
滨海湿地生态系统土壤微生物及其影响因素研究综述
2021-02-25 06:56:20解雪峰蒋国俊孙晓敏濮励杰
生态学报 2021年1期
解雪峰,项 琦,吴 涛,蒋国俊,孙晓敏,朱 明,濮励杰,*
1 浙江师范大学地理与环境科学学院, 金华 321004
2 南京大学地理与海洋科学学院, 南京 210023
3 自然资源部海岸带开发与保护重点实验室, 南京 210023
滨海湿地作为介于陆地生态系统和海洋生态系统之间的过渡区域,是指受海陆交互作用下经常被静止或流动的水体所浸淹的沿海低地,潮间带滩地及低潮时水深不超过6 m的浅水水域,主要包括滩涂、盐沼、浅海水域等[1]。受海陆交互作用干扰,滨海湿地生态系统已成为全球环境变化的敏感区和生态系统的脆弱区,具有显著的区域特性[2]。滨海湿地土壤作为滨海湿地生态系统的重要组成部分,是滨海湿地生物的基质和载体,也是滨海湿地生物地球化学循环的中介[3]。滨海湿地常年处于滞水或周期性淹水环境下,透气透水性差,氧化还原电位低,表现出强烈的还原环境;同时,滨海湿地土壤微生物在厌氧环境下以嫌气性细菌为主,对动植物残体分解缓慢,表现出有机物质的累积[4]。
土壤微生物是土壤中存活的原核微生物和真核微生物的统称,主要包括细菌、真菌、放线菌、藻类等[5]。土壤微生物是土壤生态系统中最活跃的组成部分,参与土壤有机质矿化分解、腐殖质合成、土壤养分的循环和转化,对土壤团聚体形成与稳定具有显著作用,是实现土壤功能的重要保障,也是衡量土壤生态系统稳定性的一个重要指标[6-7]。滨海湿地土壤微生物还具有独特的功能和基因资源,在污染物净化、生态系统维稳、生态系统健康等方面起着重要的作用,比如其生物电化学作用在土壤脱盐过程中发挥着重要作用[8]。滨海湿地土壤微生物对生存环境极度敏感,并能够迅速做出响应[9]。近年来,对滨海湿地生态系统土壤微生物的研究逐步扩展,该领域的相关研究已成为研究热点[10-11]。但与森林、农田、内陆湿地等生态系统相比,我国滨海湿地生态系统土壤微生物生态学研究起步较晚,且主要集中在不同滨海湿地类型土壤微生物活性的高低和土壤微生物与环境因子之间的简单关系,而对土壤微生物功能的作用机理,以及微生物对环境因子变化的响应机制等问题研究不足[12-15]。
本文系统梳理了国内外对滨海湿地土壤微生物及其影响因素等相关研究,分析和总结了不同环境因素对滨海湿地土壤微生物的影响,以期为更好地保护和修复滨海湿地生态系统提供一定的理论参考。
1 滨海湿地土壤微生物特征
1.1 滨海湿地土壤微生物数量
滨海湿地频繁地受到咸淡水交互作用的影响,加上地质环境条件的梯度变化和人类活动的影响,共同造就了滨海湿地中好氧微生物和厌氧微生物共存于土壤和沉积物中,即具有与陆相和海相因素都相关的特殊的微生物群落,且两种微生物处于互补状态[16-17]。由于滨海湿地土壤具有较高的含盐量及pH值,滨海湿地土壤微生物数量较其他类型的湿地生态系统低,但更耐盐碱性[18]。滨海湿地中细菌数量占绝对优势,其次是放线菌和真菌,随着土壤盐碱化程度的加重,土壤细菌、真菌和放线菌数量及微生物总数呈现显著降低的趋势(表1)[19]。滨海湿地土壤微生物数量还存在明显的时空分异规律。在海陆梯度方向上,受潮汐作用的影响,土壤微生物数量自潮下带向潮上带逐渐递增[24]。土壤微生物数量在土壤剖面上存在显著的垂向分异,土壤微生物大多活跃在土壤表层,随着土壤深度的增加,其丰度逐渐降低[25]。受温度的季节变化和海洋有机物的季节性输入等因素影响,滨海湿地土壤微生物数量也呈现出明显的季节变化特征[21]。一般来说,夏秋季节微生物数量高于冬春季节,这是因为夏秋季节较适宜的温度、湿度,枯落物分解以及新生植被代谢带来的大量营养输入等因素可能共同增加了微生物的数量;相反的,冬春季节水热条件较差,植被生长停滞,凋落物减少,土壤返盐等因素共同导致了微生物数量的降低[24]。
1.2 滨海湿地土壤微生物群落组成及多样性
滨海湿地中土壤微生物群落的丰富度较高,且与植物根系密切相关[26]。土壤细菌群落是滨海湿地土壤微生物组成成分最丰富的微生物群落,也是滨海生态系统地球化学循环的驱动力,广泛地参与分解和转化有机物质[27]。研究表明,滨海湿地土壤中的细菌群落主要由变形菌门(Proteobacteria)和拟杆菌门(Bacteroidetes)组成[28]。此外,酸杆菌门(Acidobacteria)、绿弯菌门(Chloroflexi)、浮霉菌门(Planctomycetes)和放线菌门(Actinobacteria)也在芦苇、碱蓬、红树林、互花米草等滨海湿地土壤中大量存在[28-29]。变形菌门的相对丰度在滨海湿地土壤中所占的比例约为27%—55%,主要由γ-变形菌纲(γ-proteobacteria)、δ-变形菌纲(δ-proteobacteria)和α-变形菌纲(α-proteobacteria)组成,包含较多的固氮细菌,能够明显提升湿地养分循环[29-30]。滨海湿地土壤中拟杆菌门主要包括拟杆菌纲(Bacteroidetes)和黄杆菌纲(Flavobacteriia),其相对丰度约为2%—25%,包含多种营养型细菌,可以广泛生存于滨海湿地环境中[30-31]。古菌所具有的嗜盐碱、寡营养特性使得它们高盐碱且缺氧的滨海湿地广泛分布。滨海湿地中的古菌群落主要包括深古菌门(Bathyarchaeota)、泉古菌门(Crenarchaeota)和广古菌门(Euryarchaeota),其中海洋古菌群I(Marine group I)、热原体纲(Thermoplasmata)和甲烷杆菌纲(Methanomicrobia)是优势类群[31-32]。真菌作为滨海湿地土壤微生物区系的重要组成部分,参与滨海湿地土壤的腐殖化和矿质化过程,将湿地中的有机物逐渐降解转化[33]。子囊菌门(Ascomycota)和担子菌门(Basidiomycota)是滨海湿地真菌群落的主要优势类群,二者相对丰度约占真菌群落的85%以上(表2)[30,34-35]。
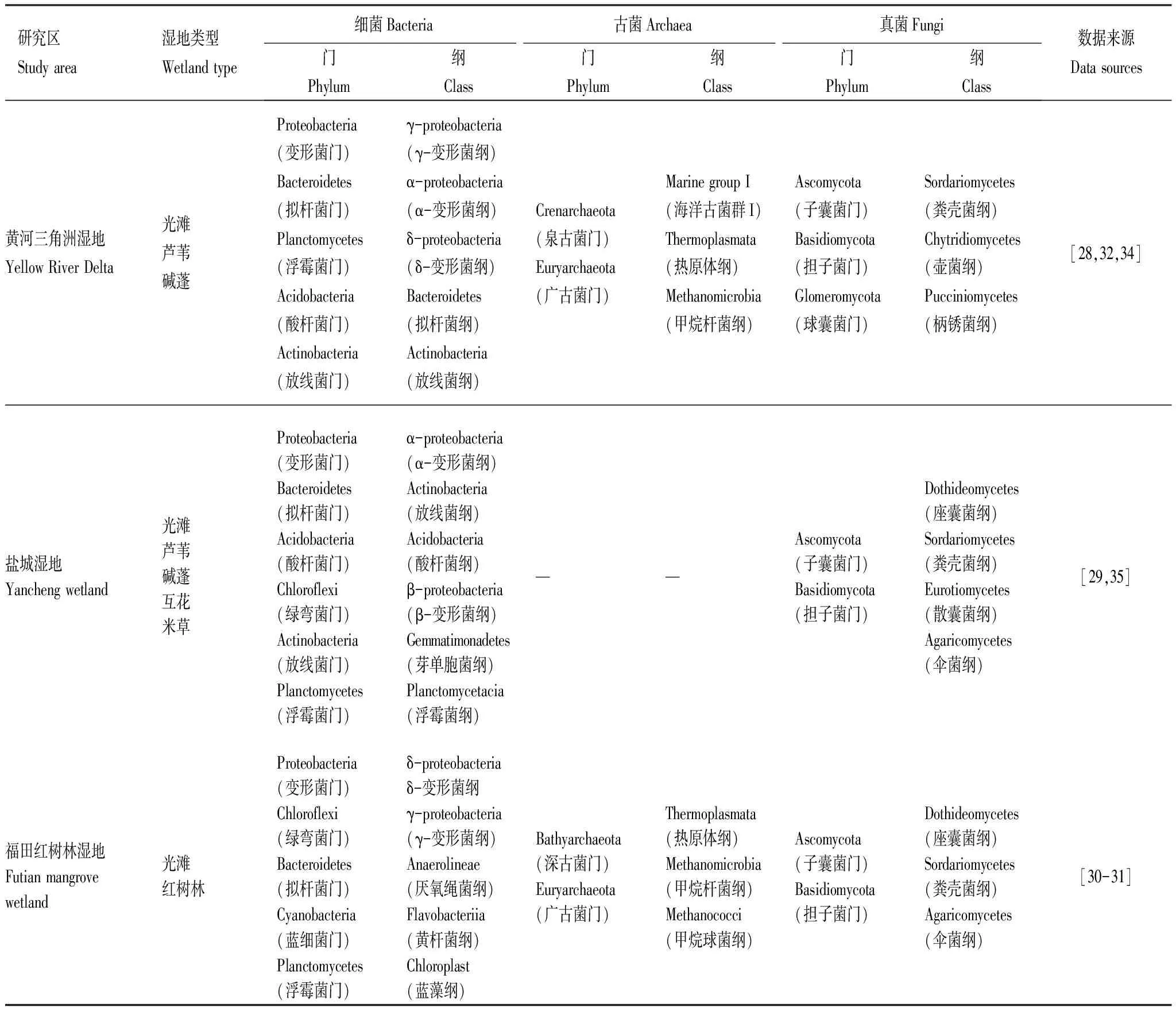
表2 不同滨海湿地类型土壤微生物群落组成
滨海湿地土壤微生物群落多样性因地理位置、植被类型和季节变化呈现出一定的差异性(表3)。从海陆梯度方向上,受潮汐作用影响,土壤微生物群落多样性随潮滩的演替逐渐递增[29]。在黄河三角洲,碱蓬、芦苇群落的土壤微生物群落多样性及丰富度显著高于光滩,且细菌群落的多样性随深度增加逐渐下降, 而古菌群落多样性随深度增加则呈上升趋势[37]。在深圳福田红树林湿地,细菌、古细菌和真菌的物种丰富度和香农多样性均显著高于光滩,且夏季显著高于冬季[38]。相似的,在江苏盐城和崇明东滩,海三棱藨草群落、芦苇群落的土壤微生物群落多样性及丰富度同样显著高于光滩,且夏秋季显著高于春季[10,29]。
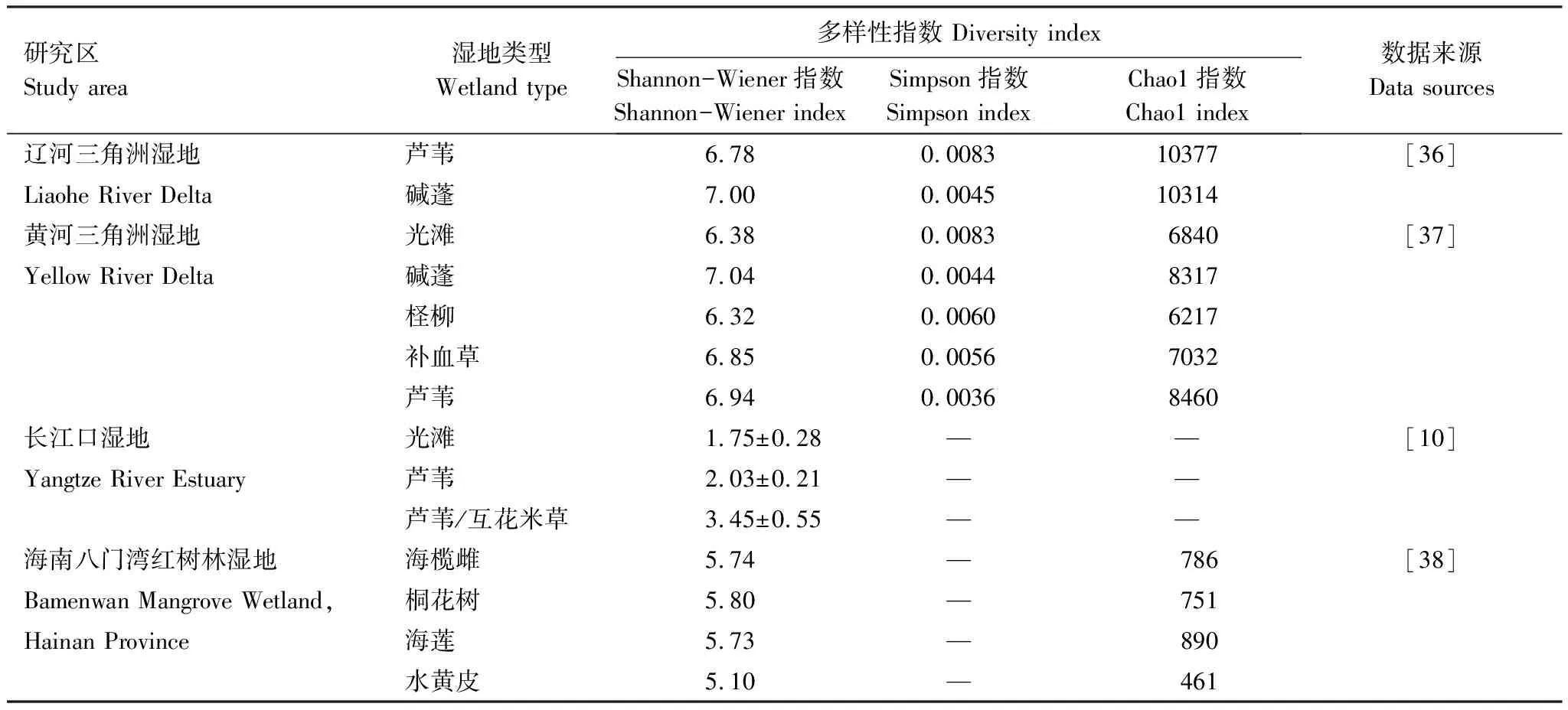
表3 不同滨海湿地类型土壤微生物群落多样性
2 自然环境因子对滨海湿地土壤微生物的影响
2.1 土壤理化性质
滨海湿地生态系统土壤微生物群落组成受土壤孔隙度、土壤水热条件、土壤pH和土壤盐分等诸多理化性质的影响[10,28,30]。土壤孔隙度影响土壤微生物种群及其活动,其大小决定了微生物在土壤中的活性及多样性的高低。由于大部分微生物为好养型微生物,良好的通气状况有利于微生物的生存和活动,而滨海湿地由于容重较高、孔隙较小,使得土壤通气性较差,对土壤微生物活动存在较大影响,只有嫌气性和兼性微生物能进行正常的活动[37]。
滨海湿地生态系统中,土壤的水热条件影响着土壤的通气性能、呼吸强度和盐碱特征,进而影响土壤微生物群落结构和多样性特征。研究发现,在一定范围内土壤含水量的升高可增加土壤微生物量并促进土壤呼吸作用强度,而过量的水分条件会使湿地土壤通透性降低,导致土壤微生物的丰富度及生物量下降[39]。对辽河口芦苇湿地的研究发现,受潮汐作用影响,湿地在淹水状态下土壤通气性降低,呼吸作用减弱,盐碱程度升高,使得土壤微生物群落的生物量和丰富度显著下降[40]。温度对土壤微生物的活性和群落结构的影响目前仍无统一的结论。多数研究认为,当土壤温度在0—35 ℃范围内时,微生物的数量和多样性与温度的变化呈正相关关系,但当土壤温度大于40 ℃时,微生物的活动会受到显著抑制[41]。李艳等在崇明东滩芦苇湿地进行长期模拟增温,结果发现增温下芦苇湿地土壤细菌和真菌数量均显著上升,微生物多样性也显著提升[42]。由于土壤温度具有清晰的季节性特征,因此滨海湿地土壤微生物的丰度和活性在季节上也存在差异,大致表现为从春季开始逐步上升,在夏季到达峰值,随后又逐渐回落[25]。
土壤pH被认为是影响土壤微生物种类和结构的主要因素,对土壤微生物在生物群落和区域尺度上的多样性和群落组成有重要影响[43]。一般情况下,酸性条件有益于真菌的活动,中性至微碱性的土壤环境益于土壤细菌和放线菌生存,真菌、细菌和放线菌生长的土壤pH值最宜区间分别为4.0—6.0、6.5—7.5、7.0—8.0[44]。在滨海湿地生态系统中,由于土壤呈碱性且pH波动范围较小,加上滨海湿地土壤的透气透水性差,而真菌作为典型的好氧菌在这种环境下活性较低,使得滨海湿地土壤微生物中细菌和放线菌数量占主导地位[19]。大量研究表明土壤微生物数量及多样性与土壤pH大致成反比,这主要是因为滨海湿地生态系统中较高的土壤pH改变了细胞中的生物大分子的电荷,从而抑制细胞的活性;同时土壤pH抑制了生物体所处环境中营养物质的可利用性并增加了有害物质的毒性[26,30]。然而,堀越芽氏孢杆菌(Bacillus horikoshii)和列城芽孢杆菌(Bacillus lehensis)等由于具有较强的耐强碱特性而得以较好的生长[45]。
土壤盐分的改变引发土壤渗透压的波动和地表覆被变化,通过影响微生物生物量和活性进一步影响滨海湿地微生物的组成和功能,主要表现为滨海湿地土壤微生物群落结构沿盐度梯度显著变化[46]。诸多研究表明土壤盐分对大多数土壤微生物的数量、组成和多样性有显著的抑制作用,在低盐环境中土壤微生物的数量和类型较多,当盐度过高时,土壤微生物活性显著下降[32,46]。以硫酸还原菌(Sulfate-Reducing Bacteria)为例,过高的盐度会抑制其生长,只有在盐度低于2.28%时才可以正常生长[47]。相似的,盐分被认为是影响黄河三角洲植被演替系列丛枝菌根真菌群落组成的最主要因素[48]。当然也存在一些嗜盐微生物,如产微球茎菌属(Microbulbifer)只有在土壤盐分含量达到2.69%时才会出现[45]。
2.2 植被状况
滨海湿地土壤微生物的数量、群落组成及多样性均受湿地植被状况,如植物类型、覆盖率、植被演替阶段、生物量和生物多样性等因素的影响。滨海湿地植被通过提供氧气、凋落物和根系分泌物,创建更适合土壤微生物繁衍的土壤环境,从而影响土壤微生物群落[24]。如黄河三角洲湿地植被覆盖区域(芦苇、碱蓬、柽柳)的土壤微生物数量和多样性均显著高于光滩[14]。随着滨海湿地植被演替的进行,凋落物的含量和组成也随之发生了较大改变,进而使土壤微生物在群落结构和多样性等方面产生不同程度的变化[28]。在互花米草和红树林混交区,土壤微生物活性和多样性均高于单一植被群落,即存在边缘效应,这可能是由于植被混交区具有较好的缓存能力和更强的抵抗力所致[49]。
滨海湿地土壤微生物在分布上还存在根际效应,即根际微生物在多样性、丰富度以及代谢活性都远高于非根际微生物。植物根系含有特殊的微生物种群,它们生活在共生环境中,彼此之间以及与根系之间存在复杂的交互作用[50]。发达的植物根系相互缠绕,给土壤微生物搭建出一个良好的生存场所,同时根系分泌物又为其生存和大量繁殖提供了充足的营养物质,但由于不同植物类型所产生的根系分泌物也有所差别,从而有选择性地影响微生物的类型和丰度及其产生效应的大小,促进特定根际微生物群落的建立[51]。研究发现,在红树林湿地中,与非根际沉积物相比,根际沉积物中细菌丰度略低,而真菌丰度显著高于非根际沉积物[52]。在印度Gujarat滨海湿地,潮间带盐生植被的根际沉积物中细菌、真菌和放线菌PLFAs生物标记物的丰度明显高于非根际沉积物[26]。
2.3 水文因素
潮汐作用引起的干湿交替和咸淡交替等水文过程是滨海湿地所经历的最频繁的自然过程,使湿地土壤经历了干旱-再湿润-淹水-湿润的变化过程[53]。周期性的涨潮、落潮使滨海湿地土壤处于干湿交替状态,对滨海湿地土壤理化性质造成极大影响(土壤水分、土壤pH、土壤盐分等),进而对滨海湿地土壤微生物的群落组成、分布及其活性造成影响[54]。研究表明,滨海湿地在小潮期间,水文条件主要是地下水位的变化,而大潮期间,潮汐水流的变化构成了水文条件的主体[55]。由于滨海湿地地下水位较浅且为咸水,地下水位的上升及土壤含水量的增加会使溶解氧浓度下降,从而降低湿地微生物种类与活性[56]。Unger等研究也发现,淹水时间过长在降低土壤微生物生物量的同时,也会干扰滨海湿地土壤微生物群落组成[57]。此外,周期性潮汐通过横向的输运作用把海洋和河流带来的大量营养盐输入滨海湿地,从而使滨海湿地生态系统的生产力维持在较高的水准,并影响滨海湿地的植被分带,进而通过影响植物的生长及湿地植被结构来影响滨海湿地土壤微生物的群落结构和活性[54]。
滨海湿地水文条件所驱动的土壤氧化还原电位(Eh)对土壤微生物的影响也十分显著。一般来说,土壤含水率越低,透气性越好,土壤Eh越高,好氧微生物的活性随之增加;而在淹水环境下,土壤Eh较低,土壤微生物活性相对较低[15]。研究表明,细菌、真菌和放线菌的数量均与土壤Eh呈显著正相关。好氧微生物如硝化细菌、反硝化细菌、放线菌和真菌等在Eh值+100 mV以上均可生长,以+300—+400 mV最适宜生长;兼性厌氧微生物在Eh值+100 mV以上时进行好氧呼吸,在Eh值+100 mV以下时进行无氧呼吸和发酵作用;厌氧微生物在Eh值低于-100 mV以下时能较好生长[58]。以硫酸还原菌和产甲烷菌为例,作为典型的厌氧微生物,硫酸还原菌在Eh值在-200—-250 mV时开始进行硫呼吸;甲烷产生菌只有在Eh值低于-330 mV才能生长,并分解底泥中的有机质[59]。
2.4 生物入侵
生物入侵通过影响生态系统结构、过程和功能来威胁生物多样性。多项研究表明,入侵植物会干扰侵入地土壤的温度、湿度、有机质含量等理化性质,对植物群落的物种组成和结构造成不同程度的影响,从而改变凋落物和根系的数量和成分,引发原生土壤微生物群落结构的变动[24]。滨海湿地复杂的动态异质环境和强烈的人类活动则为外来物种的入侵及扩张提供了广阔的生态空间,成为生物入侵的热点区域[22,35]。以互花米草为例,互花米草入侵原生湿地后大量繁殖,通过凋落物和根系分泌物等使湿地土壤理化性质发生改变,破坏湿地土壤的微生态,弱化滨海湿地土壤生态系统的群落结构,使得侵入地微生物多样性大大下降[60]。互花米草入侵红树林湿地后,显著提高了土壤中革兰氏阳性细菌、真菌的PLFAs生物标记物丰度,降低了革兰氏阴性细菌、放线菌的PLFAs生物标记物丰度;同时,互花米草入侵抑制了红树林根际土壤微生物的代谢活性,进而引起微生物群落结构发生改变[61]。互花米草入侵碱蓬、芦苇和海三棱藨草等本地植物时,细菌和真菌的丰富度和多样性均显著增加,但随着入侵年限的推移呈现出先上升后下降的趋势[35]。互花米草入侵还影响微生物功能群落的组成。互花米草入侵后通过影响土壤中的碳和氮来驱动微生物群落,促使根际固氮微生物的数量和丰富度明显提高,如互花米草群落中氨化菌功能群、固氮菌功能群和硝化菌功能群中微生物数量显著高于光滩[62]。互花米草入侵中国东部沿海芦苇、碱蓬湿地时,极大地增加了硫酸还原菌群落的丰富度和多样性,改变了硫酸还原菌群落的结构,如互花米草的脱硫杆菌科相对丰度较本地种提高了13%[63]。
2.5 全球变化
20世纪以来,以气候变暖为主要标志的全球变化深刻地影响着地球生态环境尤其是滨海湿地生态系统,主要包括以CO2为代表的温室气体浓度增加、气候变暖和氮沉降增加等[64]。大气CO2浓度升高对植物产生巨大影响的同时,还间接的影响土壤微生物。就滨海湿地而言,CO2浓度的增加以及全球变暖所带来的气温、降水等变化对土壤微生物的群落结构、丰度和多样性均会造成不同程度的影响[65]。研究表明,大气中CO2浓度升高会提升植物光合速率,增加根系生物量和分泌物,从而使土壤的物质组分和土壤理化性质发生改变,间接影响土壤微生物生态系统,改变土壤微生物生物量及群体组成[66]。此外,二氧化碳浓度的上升也会提高植物的代谢作用,从而使土壤微生物更为活跃;但也有学者提出不同观点,认为大气中CO2浓度的上升也可能会抑制凋落物的分解,进而导致滨海湿地土壤微生物活性的降低[65]。
全球变暖可能会影响滨海湿地生态系统固有的生物地球化学平衡,温度的急剧升高被认为是影响滨海湿地土壤生态系统功能的主要因素[67]。一般来说,气候暖化对滨海湿地土壤表层温度的影响更为明显,从而对生活在土表的微生物产生干扰,而对深层土壤中的微生物影响不大[68]。全球变暖所引起的极端降水事件趋多也会对滨海湿地土壤微生物产生影响。由于滨海湿地地下水位较浅,强降水事件的发生常会造成短时湿地土壤饱和甚至淹水,抑制土壤微生物活性,不利于微生物的生存和繁殖;反之,干旱事件发生时,土壤中的盐分随毛管水的上升而在土壤表层富集,引起盐渍化现象的发生,造成大部分微生物生存环境的丧失,不利于多样性的发展[69]。
此外,大气氮沉降也是全球变化的重要组成部分,长期增加的氮沉降会使滨海湿地土壤生态系统的生产力和稳定性经受巨大考验[70]。研究表明,氮沉降会使滨海湿地土壤中氮素的浓度和形态发生变化,改变土壤中有效元素组成,加速凋落物的分解,影响土壤营养状况,改变土壤理化性质,最终影响微生物群落组成、活性以及物种相对丰度[71]。微生物对于氮浓度有一定的耐受范围,少量的氮沉降会增加微生物多样性和丰度,活性也有所增强,而大量的氮沉降会导致土壤矿质氮的急剧增加,使土壤酸性略有提高,降低真菌的多样性,从而改变土壤微生物群落的组成和相关功能[72]。
3 人类活动对滨海湿地土壤微生物的影响
3.1 湿地开垦
随着人口的日益增长和社会经济的快速增加,沿海地区的人地矛盾日渐突出,为适应经济发展的需要,大面积的滨海湿地被围垦后用于农业生产、城市建设和工业发展[73]。高强度的滩涂围垦活动必然引致滨海湿地生态系统的结构与功能发生重大变化,甚至是发生剧烈逆转,极大地改变滨海湿地土壤微生物的群落结构及功能多样性[74]。由于围垦后的滨海湿地不再遭受潮汐作用的干扰,湿地厌氧环境相对减弱,土壤微生物群落结构及其代谢活性也会随之变化;围垦后湿地的水盐运移情况也会有所不同,从而间接扰动滨海湿地土壤微生物[75-76]。此外,湿地围垦过程中堤坝的兴建也会阻碍滨海湿地与海洋之间的物质传输和能量流动,严重影响了滨海湿地原本的横向物质通量[77]。研究表明随着围垦年限的增加,围垦区土壤中的细菌、真菌和放线菌数量均显著增加,微生物多样性也随之发生较大改变,主要表现为数量占比最大的微生物种群由细菌变为真菌[17]。
滨海湿地围垦过程中,由于耕作方式和化肥农药的施用,滨海湿地土壤微生物的群落结构、多样性及其活性都会遭到影响。长期的耕作会改变土壤中矿物质、有机质等多种组分的含量和分布,改善土壤结构,有利于土壤微生物多样性的增加,间接影响了微生物的代谢活性及群落组成[78]。滩涂湿地围垦后施用化肥或有机肥均可以迅速改变土壤基底营养状况,继而改变土壤细菌和真菌群落结构,表现为细菌群落中放线菌门相对丰度的下降和酸杆菌门相对丰度的上升,以及真菌群落中子囊菌门相对丰度的下降和担子菌门相对丰度的上升[79]。研究发现,适量施肥能起到促进微生物繁殖的作用,但过度施肥反而会造成养分过量输入,抑制微生物的生长[80]。此外,有机肥与化肥混合使用的效果要比单独使用一种肥料的效果更为明显,且中高量的有机肥可以显著提高微生物群落的功能多样性[81]。农药的种类和施用剂量对土壤微生物活性、群落结构及丰度的影响具有明显差异[82]。以除草剂为例,除草剂对微生物群落的影响主要取决于除草剂浓度和土壤类型[83]。需要注意的是,农药的作用效果随着微生物的代谢活动会逐渐减弱甚至消失,因此在常规剂量下长期使用农药不会对土壤微生物造成太大影响,由此部分研究认为农药的合理使用对土壤微生物群落的影响是较小且较短暂的[84-85]。
3.2 石油污染
滨海湿地石油开采过程中产生的石油污染会对植物、土壤动物和微生物产生毒害作用,进而引起土壤微生物群落结构和多样性发生变化[86]。研究表明,受污染滨海湿地生态系统中石油组分的浓度对微生物多样性、丰度和群落结构有较大的影响,随着石油污染物浓度的上升,土壤微生物群落的均匀度、多样性和丰度均逐渐下降,出现了Gulosibacter、Halomonas(盐单胞菌属)、Petrobacter、Methylocystis、Pseudoalteromonas等变形菌纲的优势菌群[87-88]。也有研究表明石油污染会使红树林湿地细菌群落的丰度和多样性显著下降,但对古菌群落的影响并不明显[89]。由于石油污染滨海湿地土壤后,具有降解功能的微生物类群逐渐富集,功能微生物(如石油降解菌群)所占的比例升高,土壤微生物活性反而有所增加,但随着土壤中污染物逐渐被分解,该类微生物的数量也随之下降[90]。此外,石油污染物会使滨海湿地土壤自身理化性质产生变化,从而间接对微生物活性及其群落结构产生影响[86]。由于石油的密度小、黏着力较强,易破坏土壤结构,造成土壤孔隙堵塞,使土壤透水性降低,从而影响土壤的通透性;同时,石油污染物会引起土壤电导率和总有机碳升高,土壤含水量、总氮和总磷下降,改变土壤结构和有机质组成和结构,改变微生物生境[91]。
4 展望
滨海湿地是物质转化与能量流动最活跃的区域,也是微生物与矿物质交互作用形式最为多样区域,具有高度的环境异质性和微生物群落多样性,是碳氮转化的主要场所[92]。与其他生态系统微生物领域的研究相比较,滨海湿地生态系统土壤微生物的研究起步相对较晚;此外,由于滨海湿地地理位置的独特性,其土壤微生物受到的作用更为繁杂,表现出更多的变异性[24]。因此,在滨海湿地生态系统土壤微生物领域还需针对如下方向进行进一步探索。
4.1 加强全球变化多因子交互作用下滨海湿地土壤微生物的响应机制研究
在全球变化背景下,温室气体(CO2、CH4、N2O)、气候变暖和氮沉降等因子都与土壤微生物的结构和功能密切联系。目前针对滨海湿地土壤微生物对全球变化单一因子的响应机制的研究相对成熟,而缺少多因子间的耦合交互作用对土壤微生物的影响研究。全球变化各因子之间并非是相互孤立的,而是交互作用的[93]。研究全球变化多因子耦合交互作用下滨海湿地土壤微生物群落的响应机制可以更为全面地揭示全球变化背景下滨海湿地生态系统结构与功能的演变过程。
4.2 强化滨海湿地土壤微生物与环境因子的互作机理研究
近年来,滨海湿地土壤微生物的影响因子研究多停留在环境因子对土壤微生物影响的现象特征描述和线性关系表达阶段,难以定量阐明环境因子对滨海湿地土壤微生物的影响机理[94]。滨海湿地生态系统中,植被、土壤和微生物共同组成了小型的功能微系统,三者之间的互作机理是复杂而又多样的耦合机制,权衡和协同作用同时存在。因此,应加强滨海湿地生态系统中植被-土壤-微生物三者之间的互作机理研究以更好地阐释滨海湿地土壤养分的固定、转化和循环机制。
4.3 深化滨海湿地水动力条件对土壤微生物的影响机制研究
滨海湿地因频繁地受到淡水与海水交互作用的影响而具有独特的水动力条件,如土壤水的流速、流量、水体扰动等[54]。此外,除了普遍存在的垂向物质通量外,由于受潮汐作用的影响,滨海湿地生态系统还存在明显的横向物质通量,但由于当前研究手段的局限性等原因所致,关于这种横向物质通量对滨海湿地土壤微生物的影响的研究相对不足[52]。因此,应加强滨海湿地特殊水动力条件对土壤微生物影响机制的研究。
4.4 开展土壤微生物与滨海湿地生态系统物质循环综合研究
土壤微生物是滨海湿地生态系统物质循环的主要驱动力,而当前滨海湿地土壤微生物的研究多局限于微生物本身,缺乏微生物与碳、氮、硫等物质循环过程的有机结合,难以为滨海湿地物质循环过程的驱动机制分析提供更为有效、直接的基础数据[92,95]。因此应加强滨海湿地土壤微生物驱动的碳、氮、硫生物地球化学过程及其耦合机制和生态效益研究,开展驱动滨海湿地碳、氮、硫转化与循环的微生物代谢机制研究。
猜你喜欢
昆明医科大学学报(2022年2期)2022-03-29 00:51:58
食品安全导刊(2021年20期)2021-08-30 06:40:50
华人时刊(2020年21期)2021-01-14 01:33:28
科学(2020年2期)2020-08-24 07:56:56
现代装饰(2019年12期)2020-01-19 02:59:16
宝藏(2018年12期)2019-01-29 01:50:56
小学科学(学生版)(2018年8期)2018-08-21 12:05:50
微生物与感染(2015年5期)2015-12-08 07:03:20
太空探索(2015年2期)2015-07-12 11:28:16
水生生物学报(2015年1期)2015-02-28 16:01:05
