E~115°43′06″E、29°16′37″N~29°17′40″N). 该区属于亚热带季风气候区,平均海拔30~90 m,多年平均降水量为 1 537 mm,多年平均气温为16.7 ℃,年日照时数为 1 700~2 100 h. 地貌类型主要为浅丘岗地,植被类型属于亚热带常绿阔叶林,植被类型主要有针叶林、山地针叶林、常绿阔叶林等,土壤主要为第四纪红黏土发育的红壤,土壤厚度为0.5~1.5 m. 该区位于中国红壤分布的中心区域,属全国土壤侵蚀二级类型区,在江西省和南方红壤丘陵区具有典型代表性.
1.2 样地设置及采样方法
试验地为柑橘果园水土保持试验区,柑橘果园按照 1 335株hm2的初始密度于2001年栽植. 试验设置4个水保措施(从2001年持续至2019年),分别为狗牙根条带覆盖(TD)、狗牙根全园覆盖(QY)、经济作物萝卜-大豆轮作条带覆盖(LZ)和净耕果园(JG),每个水保措施设置1个小区,共4个试验小区. 试验小区坡度均为10°,面积均为20 m2(宽2 m、长10 m),各小区其他管理措施一致.
2019年5月进行采样. 在每个试验小区内进行3次重复采样,每次采样为5个采样点的混合,按照S型选取5个采样点. 每个采样点去除表面凋落物后,用土钻按照表层(0~10 cm)、中层(10~20 cm)和底层(20~40 cm)进行土壤分层取样,去除石头、植物残留物、动物和其他杂物后分成2份,冷藏带回实验室. 其中,一份放在-80 ℃冰箱保存,用于高通量测序,另一份风干后用于测定土壤基本理化指标. 因测序成本较高,用于高通量测序的样品是3次重复采样样品充分混合而成的.
1.3 试验分析
全磷(TP)含量采用HJ 632—2011《碱熔-钼锑抗分光光度法》测定;对于全氮(TN)含量,将土壤经过浓硫酸-高氯酸消解后,采用HJ 634—2012《氯化钾溶液提取-分光光度法》测定;对于pH,样品加水后(水土比为1∶2.5)采用便携式水质分析仪(HI9828,HANNA,Italy)测定;土壤有机碳(SOC)含量采用碳氮元素分析仪(Elemantar vario MAX,Germany)测定;土壤有机质(SOM)含量采用重铬酸钾-硫酸外加热法测定(总共36个样品,即每个土层设置3个重复).
细菌16S rRNA基因测序采用E.Z.N.A.® Soil DNA Kit (Omega Bio-tek, Norcross, GA, U.S.)提取土壤样品基因组DNA,采用1%琼脂糖凝胶电泳检测抽提的DNA. 对16S rRNA基因的V3~V4高变区片段进行PCR扩增,引物序列为338F(5′-ACTCCTA CGGGAGGCAGCA-3′)和806R(5′-GGACTACHVGGG TWTCTAAT-3′). 扩增条件:95 ℃预变性2 min,接着进行25个循环,包括95 ℃变性30 s,55 ℃退火30 s,72 ℃延伸30 s;循环结束后72 ℃最终延伸5 min(PCR仪,ABI GeneAmp® 9700型,USA). 总共12个样品(每个土层不设置重复),将同一样本的PCR产物混合后用2%琼脂糖凝胶电泳检测,使用AxyPrepDNA凝胶回收试剂盒(AXYGEN,USA)切胶回收PCR产物,Tris_HCl洗脱;2%琼脂糖电泳检测. 参照电泳初步定量结果,将PCR产物用QuantiFluorTM-ST蓝色荧光定量系统(Promega,USA)进行检测定量. 16SrRNA基因测序在上海美吉生物医药科技有限公司的Illumina Miseq PE300平台进行. 原始数据上传至NCBI数据库中(序列号为SUB7239364).
细菌的绝对丰度采用Real-time荧光定量 PCR法获得,在博日LineGene9600plus平台上进行. 反应体系(20 μL):ChamQSYBR ColorqPCRMasterMix(2X)10 μL,5 μmolL正、反向引物各 0.5 μL,样品模板 DNA 2 μL,ddH2O 4.5 μL. 细菌荧光定量PCR的扩增引物为338F(5′-ACTCCTACGGGAGGCAGCA-3′)806R(5′-GGACTACHVGGGTWTCTAAT-3′). 反应条件:95 ℃预变性5 min,95 ℃变性30 s,56 ℃退火30 s,72 ℃延伸40 s,72 ℃修复延伸,共40个循环. 最后按照博日LineGene9600plus实时荧光定量检测系统默认程序制备熔解曲线并检测是否存在非特异性扩增. 每个样品均设置3个平行,并利用不含DNA的去离子水作为阴性对照. 利用含有细菌16S rRNA基因全长的质粒构建标准曲线,稀释梯度为原浓度的10-3~10-8. 所有 PCR反应的扩增效率均在 90%以上,相关系数(R2)均大于99%.
1.4 数据处理
利用Mothur(V.1.36.1)对原始DNA序列进行过滤处理,去除嵌合体,得到优化序列;按照97%相似性将优化序列划分可操作分类单元(Operational Taxonomic Units, OTU);基于OTU进行稀释性曲线分析,计算Chao1指数和Shannon-Wiener指数. 采用RDP classifier贝叶斯算法对照Silva数据库以70%置信度对OTU代表序列进行物种分类,并在各分类水平上统计每个样品的群落组成(为避免各样品微生物量的差异,按最小样本序列数进行样本序列抽平,得到标准化数据用于后续统计分析). 采用单因素方差分析进行不同土层和不同水保措施下土壤样品间细菌16S rRNA的基因拷贝数和土壤理化指标的显著性检验(显著性水平P≤0.05视为差异显著);多重比较分析采用LSD法;相关性分析采用Spearman相关分析法. Veen图用于反映不同水保措施下共有和独有OTU的数量. 基于Bray-Curtis距离做PLS-DA分析以反映细菌群落结构的整体差异,通过冗余分析来研究土壤理化指标与细菌群落的关系.

注: 同一列标有不同小写字母的表示不同处理间差异显著(P<0.05);标有不同大写字母的表示同一处理不同土层之间差异显著(P<0.05).图1 不同水保措施下土壤理化性质Fig.1 Soil physicochemical properties under different soil and water conservation measures
为确定细菌群落成员之间的联系(正相关和负相关),使用Cytoscape 3.4.0进行网络可视化和模块化分析[16]. 网络分析采用CONET插件(http:psbweb05.psb.ugent.beconet)[17],每个OTU的分类标识在门类级别分配,将获得的原始OTU数据(具有分类学丰度的表格)用作输入矩阵,根据CONET网站(http:psbweb05.psb.ugent.beconettutorial4.php)上提供的指南进行网络构建. 参数设置如下:每个OTU至少进行30个序列的预处理和过滤,4个相似性度量(Spearman,Pearson,Kullbackleibler和Bray-Curtis),自动设置阈值并将错误发现率(FDR)校正设置为0.05,导出邻接矩阵,用邻接矩阵来编码每对节点之间的连接关系,对每一条边进行100次的Permutation和 Bootstrap显著性检验.
2 结果与分析
2.1 土壤理化性质和细菌16S rRNA基因拷贝数
在各土层中,TD、QY和LZ土壤中的TN、SOC和SOM含量均高于JG,特别是QY,其3层土壤中的平均TN、SOC和SOM含量与JG相比分别提高了近50%、40.33%和40.26%. 与JG相比,QY表层土壤和LZ中层土壤细菌16S rRNA基因拷贝数分别提高了近1.51倍和0.64倍(见图1). TD土壤细菌16S rRNA基因拷贝数沿剖面深度无显著变化. 值得注意的是:4种水保措施下土壤中的TN、SOC和SOM含量均沿剖面深度呈下降趋势,而QY表层土壤中的TN、SOC和SOM含量在4种水保措施下均显著最高,且这3个指标均与细菌16S rRNA基因拷贝数呈显著正相关(见表1). 此外,与TD、QY和LZ相比,JG仅底层土壤中的TP含量与土壤细菌16S rRNA基因拷贝数较大,且二者呈显著正相关(见表1).
2.2 土壤细菌群落多样性
将柑橘果园不同水保措施下12个土壤样品中的质控后序列按97%相似性进行聚类,得到的细菌优化序列总数为 623 653,有效序列范围为 42 776~61 081. 各样品文库的覆盖率(Coverage)均近99%,说明此次研究样品的基因序列检出率较高,测序结果能够真实反映柑橘果园不同水保措施土壤细菌群落的真实情况.

表1 不同水保措施及不同土层土壤细菌16S rRNA丰度和土壤理化性质之间的相关性

图2 不同水保措施下特有和共有OTU分布的Venn图Fig.2 Venn diagram showing the unique and shared operational taxonomic units (OTU) of bacteria detected in soil samples under different soil and water conservation measures
与JG相比,QY和TD对柑橘果园土壤细菌均匀性、多样性和丰富度均产生了更积极的影响. 图2显示了4种水保措施下土壤样品中独有的和共有的OTU数量. 与JG相比,TD、QY和LZ土壤中独有的细菌OTU数量分别为920、973和812个. 尽管QY对底层土壤细菌多样性没有明显影响,但与其他水保措施相比,QY表层土壤细菌Shannon-Wiener指数和中层土壤细菌Chao1指数均分别为最高,而TD中底层土壤细菌Shannon-Wiener指数和表层土壤细菌Chao1指数分别为最高(见表2). 此外,与TD、QY和LZ相比,JG各层土壤细菌Shannon-Wiener指数均为最低(见表2). 相关性分析表明,TN含量、pH、SOC含量、SOM含量均与Shannon-Wiener指数呈显著正相关,而与Chao1指数无显著相关性(见图3). 在不同水保措施下,土壤细菌群落结构可通过基于Bray-Curtis距离的PLS-DA分析明显区分,并聚成4个类群(见图4).

表2 不同水土保持措施下土壤细菌群落多样性指数(Shannon-Wiener)和物种丰富度指数(Chao1)

注:TD-1、TD-2、TD-3分别表示TD措施下表层、中层、底层土壤,其他以此类推.图4 不同水保措施下土壤细菌群落结构差异(PLS-DA分析)Fig.4 Difference of bacterial community structures under different soil and water conservation measures (Partial Least Squares Discriminant Analysis)
2.3 土壤细菌群落组成
高通量测序得到的3 531条OTUs分属于34门、93纲、230目、368科、616属和1 241种. 图5为门水平上的细菌分类,结果显示,4种水保措施的优势菌门包括绿弯菌(Chloroflexi,占比为28.39%~45.32%)、酸杆菌(Acidobacteria,占比为13.20%~23.31%)、变形菌(Proteobacteria,占比为14.65%~22.95%)和放线菌(Actinobacteria,占比为7.20%~11.26%),占细菌序列的80%以上. 在不同水保措施和土层之间,单个细菌类群的相对丰度差异明显. 与TD、QY和LZ相比,JG土壤中Chloroflexi的相对丰度较高,而Acidobacteria和Proteobacteria的相对丰度较低. 在表层土壤中,QY与JG土壤中Proteobacteria以及QY与LZ土壤中Acidobacteria的相对丰度均较高. 除Chloroflexi相对丰度增加外,不同水保措施下土壤中Proteobacteria、Acidobacteria(TD除外)和Actinobacteria的相对丰度均沿剖面深度呈减少趋势.
图6为环境因子对主要门分类种群(平均相对丰度大于1%)的冗余分析结果. 由图6可见,第一主轴解释了门分类群落59.30%的方差变化,第一主轴和第二主轴共解释了61.69%的方差变化. 其中,TN含量对门分类种群影响的显著性最高(P=0.002),其次为SOC和SOM含量(P=0.005). pH和TP含量的影响均不显著(P分别为0.086和0.445). 所以TN、SOC和SOM含量(尤其是TN含量)是影响不同水保措施下柑橘果园土壤细菌群落组成的主要环境因子. TN、SOC、SOM含量均与Chloroflexi的相对丰度呈显著负相关,与Proteobacteria、Acidobacteria、Actinobacteria的相对丰度均呈显著正相关.
2.4 土壤细菌群落交互作用

注: 平均相对丰度低于1%以下的部分合并,并用others表示.图5 不同水保措施下土壤细菌群落门水平分类Fig.5 Phylum-level taxonomic composition of the bacterial community under different soil and water conservation measures
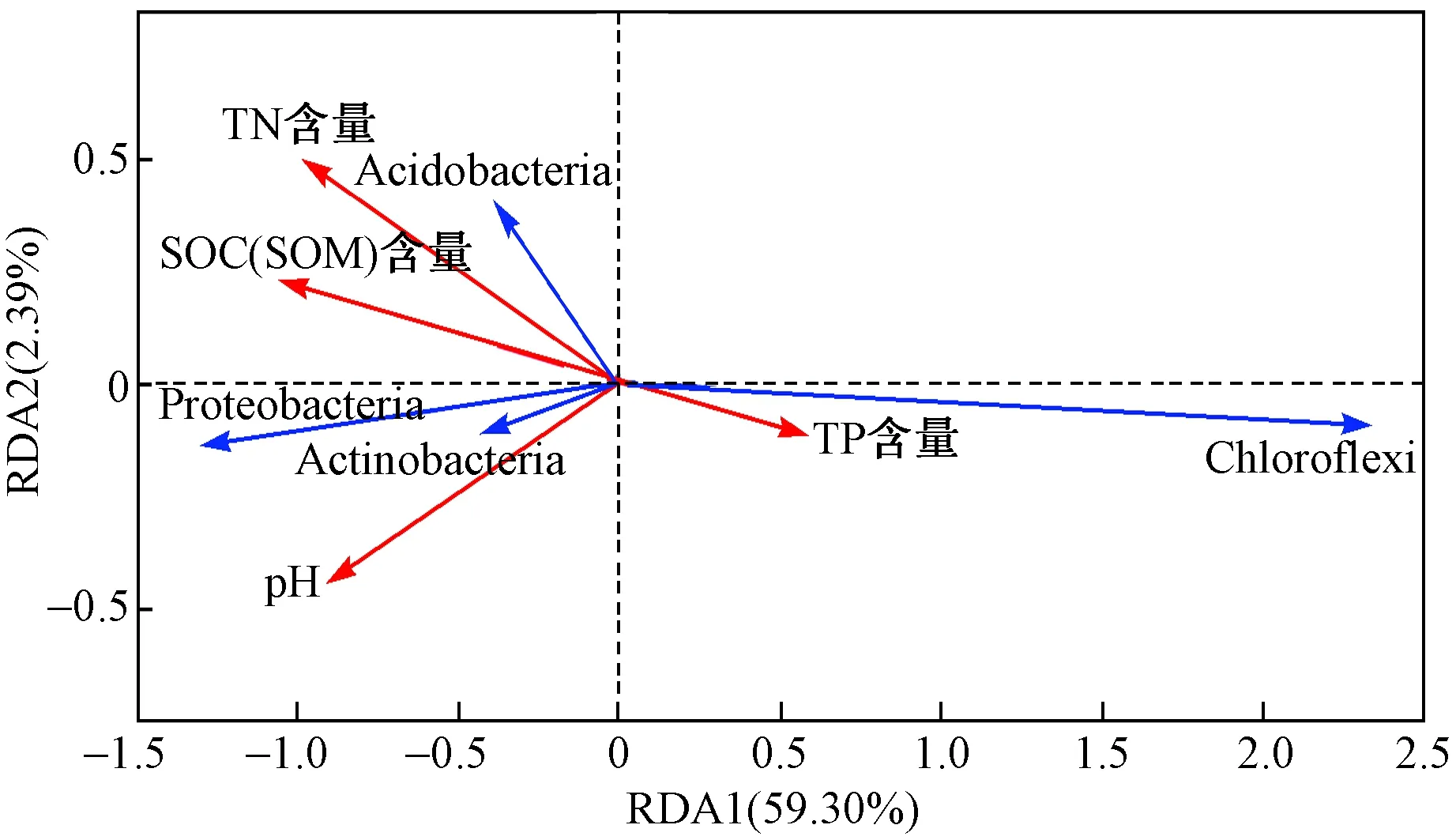
图6 不同水保措施下细菌门分类群落与环境因子的冗余分析结果Fig.6 Redundancy analysis of bacterial phyla and environmental parameters under different soil and water conservation measures
为了探索不同水保措施对土壤细菌群落交互作用的影响,笔者使用网络分析法分析了4种水保措施下土壤细菌的共生模式. 结果(见图7)显示,在不同水保措施下,土壤细菌群落交互作用网络共检测到21个细菌菌门,其中Proteobacteria、Acidobacteria、Actinobacteria和Chloroflexi为主要节点. 根据Layeghifard等[18]的研究,连通性最高的细菌门通常被认为是网络中的核心菌群. 笔者发现不同水保措施下土壤细菌交互作用的核心菌群均为Proteobacteria(见图7). 与JG相比,TD、QY和LZ土
壤细菌群落交互作用网络的节点数和负正相关连线数比较高,其中QY的节点数比JG多约47.6%(见表3),表明TD、QY和LZ对土壤细菌群落交互作用网络的影响均强于JG,尤其是QY. 特别重要的是,与TD和LZ相比,QY土壤细菌群落交互作用网络的节点数(186个)和连线数(283条)均较高(见表3),表明QY的网络更大、更复杂,且更高的平均聚类系数(0.348)和模块数(15个)使得节点之间的联系更加紧密,网络更加模块化. 而与QY相比,TD的平均聚类系数较低,仅为0.121,且LZ的平均路径长(2.08)和平均连通度(2.91)均较小(见表3). 此外,与TD、QY和LZ相比,JG土壤细菌群落交互作用网络的节点数最少,连线数却最高(见表3),特别是Proteobacteria、Acidobacteria和Actinobacteria之间的联系更紧密(见图7),表明JG可以促进细菌菌群之间的交互作用,尤其是合作.

表3 不同水保措施下土壤细菌群落的交互作用网络参数

注: 红线和绿线分别表示显著正相关(Spearman相关,P<0.05,R>0.8)和显著负相关(Spearman相关,P<0.05,R<-0.8);点的大小代表细菌相对丰度的多少.图7 不同水保措施下土壤细菌群落的分子生态网络Fig.7 Molecular ecological networks of bacterial communities under different soil and water conservation measures
3 讨论
3.1 水保措施对土壤细菌丰度和多样性的影响
该研究发现,与其他水保措施相比,狗牙根全园覆盖(QY)增加了表层土壤细菌数量,丰富了中表层土壤细菌种类,而狗牙根条带覆盖(TD)下中底层土壤细菌种类最多,二者是增加柑橘果园土壤细菌多样性的有效水保措施. 由于土壤微生物群落对生态系统服务具有重要作用[2],因而在土壤微生物群落中16S rRNA基因拷贝数较高是有益的. 与净耕果园(JG)相比,狗牙根全园覆盖(QY)显著提高了表层土壤中的TN、SOC和SOM含量,且土壤细菌16S rRNA基因拷贝数提高了近1.51倍(见图1). 狗牙根草通过固定大气中的氮,增加了土壤TN含量,这一结果与Reckling等[19]的报道相似. 植物残渣和凋落物会作为新鲜有机质被掺入土壤中,促进SOC和SOM含量增加并改善土壤质量. 草根和根系分泌物在SOC和SOM的积累中也起着十分重要的作用[20]. 相关性指数分析(见表1)显示,TN、SOC和SOM含量是表层土壤细菌16S rRNA基因拷贝数的驱动因素,因此狗牙根全园覆盖(QY)下表层土壤中具有更高的TN、SOC和SOM含量可以解释细菌生存和繁殖所需的营养来源.
虽然与其他水保措施相比,狗牙根全园覆盖(QY)下底层土壤细菌的Chao1指数最小,但其对底层土壤细菌多样性没有明显影响,这与ZHONG等[21]的结论存在差异. 而狗牙根全园覆盖(QY)中表层土壤细菌Shannon-Wiener指数更高,相关性分析(见图3)表明研究区的TN、SOC、SOM含量均与Shannon-Wiener指数呈显著正相关,这表明与其他水保措施相比,狗牙根全园覆盖(QY)增加了土壤中的TN、SOC和SOM含量,从而适度干扰了细菌群落,降低了种群之间竞争性生态位的排除和选择机制,增加了多种物种在不直接争夺资源的情况下紧密共存的可能性,碳源利用和代谢活性的改善最终促进了其中表层土壤细菌多样性以及物种丰富度的增加. ZHONG等[21]也认定全园覆盖是有益的. 与其他水保措施相比,狗牙根全园覆盖(QY)下土壤细菌群落在土壤剖面深度上的差异性最大(见图4). 这可能是由于在根际之外,土壤细菌的活性相对较小,草根分泌物有利于改善细菌多样性[22]. 狗牙根全园覆盖(QY)的草根根系密度较大,富含碳的狗牙根根系分泌物可能激活了土壤细菌的活性,因此具有更高的细菌多样性. 与其他水保措施相比,狗牙根条带覆盖(TD)下土壤细菌16S rRNA基因拷贝数虽较小,但其中底层土壤细菌Shannon-Wiener指数最高. 魏常慧等[23]亦表示条带覆盖的中底层土壤蕴藏着丰富的物种. 因此,笔者认为,狗牙根条带覆(TD)盖能改善中底层土壤细菌多样性,其较少的土壤扰动也意味着能更好地保护土壤结构,促进SOC固存和团聚体稳定. 而与狗牙根全园覆盖(QY)相比,经济作物萝卜-大豆轮作条带覆盖(LZ)表层细菌数量和多样性均较少,这可能是由于其土壤的频繁扰动增加了土壤中凋落物数量和根系分泌物,提高有机质输入量并降低了碳氮比,从而刺激表层土壤细菌群落的特定功能并降低了细菌多样性.
此外,相关性指数分析(见表1)表明,TP含量是底层土壤细菌16S rRNA基因拷贝数的驱动因素,因而与狗牙根条带覆盖(TD)、狗牙根全园覆盖(QY)和经济作物萝卜-大豆轮作条带覆盖(LZ)相比,底层土壤TP含量更高的净耕果园(JG)其底层细菌16S rRNA 基因拷贝数显著更大. 但与DONG等[10]研究结果不同的是,与狗牙根条带覆盖(TD)、狗牙根全园覆盖(QY)和经济作物萝卜-大豆轮作条带覆盖(LZ)相比,净耕果园(JG)各层土壤细菌的Shannon-Wiener指数均最低,这表明其对各层土壤干扰较大,土壤细菌均质化严重.
3.2 水保措施对土壤细菌群落组成的影响
该研究发现,不同水保措施改变了柑橘果园土壤细菌群落在土壤剖面上的组成,土壤细菌群落差异显著,TN、SOC和SOM含量(尤其是TN含量)是主要影响因素. 高通量测序结果(见图5)表明,4种水保措施下土壤细菌优势菌门中相对丰度最高的是Chloroflexi,其次为Acidobacteria、Proteobacteria和Actinobacteria,这与已有研究结果[24-26]一致. TN含量是4种水保措施下土壤细菌群落最主要的影响因素,因而Chloroflexi最高的相对丰度可以通过它对TN含量的高需求性来解释[27]. 值得注意的是,与狗牙根条带覆盖(TD)、狗牙根全园覆盖(QY)和经济作物萝卜-大豆轮作条带覆盖(LZ)相比,净耕果园(JG)土壤中Chloroflexi的相对丰度较大,冗余分析结果(见图6)表明,Chloroflexi的相对丰度与TN、SOC和SOM含量均呈显著负相关;与净耕果园(JG)相比,狗牙根条带覆盖(TD)、狗牙根全园覆盖(QY)和经济作物萝卜-大豆轮作条带覆盖(LZ)土壤中Acidobacteria和Proteobacteria的相对丰度较大,冗余分析结果(见图6)显示,上述细菌的相对丰度与TN、SOC和SOM含量均呈显著正相关. 以上分析表明,水保措施可以通过影响某些土壤特性来改变细菌群落结构,TN、SOC和SOM含量(尤其是TN含量)的差异是造成柑橘果园土壤细菌群落差异的主要原因. 这与DONG等[10]的研究结果相似,TN含量是影响不同种植系统中土壤群落差异的最重要因素之一. SOC和SOM在其他塑造细菌群落中的重要性也已有报道[28]. 此外,Chloroflexi是嗜热菌,在高温下生长良好,在有机物的分解中具有重要作用[29]. 柑橘果园土壤容易暴露在阳光下,因此Chloroflexi的相对丰度最高,尤其是净耕果园(JG). Chloroflex是多糖的主要降解菌,主要生长在低氧土壤中[30],在果园缺氧区多糖降解中起着重要作用. 净耕果园(JG)致密的表层土壤易将植物根部的生长限制在表层[31],从而导致土壤微生物的氧气供给不畅.
除Chloroflexi的相对丰度增加外,不同水保措施下Proteobacteria、Acidobacteria〔狗牙根条带覆盖(TD)除外〕和Actinobacteria的相对丰度均沿剖面深度呈减少趋势. Proteobacteria主要分布在植物凋落物和根系分泌物中,与高碳利用率有关,在碳循环中起关键作用,富营养特性明显[32-33]. 成长快速的富营养菌群丰度在富营养条件下更容易增加[34],因而Proteobacteria在营养状况较好的狗牙根全园覆盖(QY)表层土壤中更为丰富. 随着土壤深度增加,笔者发现中底层土壤逐渐转为贫营养的Chloroflexi,其在增加的同时也抑制了其他富营养菌的生长. 如果说碳是细菌的能量来源,那么氮则是养分循环的关键部分. Chloroflexi对土壤氮的需求很高,但无法固定氮[27]. 因此能矿化氮的Chloroflexi的相对丰度与土壤TN含量呈显著负相关(见图6),其可能与作物在争夺氮资源. Acidobacteria属贫营养细菌,是全球碳循环的重要参与者,不但具有降解复杂和难降解的碳化合物的能力,还能够利用NO3--N作为氮源[35-36]. 狗牙根条带覆盖(TD)中底层土壤TN含量较高,因而Acidobacteria的相对丰度沿剖面深度呈增加趋势. LIU等[37]发现TN、SOC和SOM含量均与Actinobacteria的相对丰度存在正相关关系,这与笔者所得研究相似. 但Sul等[38]观察到Actinobacteria在较低的SOC位点更为丰富,原因可能在于不同研究区的生态系统或SOC含量范围有所不同.
3.3 水保措施对土壤细菌群落交互作用的影响
该研究发现,Proteobacteria是4种水保措施下土壤细菌交互作用的核心细菌,狗牙根全园覆盖(QY)的土壤细菌群落交互作用网络的稳定性优于其他水保措施. 与其他优势菌门相比,作为拥有富营养优势的Proteobacteria可能具有更宽的生态位宽度和更高的抗干扰能力,在土壤细菌群落交互作用网络的稳定性维持中起着主导作用. 而其他非优势菌门可以作为多样化的库来增强细菌的抗逆性和抵抗环境干扰的能力[39]. 因此,水保措施可能会重塑土壤中复杂的细菌群落交互作网络. 模块数量增多,表明生态位越多,网络模块化可以增强网络在环境扰动下的稳定性[40]. 当4种水保措施中模块数量最多的狗牙根全园覆盖(QY)受到外界环境干扰时,细菌菌群更容易通过相互合作应对环境干扰. TIAN等[41]认为更大的网络复杂性可以稳定混合交互的细菌群落,提高资源的转移效率. 与经济作物萝卜-大豆轮作条带覆盖(LZ)和狗牙根条带覆盖(TD)相比,狗牙根全园覆盖(QY)的土壤细菌群落交互作用网络具有更大的网络直径、更长的平均路径和更大更复杂的网络(较多的节点数和连线数),这种变化增加了生态位宽度,增强了土壤微食物网中不同细菌之间的相互联系,有助于其更有效地利用土壤养分. 由于生态位重叠,土壤碳输入的选择效应可能加剧共存种群之间的养分竞争,而营养竞争是代谢能力相似的物种间拮抗互作的主要驱动因素之一[42],最大碳输入使狗牙根全园覆盖(QY)土壤潜在竞争对手之间的养分竞争加剧,土壤细菌群落交互作用的稳定性最好. 而与狗牙根全园覆盖(QY)相比,经济作物萝卜-大豆轮作条带覆盖(LZ)土壤细菌群落交互作用网络的平均连通度更小,狗牙根条带覆盖(TD)土壤细菌群落交互作用网络的平均聚类系数更低,这表明二者土壤细菌群落的交互作用网络易受外界环境的干扰,细菌群落内部的资源竞争在土壤中较弱,土壤细菌群落交互作用的稳定性较差.
此外,与狗牙根条带覆盖(TD)、狗牙根全园覆盖(QY)和经济作物萝卜-大豆轮作条带覆盖(LZ)相比,当负正相关连线数比较低的净耕果园(JG)环境发生扰动时,其土壤细菌群落的交互作用网络会在很短的时间内将环境扰动传递到整个网络,导致网络结构不稳定. 同时这种不稳定的网络可能导致参与土壤碳氮等养分循环的细菌物种群落发生显著改变[43],进而影响植被生长. 因而土壤底部养分较差的净耕果园细菌群落只能通过减少竞争和加强合作来应对环境压力,特别是Proteobacteria、Acidobacteria和Actinobacteria. 与其他3种水保措施相比,净耕果园(JG)虽然增加了网络的正负相关连线数量,但各层土壤细菌Shannon-Wiener指数均最低. 笔者推测,细菌多样性的减少可能会促进柑橘果园土壤中部分细菌(主要节点)之间相互作用的频率和幅度,但土壤细菌群落交互作用网络的稳定性随之变差. 净耕果园(JG)复杂的土壤细菌群落交互作用多集中在部分细菌(Proteobacteria、Acidobacteria和Actinobacteria)之间,而大多细菌物种被排除在交互作用之外,这可能导致了细菌多样性的丧失. 同时,这些较强的交互作用降低了细菌群落交互作用网络的稳定性,也为细菌群落间的交互作用、多样性和稳定性之间提供了一种机制联系[44].
4 结论
a) 与净耕果园(JG)相比,狗牙根条带覆盖(TD)、狗牙根全园覆盖(QY)和经济作物萝卜-大豆轮作条带覆盖(LZ)各土层土壤中的TN、SOC和SOM含量均呈增加趋势,狗牙根全园覆盖(QY)对土壤养分的贡献最大. 与经济作物萝卜-大豆轮作条带覆盖(LZ)和净耕果园(JG)相比,狗牙根全园覆盖(QY)增加了表层土壤细菌数量,丰富了中表层土壤细菌种类,狗牙根条带覆盖(TD)提高了中底层土壤细菌多样性,二者是增加柑橘果园土壤细菌多样性的有效水保措施.
b) 不同水保措施改变了柑橘果园土壤细菌群落在土壤剖面上的组成. 除Chloroflexi的相对丰度增加外,不同水保措施下Proteobacteria、Acidobacteria〔狗牙根条带覆盖(AD)除外〕和Actinobacteria的相对丰度均沿剖面深度呈减少趋势. 净耕果园(JG)导致厌氧细菌(Chloroflexi)的含量增加,后者可能与作物在争夺氮资源. 狗牙根条带覆盖(TD)、狗牙根全园覆盖(QY)和经济作物萝卜-大豆轮作条带覆盖(LZ)均提高了参与碳循环细菌的相对丰度,其中包括Acidobacteria和Proteobacteria. 不同水保措施下土壤细菌群落差异显著,TN、SOC和SOM含量(尤其是TN含量)是主要影响因素.
c) Proteobacteria是4种水保措施下土壤细菌交互作用的核心细菌,狗牙根全园覆盖(QY)的土壤细菌群落交互作用网络的稳定性优于其他水保措施.
d) 狗牙根全园覆盖(QY)提高了土壤中的TN、SOC和SOM含量,增加了土壤细菌多样性,增强了土壤细菌群落交互作用的稳定性. 因此,笔者鼓励采取狗牙根全园覆盖(QY),这一结果将进一步为土壤生态系统研究提供参考. 但狗牙根全园覆盖(QY)对土壤的细菌多样性和细菌群落交互作用等影响非常复杂,在不同的土壤类型和气候条件下是否具有普遍有效性有待于进一步研究.
猜你喜欢
山西果树(2022年6期)2022-11-30 22:49:34
中国水土保持(2022年4期)2022-04-07 01:48:30
当代水产(2021年4期)2021-07-20 08:10:48
建材发展导向(2021年9期)2021-07-16 07:10:50
农村农业农民·B版(2018年1期)2018-01-31 20:51:48
水土保持研究(2016年3期)2016-10-28 07:44:42
浙江林业(2016年6期)2016-07-05 06:39:36
中国当代医药(2015年20期)2015-03-01 02:04:33
西南军医(2015年1期)2015-01-22 09:08:26
河南科技(2014年5期)2014-02-27 14:08:50
