
牛十二指肠贾第虫的分子流行病学研究进展
2021-02-27 04:56蔡伟龙冯耀宇肖立华
畜牧兽医学报 2021年2期
蔡伟龙,李 娜,冯耀宇,肖立华
(华南农业大学,广州 510642)
贾第虫(Giardiaspp.)是常见的肠内寄生原虫,能感染人和多种脊椎动物。贾第虫的生活史存在两个形态不同的阶段,即滋养体和包囊[1]。滋养体为营养繁殖阶段,为贾第虫在宿主小肠内寄生的形态;包囊为传播阶段,在小肠内形成后随粪便排出体外,对外界环境有很强的抵抗力[2-3],主要通过粪-口途径经污染的饮水和食物传播,引起以腹泻、腹胀、营养不良和体重减轻为主要症状的贾第虫病(giardiasis)[4-6]。贾第虫病呈世界性分布,全世界每年有2.5亿~3.0亿人感染贾第虫[4, 7],大多数为无症状感染者[4-5,8]。据统计,发达国家贾第虫病的感染率普遍较低,为0.4%~7.5%;发展中国家的感染率远高于发达国家,为8.0%~30.0%[4]。许多因素与贾第虫感染率存在相关性,如社会经济、卫生条件、地理区域和诊断敏感性[9]。目前,已出现100多 起水源性贾第虫病的暴发[4, 10],绝大部分与饮用污染的水有关,且大多数暴发出现在北美和欧洲[4]。贾第虫的宿主范围非常广泛,能感染多种动物,包括人类、家畜、伴侣动物和野生动物[4, 11]。贾第虫病在我国广泛存在,尽管尚未出现大规模的贾第虫病暴发。我国每年的感染者约占世界的10%[12],不同地区的贾第虫感染率在0.85%~9.46%浮动[13],高感染率人群集中在15岁以下的儿童,其中,感染率最高的是5~10岁的儿童[14-15]。
1 贾第虫分类
1.1 贾第虫虫种分类
贾第虫分为Giardiaduodenalis、Giardiaagilis、Giardiamuris、Giardiapsittaci、Giardiaardreae、Giardiamicroti和Giardiavarani7个虫种,每个虫种都有其特定的宿主范围[5, 16](表1)。十二指肠贾第虫Giardiaduodenalis(又称Giardiaintestinalis或Giardialamblia)能感染人和大多数哺乳动物[4, 17-18],其他虫种主要感染两栖类、爬行类、鸟类和哺乳动物中的啮齿类[17,19]。十二指肠贾第虫的一些基因型(又叫集聚体)可在人和动物之间流行,具有一定的人兽共患意义[20],目前,绝大多数研究都集中于十二指肠贾第虫[16-17]。

表1 已确定的贾第虫虫种和主要宿主
1.2 十二指肠贾第虫集聚体分类
分子生物学研究表明,十二指肠贾第虫可被进一步细分为具有遗传相关性的集聚体[21],而这些集聚体有的来源于特定的宿主[16]。十二指肠贾第虫分为A~H 8种集聚体(assemblage A~H),不同集聚体的宿主范围都不一样[4,24-29](表2)。其中,集聚体A和集聚体B的宿主范围非常广泛,能感染人和大多数哺乳动物,是人兽共患型集聚体;集聚体C和D主要感染犬科动物;集聚体E主要感染有蹄类动物,如牛、羊、猪、马等家畜;集聚体F主要感染猫;集聚体G主要感染啮齿类动物;集聚体H主要感染海豹等海洋动物[5, 23](表2)。

表2 十二指肠贾第虫的集聚体及其宿主范围
2 十二指肠贾第虫的分型工具
目前,常用于十二指肠贾第虫基因分型的遗传位点,包括谷氨酸脱氢酶基因(gdh)、核酸丙糖异构酶基因(tpi)和β-贾第素基因(bg)[4, 17]。通过对这3个靶基因进行巢式PCR扩增,分析PCR产物序列的单核苷酸多态性(SNPs),可将十二指肠贾第虫的分离株分为A~H 8种集聚体[4-5,16]。图1为十二指肠贾第虫的集聚体在3个遗传位点的系统发育树。
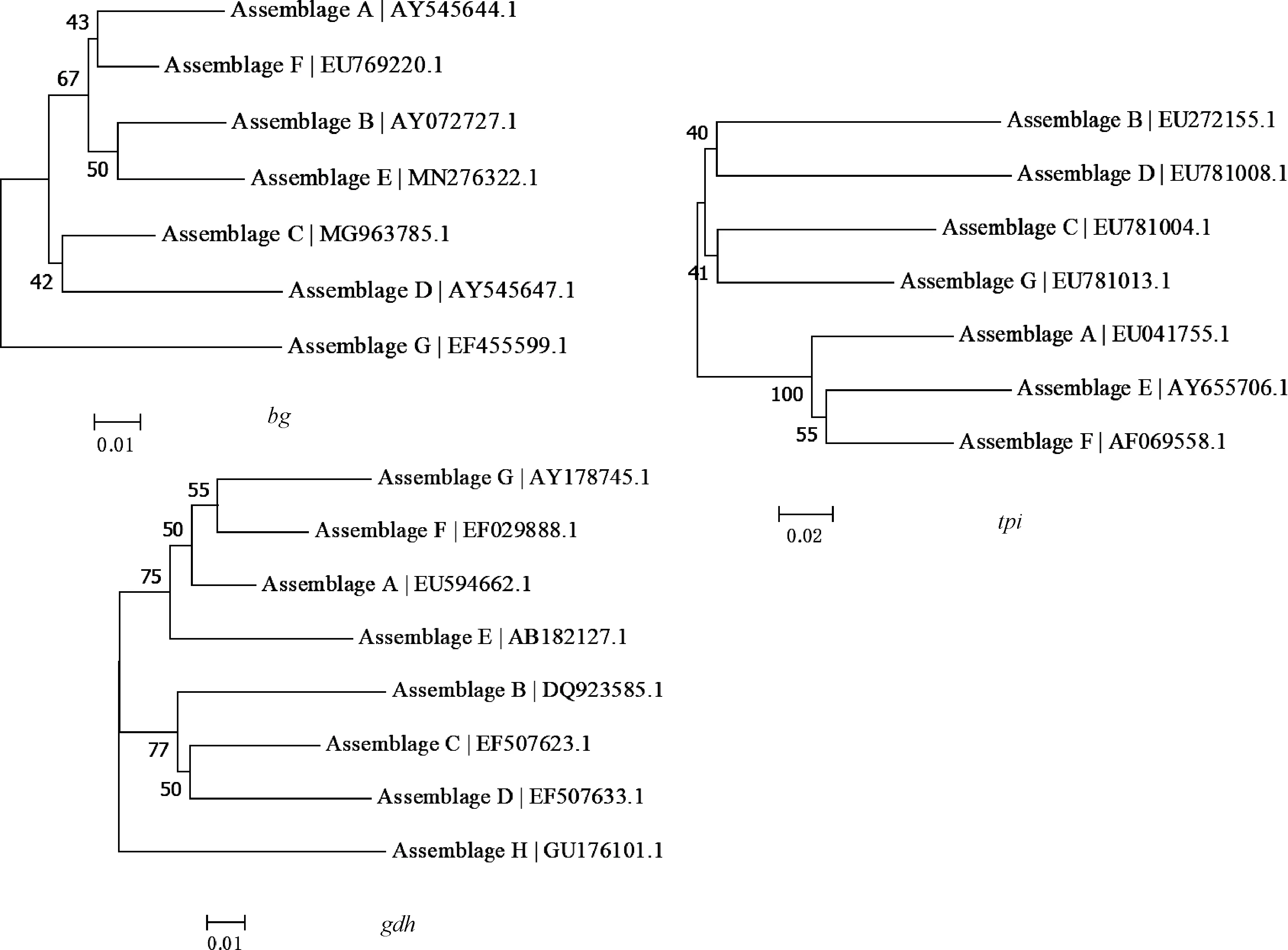
图1 十二指肠贾第虫集聚体在3个遗传位点(bg、tpi、gdh)的系统发育分析(NJ树)
在每种集聚体中,还可以通过SNPs分析,进一步对其进行亚型分型[30]。集聚体A和集聚体B的宿主范围非常广,存在一定的人兽共患风险,所以了解这两种集聚体在宿主内的亚型分布及基因差异可以提供关于亚型与宿主关系、每个亚型的致病潜力、传染源追踪、贾第虫病暴发调查以及传播动力学的信息[4]。在集聚体A中,每个基因位点都有多个亚型,通常称为A1~A6[4,30]。在这几个位点中,A1和A5亚型具有相似的序列,形成了一个与A2~A4及A6形成的第二亚组不同的亚组[4]。一些亚型有明显的宿主适应性,目前发现A1大多在动物中发现,A2大多在人类中发现,而A6几乎只出现在野生反刍动物中[4,31]。集聚体B的亚型比集聚体A的多,但各亚型在3个位点不形成稳定的系统关系[31]。原因之一是集聚体B的亚型存在等位基因序列杂合性[32],难以用基于PCR产物的序列分析对集聚体B进行亚型分型[31-32]。集聚体E也有许多亚型,但在其各种位点的核苷酸序列中没有具宿主特异性的亚集聚体(sub-assemblage)[30]。
对不同的遗传位点分析时会获得不完全一致的基因分型结果[24,33]。多位点基因型(multilocus genotype,MLG)是一种综合分析十二指肠贾第虫3个遗传位点(tpi、bg、gdh)后构建的用来更好地表征遗传性质的基因型[24,34]。基于这3个位点的多位点分型工具(multilocus genotyping tools)越来越多地用于十二指肠贾第虫的基因分型和集聚体A的亚型分型[4],并且已经运用到十二指肠贾第虫的群体遗传学研究中[34-36]。现在,利用多位点分型工具可将集聚体A的亚型归到3个亚集聚体:AⅠ、AⅡ和AⅢ,以此为基础建立的系统发育树可清晰地描述集聚体A各亚型和亚集聚体之间的亲缘关系[4]。此外,集聚体B的序列杂合性也使得多位点分型工具无法用来定义其亚集聚体[4, 24]。
3 我国牛十二指肠贾第虫的流行情况
牛十二指肠贾第虫感染在全世界广泛分布。在多项研究中,十二指肠贾第虫的阳性率在1.09%~74.2%浮动,且奶牛的阳性率明显高于肉牛[37-40]。在集聚体的分布上,集聚体E是感染牛的主要基因型,其次是集聚体A,集聚体B仅出现在少数研究中[4, 41]。其他集聚体仅有少数不确定的报道[4, 42]。
目前,我国牛十二指肠贾第虫的流行情况与世界上其他国家的情况大致相同(表3)。在广东的一项调查中,十二指肠贾第虫的阳性率高达74.2%。其次是上海,阳性率为60.1%,而西北地区(甘肃、宁夏、陕西、青海等)和东北地区(黑龙江、吉林和辽宁)的阳性率较低(均低于20%)。在河北、天津和北京的研究中,十二指肠贾第虫阳性率均低于5%,分别是4.7%和1.1%(河北和天津的数据来自同一研究)。同一地区不同研究所报道的十二指肠贾第虫阳性率差异明显,广东的两项研究中,阳性率分别为74.2%和2.2%,而四川的研究则分别为41.2%、9.4%和1.2%(表3)。这些差异可能主要是由采样动物年龄、健康状况及检测方法不同引起的。
在集聚体分布方面,集聚体E的比例占绝大多数(94.5%),而集聚体A和B呈散发分布,占比分别是2.7%和1.5%。值得注意的是,集聚体A的分布没有明显的地域差异;而集聚体B的出现较为集中,只在青海、宁夏、上海和黑龙江有发现,且检出的数量很少(表3)。在混合感染方面,存在集聚体A和E的混合感染,且几乎都集中在东部地区,在西部地区未有发现(表3)。

表3 我国牛十二指肠贾第虫流行情况和集聚体类型
4 牛十二指肠贾第虫的公共卫生意义
高感染率、人兽共患威胁及潜在的暴发可能使得十二指肠贾第虫病在全世界范围内都有着重要的公共卫生意义。研究十二指肠贾第虫分子流行病学的一个重要方面是深入了解其不同集聚体的宿主范围及跨物种传播的可能性[4]。如前文所述,感染人的十二指肠贾第虫的基因型绝大多数是集聚体A和B,这两种集聚体具有广泛的宿主范围和潜在的人兽共患威胁[76]。此外,在人十二指肠贾第虫的基因型研究中,集聚体C、D和F有一些不确定的报道[77-81],巴西、埃及和澳大利亚也曾报道集聚体E的出现[27,82-83]。然而,这几种集聚体在人中的分子流行病学数据非常少,这些集聚体的人兽共患潜力在逐步得到认识。
在分子流行病学的层面上,对十二指肠贾第虫的基因型研究得最多的是集聚体A。利用多位点分型工具可以将集聚体A分为3类亚集聚体:AⅠ、AⅡ和AⅢ,且这些亚集聚体存在一定的宿主差异[4, 17]。目前发现,AⅠ和AⅡ是人和动物中常见的亚集聚体[17],其中AⅠ主要感染哺乳动物,AⅡ在人中更为常见,AⅢ主要感染野生动物和反刍动物[24,84]。在牛十二指肠贾第虫的研究中,AⅠ的比例明显高于AⅡ[63]。除了集聚体A外,集聚体B也在牛中发现,但其分布有限[17]。这些因素,加上集聚体E感染率占绝对优势,使得牛十二指肠贾第虫的公共卫生意义可能有限[4]。
集聚体A的实际感染率可能比过去认为的要高[4]。在一些牛十二指肠贾第虫病的调查中,集聚体A的感染率就出现了预料之外的结果。在澳大利亚的一项研究中,集聚体A在贾第虫阳性样品中的占比为43%[85]。此外,美国的一项牛贾第虫纵向调查发现[86],小于18周龄的犊牛仅发现集聚体E的感染,在7周龄时集聚体E的累计感染率甚至可达到100%;而当牛生长到18周龄后,集聚体A开始出现,并且在15月龄时的累计感染率高达70%。有研究认为,十二指肠贾第虫在宿主体内发育的过程中,集聚体A和E可能存在竞争关系,并且两者的分布可能与牛的年龄有关[4,87]。尽管集聚体E仍然是目前感染牛最主要的基因型,然而,随着集聚体A越来越多被发现,集聚体A在牛中的分布可能更广泛[4, 17]。同时,近年来,国外已经有多起人感染集聚体E的报道,表明集聚体E可能具有一定的跨宿主传播能力[27],其人兽共患潜力需要得到更多的认识。
5 小 结
贾第虫是一种全球范围内分布的重要肠道寄生性原虫,这种带鞭毛的原生动物会引起自限性的贾第鞭毛虫病[88]。该病的典型症状表现为腹泻、腹部绞痛、腹胀、体重减轻和吸收不良,但以目前的流行病学数据来看,无症状感染的病例居多。贾第虫可在人和多种哺乳动物体内寄生,形成包囊随宿主粪便排出,污染食物、水源和界面。包囊的环境抗性很强,能在环境中长期生存[10,89-90],使贾第虫病能在宿主之间持续传播,增加了其食源性和水源性传播的风险[10, 89, 91]。但是,很多国家对贾第虫病的病原检测和水质监测并未到位,其实际上的食源性感染率可能更高[91]。全球范围内已经出现多起贾第虫病的暴发,但贾第虫病的公共卫生意义还未得到广泛的关注,贾第虫病的人兽共患风险也未得到充分的认识[91]。
牛作为重要的经济动物,与人类社会的关系非常密切。当前,牛养殖业的集约化规模越来越大,牛作为人兽共患寄生虫病的传播来源对人的威胁也越来越大。感染了十二指肠贾第虫的牛可向外界环境排出大量包囊(1头牛1 d可排出7.6×106个包囊)[92-93],而这些包囊可在环境中广泛传播。另一方面,犊牛感染十二指肠贾第虫后可出现腹泻、营养不良等症状,使其发育迟缓和生产力受限,影响着牛养殖业的经济效益[40]。传统上,贾第虫病的诊断方法是使用显微镜检查粪便中的滋养体和包囊,但耗时长,且可能出现假阴性的结果[94]。免疫学诊断(ELISA、IFA和免疫色谱检测等)的敏感性和特异性较高,现在常用于贾第虫病的检测[95]。巢式PCR技术是现今应用最广泛的贾第虫检测技术,其敏感性和特异性分别可以高达88.9%和82.9%[96]。
分子分型工具的发展已经大大改变了人们对贾第虫人兽共患潜力的理解[97],使人们更深入地了解人兽共患型的集聚体所表现出的遗传特征,并且逐渐认识其他集聚体潜在的跨宿主传播能力和人兽共患风险[4, 98]。当前,牛十二指肠贾第虫的分子流行病学研究还不够全面和深入[5, 19]。分析十二指肠贾第虫的集聚体、亚集聚体和亚型在人和动物之间的流行特点,深入研究十二指肠贾第虫分子流行病学规律,是目前研究和防控牛十二指肠贾第虫病重点所在[4],也是未来牛十二指肠贾第虫病研究的发展方向。
猜你喜欢
科学(2020年3期)2020-11-26
科学(2020年3期)2020-11-26
当代水产(2020年3期)2020-06-15
中国人兽共患病学报(2019年10期)2019-11-06
天津科技大学学报(2019年5期)2019-10-23
中国兽医杂志(2019年2期)2019-06-25
中国兽医杂志(2019年2期)2019-06-25
中国人兽共患病学报(2019年11期)2019-01-10
作文·初中版(2017年4期)2017-04-18
小星星·阅读100分(高年级)(2015年11期)2015-11-28
