
Mature stand developmental stage has ceased to constitute the most suitable habitat for the capercaillie in the Augustów Forest,Poland
2021-02-28 09:13:20StanisawDrozdowskiDorotaZawadzkaGrzegorzZawadzkiMarcinStudnickiandBogdanBrzeziecki
Forest Ecosystems 2021年4期
Stanisław Drozdowski,Dorota Zawadzka,Grzegorz Zawadzki* ,Marcin Studnicki and Bogdan Brzeziecki
Abstract
Keywords:Habitat suitability,Lek occupancy,Logistic regression,Lowland forest,Stand structure,Tetrao urogallus
Background
Forest management has a profound impact on forest habitat conditions (Angelstam 1996; Battles et al. 2001).It leads to forest fragmentation, the disappearance of a forest’s natural structure, the shortening of the stand developmental cycle, and a reduction of the area of oldgrowth stage (e.g. Angelstam et al. 2004; Lõhmus et al.2005; Zawadzka et al. 2016). Other effects include a reduction of the volume of dead wood as well as an elimination of natural disturbance patterns and structures(Linder and Östlund 1998; Lindenmayer and McCarthy 2002; Bujoczek et al. 2018). Birds are an important group of organisms affected by forest practices in many different ways (Scheringer and Schumacher 2004; Basile et al. 2019). The birds which are most sensitive to habitat changes are highly-specialised species, especially those connected with mature forests (Rosenvald and Lõhmus 2003; Roberge and Angelstam 2006; Czeszczewik et al. 2014; Zawadzka et al. 2018). The maintenance and conservation of forest biodiversity, including rare bird species, is one of the significant challenges of modern forestry(Basile et al. 2016).
In Europe,one of the most endangered forest bird species is the capercaillie Tetrao urogallus, which is considered an iconic species serving an “umbrella” role(Pakkala et al. 2003; Lõhmus et al. 2017). The capercaillie is a rare sedentary forest grouse species that lives in the old, natural forest complexes of a larger size, occurring in certain boreal and mountainous regions of Europe (Storch 2001, 2007; Jahren et al. 2016). It has proved to be among the birds which are most sensitive to the anthropogenic changes of forest habitat, not least due to a large amount of living space required by both individuals and the whole populations of this species(Rolstad and Wegge 1987; Angelstam 2004; Angelstam et al. 2004; Wegge and Rolstad 2011). The global climate warming can also have a negative impact on the grouse population (Moss et al. 2001; Storch 2007; Poirazidis et al. 2019). As a result of the anthropogenic pressure related to habitat loss, forest management, and hunting, the last two centuries witnessed the species’ retreat from large parts of both Western and Central Europe (Tomiałojć 2000).
Population declines among capercaillies have, in fact,been observed in almost all European countries for some time now.However,the process has been most rapid in the central and southern parts of the species’fragmented European range (Zawadzka and Zawadzki 2003; Storch 2007).The capercaillie is listed in Annex 1 of the EU’s Birds Directive, and is also Red-Listed as an endangered species– and under strict protection – in 21 European countries,including Poland(Głowaciński 2001;Storch 2007).
Capercaillie extinctions and population declines are both associated with habitat loss and fragmentation, as well as intensified land-use by people (e.g. in sport, tourism and recreation, and forest management) (Storch 2007, 2013; Thiel et al. 2011; Moss et al. 2014; Coppes et al. 2017, 2018). The major reasons for changes in the habitat of the species are considered to relate to the development of commercial forestry, where large areas are clear-felled (Saniga, 2003; Wegge and Rolstad, 2011;Mikoláš et al., 2015; Kämmerlea et al. 2020). The remaining areas of old-growth forests are too small and isolated, and extensive areas of young and middle-aged stands are characterised by unsuitably high densities of trees(Sirkiä et al.2011).In general,the negative influences of forestry manifest mostly in the destruction of lekking sites along with the surrounding territories occupied by males, as well as, partly, in overall habitat loss and forest fragmentation (Rolstad and Wegge 1987; Miettinen et al.2010; Wegge and Rolstad 2011; Mikoláš et al. 2015).Changes in the structure of forest landscape, induced by forest management, affect the distribution of leks and the area occupied by the capercaillie (Angelstam 2004; Zizas et al. 2017). Besides the direct influences (such as clearcutting in lek areas), modern forestry has affected populations of the capercaillie also in an indirect manner,namely through its tendency to cause change in forest structure,age, and stand composition (Broome et al. 2014;Braunisch et al. 2019). In forests which are managed,natural disturbances – characteristic of unmanaged old-growth forests that do favour the formation of a forest structure suitable for the capercaillie – play only a minor role (Mikoláš et al. 2017). The widespread fencing of forest plantations results in grouse collisions. Locally, it can represent an important cause of mortality, as is the case in Scotland (Baines and Summers 1997). Collisions with fences as a cause of mortality of the capercaillie have also been documented in Poland (Zawadzka 2014).
In the Carpathian Mountains, selective logging at low intensity was reported as affecting the population of the local capercaillies in a positive way. In contrast, the proportion of forest clear-cuts in an area as well as the intensity of tourism are both seen to exert a significant negative influence on the numbers of males present at leks (Mikoláš et al., 2015).
In recent decades, several studies have made it clear that lekking is not at all confined to areas of a mature forest (Rolstad et al. 2007). The occurrence of the capercaillie is seen to be conditioned not only by the specific stand structure in the area of the lek, but also by the quality of forest habitat analysed on the landscape scale(Storch 2002; Sirkiä et al. 2011; Brzeziecki et al. 2012;Zizas et al. 2017).
The identification of habitat requirements of the capercaillie is a precondition for its effective conservation.The same can be said about the co-existing species,whose needs are less demanding but which benefit from the conservation measures adopted with regard to the capercaillie, which makes it a recognised umbrella species (Suter et al. 2002; Pakkala et al. 2003). Recently,studies on forest grouse, mainly the capercaillie, has focused on the ecological requirements, particularly habitat preferences (Moss et al. 2010). Inadequate attention to the capercaillie’s habitat requirements is among the reasons for failures to achieve an effective reintroduction of the species (Merta et al. 2015).
Studies of habitat preferences have been conducted in order to determine the probability of the occurrence of the capercaillie. Work has also been done on areas occupied by still-relatively-large populations (Storch 2002;Braunisch and Suchant 2007; Teuscher et al. 2011), as well as by ones heading for extinction (Graf et al. 2005,2007; Bollmann et al. 2011; Mikoláš et al., 2015). There are two emerging approaches to the assessment of habitat for capercaillies. In mountainous areas of Central Europe, the main kind of work uses mathematical modelling to identify habitat features associated with the highest probabilities of the species being present (e.g.Storch 2002; Bollmann et al. 2005, 2011; Braunisch and Suchant 2007; Graf et al. 2005, 2006, 2007; Teuscher et al. 2011). In Fennoscandia and the Baltic countries,there have been analyses of the long-term influence of structural change induced by forest management on the distribution of leks and changes in numbers of the birds(Gregersen and Gregersen 2008; Miettinen et al. 2010;Sirkiä et al. 2010,2011; Wegge and Rolstad 2011;Leivitis 2014; Lõhmus et al. 2017; Zizas et al. 2017).
In the work presented here, the emphasis has been put on a detailed investigation of the habitat requirements of the species, which has been assessed in relation to the small and isolated natural population (of ca. 50–80 individuals) still living in the lowlands of North-Eastern Poland, in the managed part of the Augustów Forest.Based on the long-term monitoring of the capercaillie on the leks (Zawadzki and Zawadzka 2008, 2012), a total of 18 leks (9 active and 9 abandoned) were selected for field measurements. We investigated hierarchy and importance among 13 variables characterising the forest habitat within a 1-km radius of each of those 18 leks. At the latter stage, we considered the absolute and relative significances of each of the above variables,using a logistic regression with regard to the current status of leks within the managed forest as either abandoned or still occupied.
We hypothesised that changes in habitat structure at the stand scale were the key reason for the disappearance of the capercaillie from some locations in the Augustów Forest. Our expectation was that the identification of the importance of particular habitat features might prove to be useful in formulating recommendations concerning the active protection of habitat suitable for the capercaillie in the context of modern forestry.Moreover,the recognition of the most important variables characterising stand structure would help to determine the probability of the occurrence of species in any particular part of the forest. Our study would,therefore,offer indirect help with the assessment of the impact of forest management and other factors on the capercaillie’s habitat conditions, at the same time providing a basis for recommendations concerning habitat management.
Materials and methods
Materials
The study area and data collection
The Augustów Forest is the home of one of the country’s four isolated populations of the capercaillie. The Polish part of the Augustów Forest is located in the north-eastern part of the country (at 23°15′ E, 53°54′ N,Fig. 1) and extends over 1140 km2. The area is fairly flat,with a range of elevations between 135 and 190 m a.s.l.The climate is relatively cold, with a mean annual temperature of 6.5°C, a 135-day growing season, and snow cover typically present for around 100 days a year.Forest cover here is of around 93%, while lakes account for a further 6% of the area. Tree stands are dominated by Scots pine (Pinus sylvestris) (78%), Norway spruce(Picea abies) (8%), black alder (Alnus glutinosa) (9%), silver birch (Betula verrucosa) (5%), and pedunculate oak(Quercus robur) (1%). The understorey is formed mainly by spruce, while the bush layer consists mainly of young spruces and common junipers (Juniperus communis),with the cover of these varying from one stand to another. The mean age of tree stands here is 65 years, with stands older than 100 years accounting for about 15% of the overall area of forest. The mesic pine forest site type,distinguished by a big share of bilberry (Vaccinium myrtillus), accounts for almost 40% of the area, while 27% is occupied by mesic mixed coniferous forest site type with bilberry and bush-grass (Calamagrostis epigeios). Some 7% of the forest area is classified as a wet alder forest.The Augustów Forest is included in Europe’s Natura 2000 network as the Special Protection Area for Birds PLB200002 “Puszcza Augustowska”. Most of the area comprises managed stands supervised by six Forest Districts of Poland’s State Forests National Forest Holding.Over the past several hundred years, this forest has been intensively managed with the use of a clear-cutting system. Clear-cuts were regenerated by either natural seeding (by leaving seed trees) or by direct seeding and/or planting.The study area has not been impacted by major natural disturbances, with these affecting small areas at most (Reindl 1965; Sokołowski 2010).

Fig.1 Study areas in the Augustów Forest(North-Eastern Poland).Black points –active Key Areas around leks,open points–abandoned KeyAreas around leks. The area of the whole square is ca.4 km2. The measurements were conducted on circular plots(r=15 m radius,located across a 200 m×200 m grid)
Data on the locations of lekking sites within the Augustów Forest, history of each lek and number of cocks attending, as well as on their present status (occupied or abandoned) was taken from Zawadzki and Zawadzka (2008, 2012). The spatial distribution of leks and the number of displaying cocks were determined during the 15-years-long monitoring. Every spring all known leks were visited three times in order to determine the number of capercaillie males (Zawadzki and Zawadzka 2008, 2012). The main fieldwork was carried out in the area of 9 active leks as well as 9 abandoned by the capercaillie over the last 25 years (6 less than 10 years ago). The study plots covered key areas located within a 1-km radius from the studied leks (Fig. 1). This size of the key area was recognised as a territory of the utmost importance for the proper functioning of the capercaillie population (Wegge and Larsen 1987; Wegge et al. 2013).
The assessment of the habitat was performed within the Key Areas, on circular plots positioned across a 200 m×200 m grid (Fig. 1). Each plot had a radius of 15 m and thus covered 706 m2. We established between 53 and 133 study plots in each Key Area (depending on the local conditions, e.g. clear-cuts area, dense young plantations, roads), with the average amounting to 98; the area was spread over 4 km2. We excluded areas of open water and bogs. The nine Key Areas including active leks were represented by a total of 1032 circular plots, while there were 747 plots in the 9 Key Areas associated with abandoned leks.
Stand and site variables
Measurements or estimations made on all plots included 13 habitat features (variables) (Table 1). The description of the tree stand involved a determination of its age(ATS), tree stand structure (vertical layering) (TSS),stand density (TN), and the percentage share of Scots pine (PSP). Other recorded features included the canopy(crown) closure of the upper stand layer (CCUL) and the closure of the second stand layer (CCSL) as a measure of the amount of light penetrating through to the forest floor. The canopy closure was determined by the visual estimation from the ground – summing up the crown areas for all trees on a fixed plot area and dividing them by the ground area. We used a 5-rank scale: Open (1; 0–30% of ground covered by crowns), Loose (2; 31%–50%),Discontinuous (3; 51%–70%), Moderate (4; 71%–90%),Full (5; >91%). Other measured or assessed variables included the classes of soil-moisture (SM) and soil fertility(SF), the shrub cover (PCS), the bilberry cover (PCB),the height of ground-vegetation (GVH), the distance from roads used to transport timber (DFR), and the presence of specific habitat elements (PHE). In the latter case, we considered the occurrence of:1) large trees with horizontal branches (good for perching); 2) dead fallen trees; and 3) anthills. Values for the analysed features on scales – as detailed in Table 1 – were estimated and noted in the field, with the information contained in the BDL forest data bank (available at https://www.bdl.lasy.gov.pl/portal/mapy-en) also being used.
The values of some explanatory variables (the share of Scots pine, tree stand structure, canopy closure, the cover of shrub layer, the cover of bilberry, ground vegetation height, and distance from roads) obtained ranks that increased with greater magnitudes of the given feature (e.g., the greater the percentage of the cover of bilberry, the higher the designated rank – Table 1). In this way, the importance of a higher level of a given parameter for the suitability of the habitat for the capercaillie was reflected. In the case of stand age, soil moisture,and soil fertility, values were given to labels based on the measured features of the stand that had been inventoried in the sample plots. These were chosen in such a way as to point to the characteristic habitat conditions associated with the phase of forest development (sapling, pole,early mature, mature forest, late mature), as well as trophic (oligo-, meso-, eutrophic) and moisture (dry,fresh,wet, bog) conditions of the soil. Thus, for example,stand aged between 1 and 10 years corresponded with the developmental phases of a young plantation, while an age over 100 represented a mature forest. There was,therefore, a correspondence with the structure characteristic of a given age-related developmental stage (Table 1).It was assumed that these variables would be nominal in nature. The assignment of different values to labels describing habitat features (variables) was based oninformation in the literature as well as on expert knowledge relating to the habitat requirements of the capercaillie.

Table 1 Variables used to describe the capercaillie’s habitat in the Augustów Forest
Methods
First, the distribution of sample plots in classes corresponding with the occupied and abandoned Key Areas was compared using the χ2test. Subsequently, the occupied and abandoned Key Areas were compared by means of logistic regression, with a binary dependent variable taking on values 0 (area abandoned) or 1 (area occupied). This method was used because of the fact that logistic regression does not set out assumptions regarding the nature of explanatory variables (which can be either ordinal or nominal). Moreover, the distributions of independent variables (assessed features of the forest) do not have to conform to a normal distribution(Kleinbaum et al. 1982; Hosmer and Lemeshow 1989).Binary logistic regression is a common statistical method used in conservation biology to estimate probabilities of occurrence in relation to different predictors (Kleyer et al. 1999; Cowley et al. 2000).
The general logistic regression model was:

where Y is the dependent variable, Xi(i=1–k) are the independent variables, α is the intercept, and βiare the coefficients.
In order to avoid a strong multicollinearity between predictor variables, we studied the correlation matrix for 10 out of 13 predictors (after excluding the three nominal variables: stand age, soil moisture, and soil fertility)(Table 2). If there was a strong correlation between two variables (Spearman rank correlation, |rs|>0.70), only that parameter which is seemingly more important for the species (in line with the literature and expert knowledge) was selected for further modelling (see Hosmer Jr and Lemeshow, 1989). The first analysis used all the non-co-linear explanatory variables.It was followed by the identification of a subset of explanatory variables characterised by the greatest predictive power, i.e., those assessed features of forest which were differentiating between occupied and abandoned KAs in the most adequate manner. We applied the Akaike Information Criterion to the selection of variables(Akaike 1974;Burnham and Anderson 2002), while general model performance was assessed by calculating the area under a receiver operating characteristic curve (AUC), as well as model sensitivity and specificity(Hanley and McNeil 1982).We carried out all statistical modelling with the R 3.6.1 packages stat and lme4, using a generalised linear model (GLM) for binary response variable(R Core Team 2019).
Results
Habitat differences between active and abandoned leks
The distribution of sample plots for age classes shows that in both categories of Key Areas stands over 30 years old occurred most frequently (Fig. 2a). In the Key Areas still occupied by the species, stands aged 30–60 years were prevailing. In contrast, the abandoned Key Areas were dominated by forest stands older than 60 years(χ2=23.95; p=0.002). In the studied Key Areas, singlelayered stands dominated, while two-layered stands were limited (Fig. 2b). Stands in active and abandoned Key Areas differed in terms of structure (χ2=27.05; p <0.001), with the occupied ones dominated by singlelayered stands.
Occupied Key Areas showed bigger shares of stands with more than 90% Scots pine (χ2=21.99; p=0.002)(Fig. 2c). Canopy closure of the upper stand layer resembled a normal distribution (Fig. 2d). Abandoned stands have a higher canopy closure than occupied stands (χ2=28.57; p=0.001). Stands within active Key Areas havemainly a single-layered structure (χ2=28.57; p=0.001),while the share of stands with a well-developed second layer was bigger in the abandoned Key Areas (χ2=34.58;p <0.001) (Figs. 2e, 3a, 3b).
综上,我们可以得知,对于生产性服务业一般纳税人而言,其“营改增”前后税负变化主要取决于其购进项目的多少,且购进固定资产比购进服务对税负下降的影响更大;税率为6%的一般纳税人“营改增”对其税负的影响程度远远小于税率为11%的一般纳税人,“营改增”后税负下降可能性大大高于税率为11%的一般纳税人。在此不再赘述。
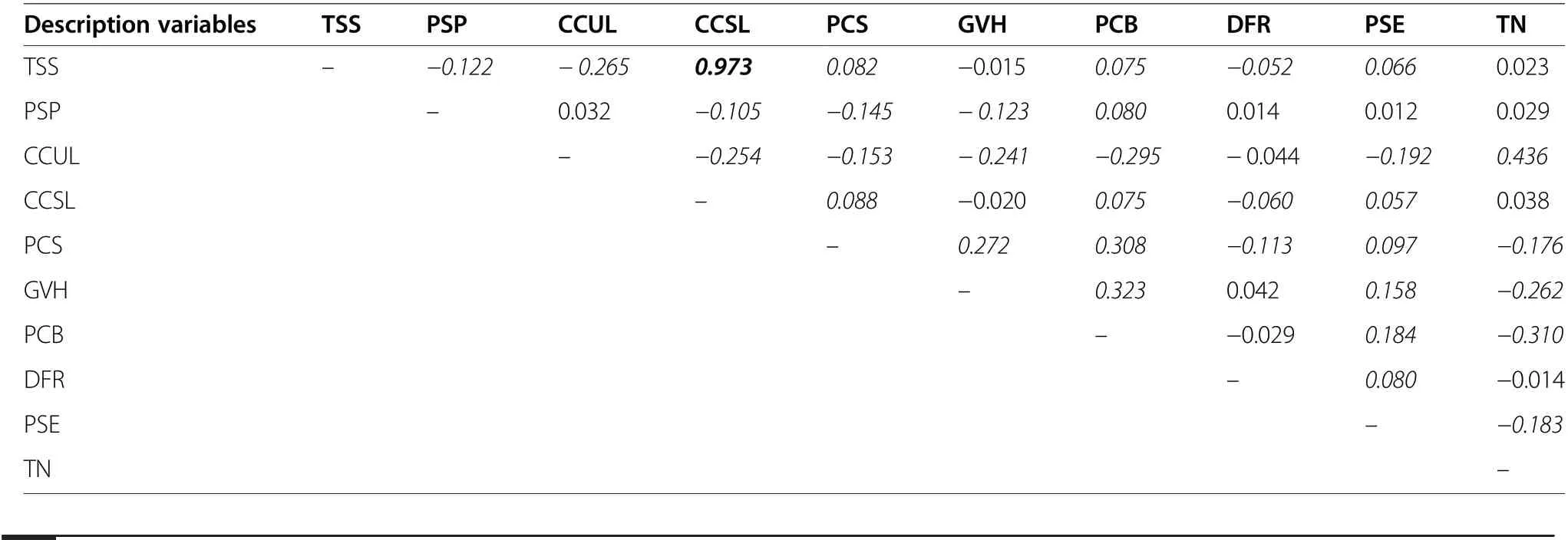
Table 2 Spearman Rank correlation matrix for variables describing the capercaillie’s habitat (Relationships statistically significant at α=0.05 are indicated by the use of italics strong correlation(Spearman rank correlation,|rs|>0.70)between two variables is indicated by the use of bold)

Fig.2 Percentage shares of sample plots in still-occupied or abandoned refuges for variables(Table 1)describing Key Areas for the capercaillie

Fig.3 The comparison of stand structure in Key Areas that are active(a)or abandoned(b)
The occurrence of shrub and ground-vegetation layers was skewed to the left (Fig. 2f), meaning that undergrowth was, most typically, only poorly-developed in the Key Areas. Areas with only a poorly-developed shrub layer occurred much more often in the still occupied Key Areas (χ2=34.58; p <0.001). On most of the plots,ground vegetation did not exceed 40 cm in height. However, the height interval 30–40 cm was mostly present in the abandoned Key Areas (χ2=94.72; p <0.001), while the still-occupied ones were dominated by shrubs, whose height was below 30 cm (Fig. 2g). The occupied Key Areas featured the prevalence of bilberry cover shares between 1% and 30%, while the abandoned Key Areas featured the share of bilberry cover that often exceeded 30% (χ2=65.2098; p <0.001) (Fig. 2h). Mesic forest site types occurred most often in both occupied and abandoned Key Areas (χ2=4.16; p=0.245), while the link between the age-class structure and soil humidity proved very similar in both variants (Fig. 2i). A somewhat greater share of nutrient-poor site types characterised the stilloccupied Key Areas,as opposed to the ones abandoned by the capercaillie(χ2=21.96;p <0.001)(Fig.2j).
We have found that distance from frequently used forest roads points to the prevalence of plots at a distance (>150 m)among the still-occupied Key Areas,as opposed to the abandoned ones(χ2=20.34;p=0.001)(Fig.2k).
The numbers of special habitat elements proved to be similar in areas representing both categories (χ2=3.03;p=0.387) (Fig. 2l). Nevertheless, it was noteworthy that 33.2% of sample plots featured all these three structural elements identified as important to the capercaillie.
The numbers of trees with DBH ≥7 cm per hectare were in the range from 0 (i.e., no such trees on plots located in newly-established plantations) to around 16,000 per hectare among the abandoned Key Areas, and to some 14,300 in the occupied ones. Nonetheless, respective mean values were of just 844 and 876 trees·ha-1. All that can be said about this feature is that it proved to be more variable in plots representing the Key Areas abandoned by the capercaillie, as opposed to the stilloccupied ones (V=118% and 101%, respectively).
The logistic regression model
Our inspection of the habitat correlation matrix revealed a very high correlation between stand structure and canopy closure of the second stand layer (r=0.97). This ensured a limitation of further analysis to canopy closure of the second stand layer (Table 2).
The first parametrisation involved the full model containing all non-collinear variables, with the aim being to determine the influence of each explanatory variable on the modelled phenomenon. It emerged that higher values for canopy closure of the upper and the second layer, the cover of shrubs, the cover of bilberry, and stand density were associated with a reduced probability that a Key Area would still be found to be occupied by capercaillies, whilst higher values for proportion of Scot pine, the height of ground vegetation, distance from roads, and the presence of habitat elements were all associated with a higher probability of the Key Area being occupied (Table 3). In the case of the variable describing a forest’s developmental stage, and with the mature stage chosen as a point of reference, capercaillies were categorised as less likely to be present in newly-planted areas and in young plantations and stands in the polewood stage, whereas they were more likely to occur in stands aged 30–60 or 60–100 years. A particularly high value for OR was obtained for stands that were 30–60 years old, with capercaillies being 1.6 times as likely to be present there as in the mature stage.
The reference variants for soil moisture and soil fertility were taken to be the wettest (boggy) soil and the soil of the highest fertility, respectively. Lower moisture and/or fertility were associated with a higher probability of the presence of capercaillies. The chance (OR) of the species being present in the forest on dry soil was 2.4 times as high as in the reference conditions, while the corresponding figure was even 7.3 times as high where soil was poorer in nutrients, as opposed to the most fertile one (Table 3).
In fact, the model has six explanatory variables that achieve statistical significance, i.e., the stand age, the share of Scots pine, upper canopy closure, the cover ofshrubs, and the cover of bilberry. The variables best distinguishing between the occupied and the abandoned Key Areas for the capercaillie proved to be shrub layer(OR=0.111) as well as the extent of the closure of the second canopy layer (OR=0.441). Only a slightly less considerable influence was found to be exerted by the stand age. The full model is characterised by high values for the ROC area beneath the curve (AUC=0.72), with a high degree of specificity (73.3%), although with low sensitivity (18.9%) (Table 4).
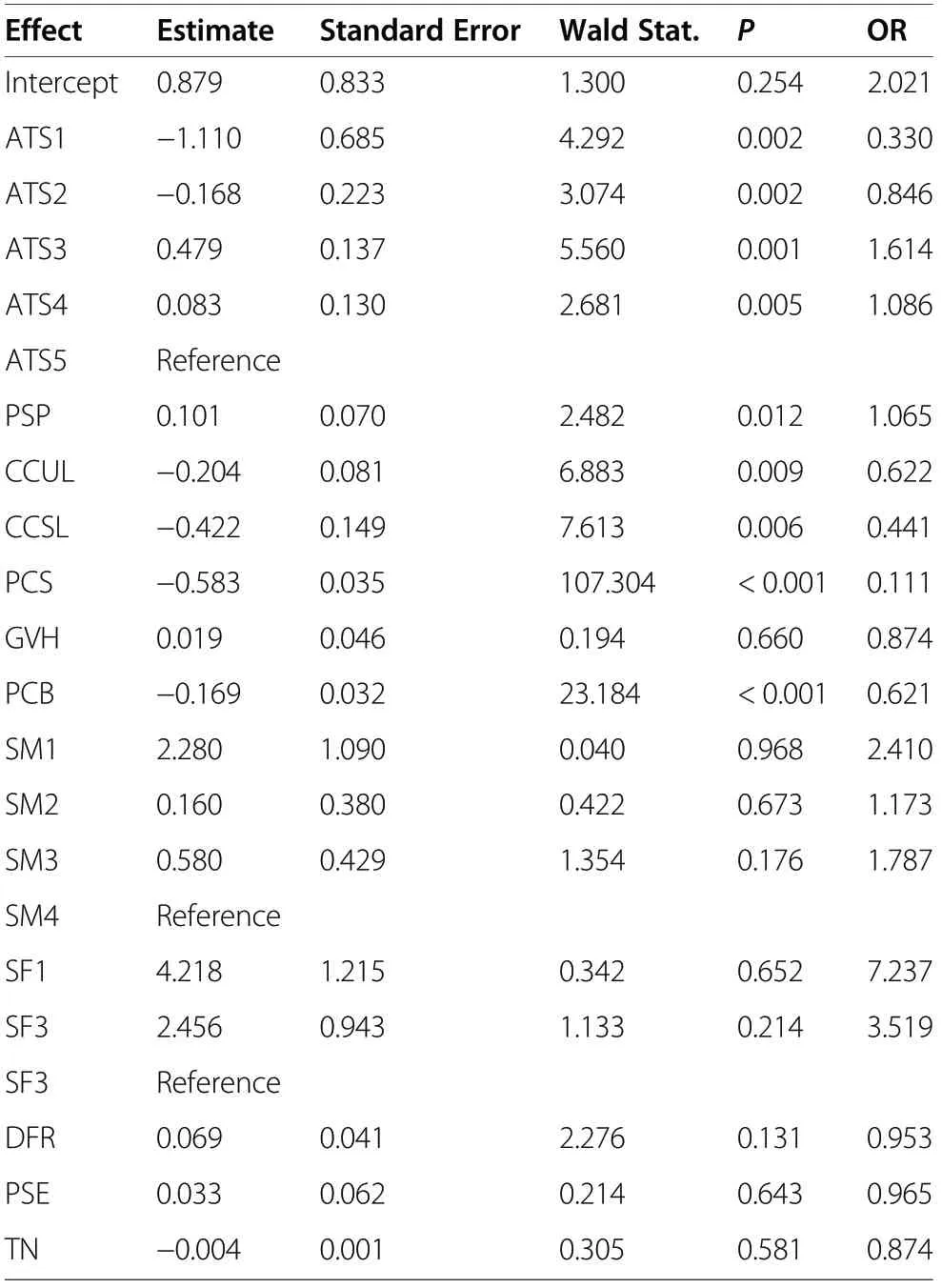
Table 3 Results of a logistic regression analysis –full model including all explanatory variables but stand structure
In order to determine the optimal model – containing the variables that are best able to distinguish between Key Areas still occupied or else abandoned by capercaillies – all the combinations of subsets of explanatory variables were analysed. The model characterized by the lowest value for the AIC criterion was made up of following variables: the stand age, upper canopy closure,the cover of shrubs, and the cover of bilberry (Table 5).It further emerged that the variables of the greatest discrimination power involved the presence of the two layers of undergrowth (shrubs and ground vegetation)?)(OR=0.325), the closure of the second canopy layer(OR=0.369), the closure of the upper layer (OR=0.565), and stand age as a manifestation of a developmental stage (Table 5). A greater cover by the lower stand layer and a higher density of trees in either the first or the second layer had a negative effect on the suitability of the habitat. Hence the chance of capercaillies being present in stands aged 30–60 years was 1.8 times as high as in the case of the mature forest, whereas it was lower in stands aged 60–100 or 10–30 years, and the lowest at all in newly-established plantations (Table 5). The cover of bilberry was also the subject of a significant relationship (PCB, OR=0.687). The share of an occupied area accounted for by bilberry was often lower than on plots already abandoned. All the explanatory variables forming part of the optimal model achieved statistical significance. The optimal model was characterised by the presence of the AUC – an area under the ROC curve of 0.709, with a very high level of specificity(91.9%), but very limited sensitivity (5.3%) (Table 6).This, in turn, ensures that the model offers a very clear indication of places with features of the biotope suitable for the capercaillie.
Discussion
The capercaillie has been commonly recognised as a bird species which needs different successional forest stages,but which prefers old, mature stands (Rolstad and Wegge 1987;Klaus et al.1989;Storch 2001).Stands suitable for the capercaillie are characterised by a loose or moderately-dense canopy cover, a well-developed layer of bilberry bushes covering the ground, the presence of patches of trees regenerating naturally, as well as the occurrence of small open areas (Storch 2001, 2002).
Field-based observations of the active and the abandoned Key Areas – supported by the results obtained by the means of the logistic model as well as other studies from the Augustów Forest (Brzeziecki et al. 2012;Zawadzka et al. 2018) – suggest that capercaillies present there occupy mainly single-layered, poor coniferous stands, with an intermediate canopy cover, strongly dominated by Scots pine, and a very limited admixture of spruce. In the active Key Areas, the cover of shrublayer rarely exceeded 10%, while in the abandoned Key Areas this level was, on average, much higher. Another finding, and a rather surprising one, concerned the preference for fresh coniferous forest site types. Over the last 20 years, the capercaillies living in the Augustów Forest have, indeed, abandoned the Key Areas located in moist or swamp coniferous forest site types, while all the leks remaining active at the time of our study were located in mesic pine forest sites. The only exception involves an old and extensive swamp pine forest, which has been in use by capercaillies for more than 90 years now (Domaniewski 1933;Zawadzki et al. 1999).
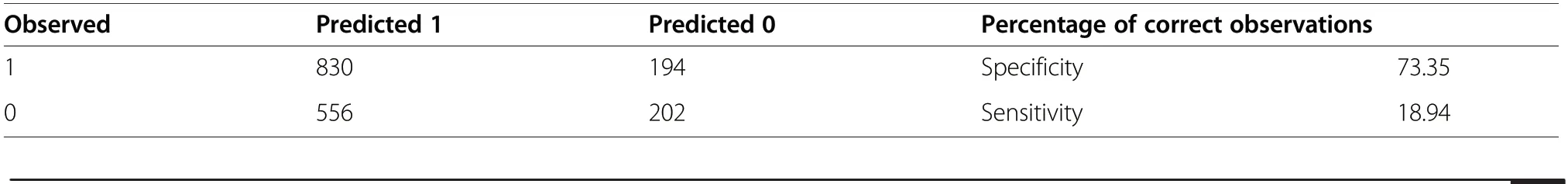
Table 4 Confusion matrix for the full model (OR=3.35, Log OR=1.21), 1 –Key Areas still occupied by the capercaillie (active), 0 –Key Areas abandoned by the species

Table 5 Results of a logistic regression analysis –model parametrised by the method of searching all the possible combinations using the AIC criterion
Both the full and the optimal logistic regression models were characterised by rather high levels of explanatory or predictive power, to the extent that they might be used to indicate areas more or less suitable for the species – e.g., with regard to the proposed efforts to resettle or reintroduce capercaillies – as well as management-related considerations as to where forests continue to fit in in terms of their economic usefulness.Specificity values proved to be particularly high (at 92%),indicating a great extent of suitability of the model when it comes to an appropriate habitat being identified. In contrast, the model sensitivity was much lower, which is to say that it is far less efficient at rejecting sites or, to be more precise, at pointing to those places that are not suitable for the capercaillie.
We explored the capercaillie’s habitat preferences within a 1-km radius. According to Storch (2002), such an approach, lacking characterisation at the landscape level, does not offers full opportunities for the quality of the capercaillie’s habitat to be assessed. Nevertheless,our results confirmed the hypothesis that changes in the habitat observed at the patch scale can represent important reasons for capercaillies to abandon some leks in the Augustów Forest. It results that by far the most important factors (differentiating most clearly between occupied and abandoned Key Areas) are linked to a forest’s vertical structure. Particularly, the presence of dense undergrowth has significantly reduced the probability of the occurrence of the capercaillie. The developed logistic regression model indicated that the filling of space in the forest with trees and shrubs – and hence an increase in overall stand volume – is inferred to do a lot to reduce the probability that capercaillies will be present.Such features are, of course, closely connected with forestry management in temperate forests, in which a high stocking rate is regarded as essential to both productivity and even the technical quality of trees producing timber(Oliver and Larson 1996). With this in mind, it is either the case that underplanting takes place in forests, or efforts are made to nurture the naturally-occurring undergrowth (Silvicultural Guidelines, 2012). In addition,installed or naturally-developing layers of undergrowth in nutrient-poor (oligotrophic) sites – such as the poor pine forest sites present in the Augustów Forest – are being stimulated anyway both by the global climate warming and by the ongoing habitat eutrophication. In the case of the Augustów Forest, the annual deposition of nitrogen is at the level of 10–12 kg·ha-1per year(Kasperowicz and Wacław 2016), and this alone suffices to ensure that forests here are different from those from several decades ago. This is particularly true for the older phases of stand development, which historical data reveal as once less resource-rich, with spruce and broadleaved trees absent from lower stand layers, and with ground cover formerly typical of coniferous forest, consisting of a fairly loose cover of bilberry (Ossendowski 1936; Andrzejczyk and Brzeziecki 1995; Szczygielski 2007; Solon and Matuszkiewicz 2008; Paluch and Gabrysiak 2019). This has been documented in Fig.3.
Today, the ongoing effects of eutrophication in the Augustów Forest include the presence of spruce in what had previously been nutrient-poor site types, with a closed understorey or the second stand layer of thisspecies. Succession involving spruce – and even broadleaved trees such as birch and the hazel Corylus avellana– is changing forest structure (including the oldest developmental stages of pine forest) in a manner unfavourable to capercaillies, even where the site had previously been nutrient-poor. This would seem to explain why today’s capercaillies in the Augustów Forest most often occupy forest stands that are 30–60 or 60–100 years old,and why the phase of development proved to have such a great and significant influence in the context of the overall model.Along the forest age gradient,competition eases and the closure of the stand is more open, which makes it possible for the undergrowth to develop in stands that are 30–60 years old.
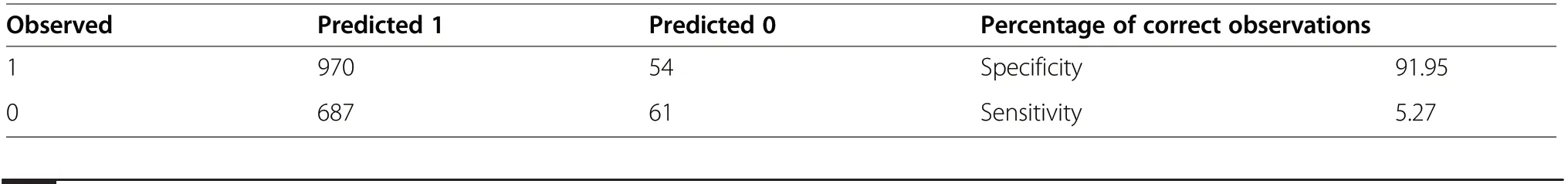
Table 6 Confusion matrix for the model parametrised by the method of searching all the possible combinations using the AIC criterion AIC(OR=3.42,Log OR=1.23), 1 –active Key Areas, 0 –abandoned Key Areas
The process referred to here also applies to sites in which lekking of capercaillies takes place. Many descriptions of leks indicate that a great share of old forest is a typical requirement for the species (Gjerde and Wegge 1989; Storch 1993; Swenson and Angelstam 1993; Saniga 1996, 2003). And yet, several studies from more recent decades document the presence of leks in middle-aged stand classes (Picozzi et al. 1992; Miettinen et al. 2010;Sirkiä et al. 2010; Wegge and Rolstad 2011). Our work suggests that the Augustów Forest’s active leks are predominantly located in stands which are 30 to 100 years old, not in the oldest parts of stands.
The process of leks in mature (>100 year-old) forest being abandoned by capercaillies has been observable in the Augustów Forest over the last few decades, while new places for the leks have been observed in younger stands (50–60 years old) (Zawadzki et al. 1999; Brzeziecki et al. 2012; Zawadzki and Zawadzka 2012;Zawadzka et al. 2018). Similar trend is visible in commercial forests in South-Eastern Norway, where the formation and establishment of new displaying grounds in stands which are less than 50 years old has been described (Rolstad et al. 2007; Wegge and Rolstad 2011).Positive relationships between the proportion of a forest of younger age classes and the densities of capercaillie populations have been also documented in Finland,where studies suggest that a managed, thinned, middleaged forest (30–40 years and more) is suitable and even has a positive effect on population density at the large scale (Miettinen et al. 2008, 2010). According to Sirkiä et al. (2010), a change of preference regarding stand age can be also indirectly connected with the extent of the fragmentation of the habitat and the spatial distribution of particular patches of a mature forest. Small fragments of old stands occurring as ‘islands’ surrounded by forest in the youngest age classes do not represent favourable conditions for the species. The importance of connectivity between habitat patches suitable for the capercaillie is also emphasised by Bollmann et al. (2011) as well as Sirkiä et al. (2012). The preferences of the species for the mature forest as once described seem to be most directly associated with the structure rather than age, as evidenced also by more recent publications (Storch 2001,2007; Watson and Moss 2008; Miettinen et al. 2010).
Researchers had previously confirmed the major importance of bilberry to capercaillies, both as a food plant for adult birds and as places in which the Lepidotera larvae representing food for chicks may develop (Storch 1993; Selas 2000; Gregersen and Gregersen 2008). In the Augustów Forest, the cover of this dwarf shrub was high across the study area, with the effect that this factor was not considered to influence any differentiation between areas still occupied or already abandoned by the birds under study. Indeed, within the analysed areas, the share of the ground covered by bilberry was actually greater in the abandoned Key Areas, perhaps due to the basic fact that the presence of bilberry bushes attracts potentiallydisturbing bilberry-pickers and also ensures a greater intensity of use of certain roads (a variable not taken into account in the optimal model on the basis of the AIC criteria). However, in line with the expectations, Key Areas which were further from roads were those still supporting capercaillies, while the negative influence of human disturbance on the species has been demonstrated with regard to other areas across the range(Summers et al. 2007; Storch 2013; Moss et al. 2014). It is not only forest works that are damaging to the capercaillie; human recreation also has a negative effect(Coppes et al. 2017,2018).
Conclusions
Altogether, changes in forest structure at the local(patch) scale are important, though they are probably not the only reasons for the decline in the numbers of the capercaillie. The grouse representing the subject of our study are shown to occupy the dry and poor,middle-aged, pine-dominated, single-layered forest with a much less-diversified habitat mosaic than in other parts of their lowland range.Our research further reveals that over the last few decades the species has abandoned the oldest stands in the Augustów Forest, with leks having been transferred to younger stands. Our analysis based on the logistic regression model points to the cover shares of the two layers of undergrowth as the most important features discriminating those Key Areas for capercaillies that remain active as lekking grounds or else have already been abandoned. Any development of a greater inclusion of trees and shrubs – stimulated by climate changes or the eutrophication of habitats, or in connection with forest management – will do much to reduce the probability of the occurrence of the capercaillie. The lack of natural disturbances also favours the development of a dense layer of undergrowth. The birds’transfer of leks to younger stands is probably associated not so much with an age preference as with a specific kind of not-too-dense stand structure. In places where under-layers of vegetation are artificially introduced or appear spontaneously under the canopy of older stands,this process changes the forest structure in a manner unfavourable to the capercaillie. Thus, habitat improvement for the species in question requires that the natural development of the understorey (mainly spruce under the conditions in the Augustów Forest) should be held back, or not introduced at all where this would be an element of managerial measures. Also recommended is the maintenance of a more open stand closure than hitherto, one not exceeding 70%. The logistic regression model based on stand variables is well able to point to habitat that is suitable for capercaillies, even if it is less precise in indicating which habitat features are inappropriate for the species. However, it is strongly recommended that the habitats which the study revealed to be suitable should not be transformed with practical measures.
Authors’contributions
SD conducted the conception,design of the work, data collection, statistical analysis, data interpretation, draft of the work. DZ conducted the conception,data collection, draft of the work, significant revision, review of the literature.GZ conducted the data collection and analysis, design of graphics, review of the literature.MS conducted the statistical analysis. BB conducted the conception, design of the work, data collection, review of the literature,significant revision. The authors' read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Author details
1Institute of Forest Sciences, Warsaw University of Life Sciences,Nowoursynowska 159, 02-776 Warszawa, Poland.2Department of Forest Sciences, University of Łódź, Branch in Tomaszów Mazowiecki, Konstytucji 3 Maja 65/67, 97-200 Tomaszów Mazowiecki, Poland.3Institute of Agriculture,Warsaw University of Life Sciences, Nowoursynowska 159, 02-776 Warszawa,Poland.
Received: 11 January 2021 Accepted: 6 June 2021
猜你喜欢
数理天地(高中版)(2022年19期)2022-05-30 10:48:04
郑州大学学报(理学版)(2020年1期)2020-02-08 08:40:00
中学生数理化(高中版.高考数学)(2019年9期)2019-11-27 20:09:58
Chinese Medical Sciences Journal(2019年1期)2019-04-11 09:26:46
铜仁学院学报(2018年4期)2018-06-13 03:21:38
临床医药文献杂志(电子版)(2017年11期)2017-05-17 04:48:46
经济与管理(2016年2期)2016-12-01 04:28:52
新闻传播(2016年2期)2016-07-12 10:52:16
无锡商业职业技术学院学报(2015年1期)2015-12-22 12:35:44
中国流通经济(2015年10期)2015-12-05 04:01:37

- Forest Ecosystems的其它文章
- Distribution of Panama’s narrow-range trees:are there hot-spots?
- Linkage of microbial living communities and residues to soil organic carbon accumulation along a forest restoration gradient in southern China
- Simultaneous optimization of even flow and land and timber value in forest planning:a continuous approach
- Forest disturbances and the attribution derived from yearly Landsat time series over 1990–2020 in the Hengduan Mountains Region of Southwest China
- Nutrient resorption strategies of three oak tree species in response to interannual climate variability
- Topography modulates effects of nitrogen deposition on microbial resource limitation in a nitrogen-saturated subtropical forest