
Reexamination of the oldest pigeon (Aves: Columbidae) from Asia: Columba congi from the Early Pleistocene of Zhoukoudian, Beijing, China
2021-02-14 03:51SHENWeiThomasSTIDHAMLIZhiHeng
古脊椎动物学报(中英文) 2021年3期
SHEN Wei Thomas A.STIDHAM* LI Zhi-Heng
(1 Key Laboratory of Vertebrate Evolution and Human Origins of Chinese Academy of Sciences, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences Beijing 100044 *Corresponding author: presbyornis@gmail.com)
(2 CAS Center for Excellence in Life and Paleoenvironment Beijing 100044)
(3 University of Chinese Academy of Sciences Beijing 100049)
Abstract Columba congi is an extinct species that was described as part of the Early Pleistocene (~1.7 Ma) fauna from locality 12 of the UNESCO Zhoukoudian or “Peking Man” site in Beijing, China.Only four partial humeri of the original type series of 11 bones can be located, and the features present in those specimens do not support the original diagnosis.However, our study and redescription shows that the straight and flat margin of the pneumotricipital fossa rim (in ventral view) and the relative distal position of the dorsal supracondylar tubercle may support the continued recognition of C.congi as a valid extinct species.Columba congi appears to be the oldest fossil of Columba in Asia, and it lived during a warmer and wetter period of time of the Pleistocene with a forested Zhoukoudian.Further study of pigeons from all localities at Zhoukoudian should help to resolve questions about pigeon biogeography and evolution, including possibly the time and center of origin of the globally distributed C.livia.
Key words Zhoukoudian, Early Pleistocene, Columba, Columba congi, humerus
1 Introduction
Members of the avian genusColumba(Columbidae) commonly are referred to as pigeons or doves (largely depending on their relative body sizes), and they have represented messengers of love and symbols of peace.In addition, these birds are a common dietary resource in cultures around the world.The long history of interactions between hominins and species ofColumbaextends back tens of thousands of years with records that Neanderthals consumedC.livia40000 years ago (Blasco et al., 2014), and archeological evidence demonstrating thatC.liviahas been common food item in the Fertile Crescent more than 10000 years ago (Shapiro and Domyan, 2013).Furthermore, in the formulation of Darwin’s theory of evolution through natural selection, variation in the domestic pigeon,C.livia, played an enlarged part (Baptista et al., 2009).Pigeons continue to have an over-sized role (as compared to many other birds) in fundamental studies of evolutionary processes, interrelationships between the genotype and phenotype, and even teaching basic avian anatomy (e.g., Baptista et al., 2009; Shapiro et al., 2013; Jones et al., 2019).Despite those significant contributions to people and science, basic questions about the interrelationships of the species ofColumbaand the timing and pattern of their evolution remain elusive.
The genusColumbaincludes 35 extant (and two historically extinct) species found on all continents except Antarctica (Gill et al., 2020).The species of Columbidae are particularly rich in China with records of 31 species placed in seven genera, and in particular 10 of the 35 species ofColumba(MacKinnon and Phillips, 2000).Among those species ofColumbain China are the sister species of the Domestic Pigeon or Rock Dove (C.livia) and the Hill Pigeon (C.rupestris; Johnson et al., 2001; Soares et al., 2016), along with the high elevation Snow Pigeon (C.leuconata).
In contrast to the abundance of the extant doves and pigeons around the world, fossils of doves and pigeons are much less common.The oldest fossils of Columbidae are from the late Oligocene or Early Miocene of Australia (Worthy, 2012).While Europe has a rich record of pigeon fossils (Weesie, 1984), with a large number published from Italy (Pavia, 1999; Abbazzi et al., 2004; Sardella et al., 2005; Bedetti and Pavia, 2007; Petronio et al., 2008), fossil records are rare in Asia, with published fossil specimens from Mongolia, China, Israel, and Azerbaijan (Hou, 1982, 1993; Rich et al., 1986; Tyrberg, 1998; Martynovich, 2002; Watanabe et al., 2018).The only published fossils attributed to Columbidae in China are specimens ofStreptopeliaandColumbafrom the UNESCO Zhoukoudian “Peking Man” site caves in Beijing, which are probably best known for their abundant hominin fossil specimens (particularly “Peking Man”), stone age technologies, and mammalian fauna.However, Rich and coauthors (1986) reported the unpublished occurrence ofColumbaat a site in Hebei Province (China), but the location of those fossil specimens is unknown.
Hou (1993) published the only overview of the diverse Pleistocene bird fossils found at Zhoukoudian, and he placed the avian fossils into 13 orders and 31 families of birds, representing a great taxonomic, morphological, and ecological diversity.Columbidae is one of the most abundant groups of birds in the Zhoukoudian fossil sites, with more than 700 specimens from localities 1, 3, 4, 12, 15, and Upper Cave allocated to the extant speciesC.livia,Streptopeliachinensis, andS.orientalis, and the extinctC.congi(Hou, 1993).Those cave localities and their columbid fossils range in age from ~1.7 Ma to ~35.1-33.5 ka (Cheng et al., 1996; Li, 2018), with locality 12 as the oldest fauna with columbid fossils.Hou (1993) erected the taxonColumba congias an extinct species of pigeon based on the assemblage of larger columbid fossils from locality 12 (occurring with specimens of the smallerStreptopelia).With an estimated age of 1.7 Ma for the locality 12 fauna, the fossils ofC.congiare possibly the oldest specimens ofColumbain Asia (Tyrberg, 1998) and certainly the oldest in eastern Asia.
In the years since its publication (Hou, 1993),C.congihas been mentioned rarely in the literature, and its validity as part of the Pleistocene Eurasian avifauna appears not to have been challenged (Tyrberg, 1998; Zelenkov, 2015).With the established sister relationship betweenC.liviaandC.rupestris(which both occur in China, and Beijing in particular) and their estimated recent divergence within the Pleistocene (Soares et al., 2016), early fossils ofColumbain China may provide a window into the evolutionary origin of these commonly studied extant species.Therefore, given the age and geographic occurrence ofC.congi, its further study may provide insight into pigeon evolution and biogeography.Columba congifrom locality 12 is unique in the series of Zhoukoudian cave deposits in that Hou (1993) identified all other specimens ofColumbafrom the Zhoukoudian cave localities solely asC.livia, despite the current occurrence of bothC.liviaandC.rupestrisin Beijing and around Zhoukoudian.
There are 27 cave localities collectively called Zhoukoudian.Locality 18 is thought to contain the earliest fauna dating back to 1.9 Ma (Cheng et al., 1996), and the Upper Cave (locality 26) is the youngest with an estimated age of 35.1-33.5 ka (Li, 2018).Of course, columbid fossils are not present in all cave faunas.Hou (1993) consideredColumba congito be a kind of small primitive pigeon, and locality 12, where all known specimens of this extinct species derive, is a pocket-shaped cave with a fauna from the middle Early Pleistocene, ~1.70 Ma (Cheng et al., 1996).The upper part of the deposit is composed of hard dark red sediments, and the lower part is made of sands and gravels (Teilhard de Chardin, 1938; Cheng, 2002).Cheng (1997) interpreted the mammalian fauna from locality 12 as deriving from a hot and humid dense forest.Representative animals from locality 12 include more than 20 species, including rhinoceroses (Rhinocerossp.), burrowing zokors (Siphneussp.), an extinct monkey (Procynocephalus wimani), saber-toothed cats (Machairodussp.), and hyraxes (Postschizotherium chardini) (Huang, 1960).In contrast to the rich mammalian diversity in locality 12 deposits,Columba congiand doves referable toStreptopeliasp.are the only birds reported (Hou, 1993).That mammalian fauna contrasts significantly with the extant fauna of northern China (lacking rhinos, monkeys, and hyraxes), and suggests a distinctly different environmental and climatic regime in the Early Pleistocene.
Institutional abbreviationsBMNHC, Beijing Museum of Natural History Collection, Beijing; IOZ, Institute of Zoology, Chinese Academy of Sciences, Beijing; IVPP, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing; OV, modern osteological specimen in the IVPP; V, fossil specimen in the IVPP.
Materials examinedModern skeletons:Columbalivia, BMNHC 21449, IVPP OV 1363, OV 1735;C.rupestris, IOZ S031 and OV 859;Streptopeliachinensis, OV 1846 and OV 1908;S.orientalis, BMNHC 108613, OV 1883, and OV 1882.Fossil specimens:C.congi, V 6436.Measurements were taken with digital calipers and rounded to the nearest 0.1 mm.
Anatomical nomenclatureTerminology for osteological features follows Baumel and Witmer (1993) with English equivalents of the Latin terms.
2 Systematic paleontology

HolotypeHou (1993) designated the specimen number IVPP V 6436 as the holotype ofC.congi.However, that single specimen number refers to 11 fragments of bones from locality 12 including parts of coracoids, ulnae, and humeri.Hou (1993) did not state how many specimens of the total of 11 are humeri, ulnae, or coracoids (or other parts of the skeleton), and he did not indicate which of these 11 specimens is the individual holotype specimen for the species.
A search of the Zhoukoudian collections housed in the IVPP resulted in only four specimens total labeled asC.congifrom locality 12 being located.Those four osteologically adult specimens (with smooth fully ossified bone surfaces) are one proximal left humerus and three distal right humeri representing a minimum of three individuals (Fig.1).All are labeled as IVPP V 6436.The location of the other pigeon fossil materials mentioned and illustrated by Hou (1993) is unknown at present.None of the four specimens appear to be those that were illustrated by Hou (1993), and thus do not appear to strictly represent the holotype for the species.They can be treated as paratypes since they are part of the original type series designated by Hou (1993).We designate the four humeri as IVPP V 6436.1-4 in order to facilitate communication about individual specimens and for future reference (Fig.1; see description below).
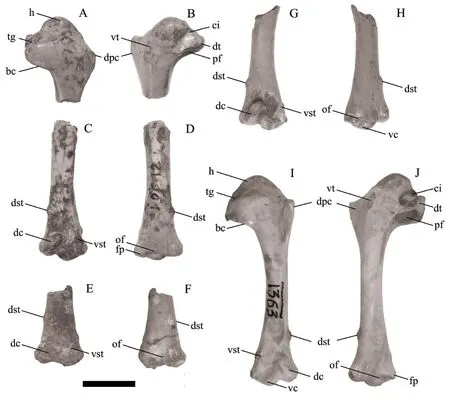
Fig.1 Comparative images of the humerus of the extinct Columba congi (A-H) and Columba livia (I, J)A, B.proximal end of left humerus of IVPP V 6436.1 in cranial (A) and caudal (B) views; C-H.distal end of right humerus of C.congi in cranial (C, E, G) and caudal (D, F, H) views: C, D.IVPP V 6436.2; E, F.V 6436.3; G, H.V 6436.4; I, J.left humerus of IVPP OV 1363 in cranial (I) and caudal (J) views Abbreviations: bc.bicipital crest; ci.capital incisure; dc.dorsal condyle; dpc.deltopectoral crest; dst.dorsal supracondylar tubercle; dt.dorsal tubercle; fp.flexor process; h.humeral head; of.olecranon fossa; pf.pneumotricipital fossa; tg.transverse ligamental groove; vc.ventral condyle; vst.ventral supracondylar tubercle; vt.ventral tubercle.Scale bar = 10 mm
DiagnosisHou’s (1993:271) original diagnosis in English states, “A more primitive and smallerColumba, the wall thick and bone cavity small.The pneumatic fossa of the humerus not developed, no pneumatic foramen or very small.The humeral shaft flat and out border straight; the internal condyle of the distal end of the humerus not developed.The ulna shortly curved, smaller than that ofC.livia.” No further description of the specimens is provided in English, but in Chinese he says (Hou, 1993:201), “the humeri have a small and curved crista deltopectoralis, shallow sulcus ligament transversus, small process supracondylaris dorsalis, large epicondyles, and undeveloped internal condyles.The only remaining coracoid is seemly straight and robust but not well preserved.”
The diagnoses (in Chinese and English) do not actually diagnose this species as separate from any other species ofColumba, and many of the features are not present among the available fossils.The size of humerus ofC.congiis smaller thanC.rupestrisandC.livia, and slightly larger thanStreptopelia orientalisandS.chinensis(Table 1).That aspect of the original diagnosis appears correct.The statement about the wall thickness does not appear accurate.The bone thickness is not supported as a diagnostic feature since it is not significantly thicker in our analyses (see details below).It is possible that Hou’s (1993) interpretation of a thick-walled bone could have been based on the presence of medullary bone (of an egg-laying female), but that is merely speculation until the original specimens can be located.In addition, the humerus is a pneumatic bone in pigeons, and should not have been filled with red blood cell producing marrow (Schepelmann, 1990).The only proximal humerus among the four bones, IVPP V 6436.1, clearly preserves a pneumatic opening in the pneumotricpital fossa similar in size with that in extant skeletons; contradicting the statement about its absence in this extinct species.The preserved shafts of the humeri also do not differ visually from those of modern skeletons in their flatness and borders.We believe that Hou’s (1993) statement about the ‘internal condyle’ refers to the ventral condyle which is clearly present (except where broken off in V 6436.3) or possibly the ventral supracondyle which appears similar to other pigeons.We are not able to locate the ulna illustrated by Hou (1993), and cannot verify any potential diagnostic features in that skeletal element.

Table 1 Measurements of fossil and modern columbid humeri (mm)
The deltopectoral crest of columbids and pteroclids has a very distinct pointed triangular outline that contrasts with the stated presence of a rounded crest inC.congi(Li et al., 2020).That statement also is contradicted by the shape presented in the photograph of the humerus ofC.congi(Hou, 1993:fig.11 on plate 6) and the morphology in IVPP V 6436.1.The transverse ligamental sulcus is not any smaller than the condition in extant comparative specimens (Fig.2).The relative sizes of the dorsal and ventral supracondylar regions are consistent with the range of variation in extant skeletons.We could not locate the coracoid to verify any potential diagnostic features of the bone.
Taken in total, we do not see any of the features listed by Hou (1993) except for size, as actually differentiating these fossils from any extant or extinct species.In part, that is because it appears that many of the features actually are not present in the specimens available.However, our close examination of the fossils reveals morphological characters that are diagnostic at a variety of taxonomic levels, including potentially that of species.
While the humerus exhibits many features common to Columbidae and Pteroclidae such as the large triangular deltopectoral crest, and a dorsal supracondylar tubercle located proximally, the humeral specimens from locality 12 display typical features of Columbidae including having an inset olecranon fossa, and they lack several features present in their close relatives Pteroclidae (like an indistinct olecranon fossa; Li et al., 2020).Comparisons with other taxa help to place the four fossil specimens inColumbausing the following characteristics: humerus, proportionately more robust, shaft straighter, proximal and distal ends more expanded (Hume, 2011).Species ofStreptopeliahave an olecranon fossa that is a wider and shallower area, and these fossils also are distinctly larger than individuals of the dove species inStreptopelia(Table 1).
Tentative revised diagnosis: a couple of features appear to differ from other Pleistocene fossils and extant skeletons ofColumbaand may be diagnostic for this species.In ventral view, the caudodistal ventral margin or rim of the pneumotricipital fossa is straight in the proximal humerus ofC.congi(Fig.2), but the muscle origination scar on that rim projects distally (making the margin slightly convex) inColumbarupestrisandC.livia.Though our sample sizes are small, this may be a diagnostic feature.The dorsal supracondylar tubercle also is positioned more distally in all distal humeri fossils as compared toC.liviaandC.rupestris(see description below; Table 1).

Fig.2 Comparison of proximal pigeon humeri in ventral view illustrating a potential diagnostic featureA.Columba congi (IVPP V 6436.1); B.C.rupestris (IVPP OV 869)Arrow indicates the convex margin of the rim of the pneumotricipital fossa Abbreviations: bc.bicipital crest; h.humeral head; tg.transverse ligamental groove.Scale bar=5 mm
The shared absence of these features inC.liviaandC.rupestriscould suggest that they represent synapomorphies of those species and a phylogenetic position ofC.congioutside of the sister relationship between those extant species hypothesized based on molecular data (Johnson et al., 2001; Soares et al., 2016).Conversely, they could be plesiomorphies and indicate a derived character state inC.congi.We are unable to differentiate between the near identical humeri ofC.rupestrisandC.livia, but other portions of their skeletons exhibit variable characters (like the presence or absence of lateral trabeculae on the sternum).In addition, the great age of theC.congifossils as compared to the other cave sites at Zhoukoudian (and their fossils ofColumba) also could point to a sister or even anagenetic type of evolutionary relationship with the current species in Beijing.
LocalityLocality 12, Zhoukoudian, Fangshan District, Beijing, China; 200 m to the south of locality 1.The cave deposit in a vertical cylindrical pit four meters high with one meter diameter, the upper part of the deposit is hard dark red sediment, the lower part is coarse sand and gravel (Teilhard de Chardin, 1938), and the age of the deposit is middle Early Pleistocene, ~1.70 Ma (Cheng et al., 1996).Columba congiandStreptopeliasp.are the only avian fossils present in Locality 12.However, more than 20 mammalian species have been recorded, most of which are forest-linked mammals including bears (Ursuscf.U.angustidens), saber-toothed cats (Megantereonsp.), leopards (Pantheracf.P.pardus), and rhinoceroses (Rhinocerossp.; Huang 1960; Cheng, 1997).
DescriptionCurrently, all specimens ofColumbafrom locality 12 are designated as IVPP V 6436 including those illustrated and discussed by Hou (1993) and the four specimens described here.In order to accurately document the known specimens, we informally refer to each specimen with a number 1-4 after the specimen number.Thus, V 6436.1 refers to the proximal humerus.The three distal right humeri are V 6436.2, V 6436.3, and V 6436.4 (see Fig.1 for illustrations of each specimen).V 6436.4 and V 6436.2 preserve the most of the humeral shaft (over half of the humeral length), and V 6436.3 is the least complete and missing the distal condyles.The dorsoventral width of the proximal end from the dorsal tubercle to the ventral edge of the bicipital crest is 12.8 mm.The dorsoventral width of the distal end is 10.2-10.3 mm (Table 1).The smallest diameter of the humeral shaft is 4.8-4.9 mm.The thickness of the humeral shaft bone is 0.3-0.4 mm.
The proximal left humerus (IVPP V 6436.1) is missing the ventral tubercle and the apex of the deltopectoral crest, and the shaft is broken distal to the distal end of the deltopectoral crest.The deltopectoral crest is a large triangular projection with a short proximal side and a longer distal margin.The humeral head projects distinctly proximally and has a slight concavity on its cranial ventral aspect.The caudal face of the humeral head is flush with the adjacent distal humeral surface.The ventral tubercle is an ovoid scar that is flush with the adjacent humeral surface, except for its distal end that is very slightly raised.The capital incisure is a wide U-shaped concavity that does not undercut the head.The dorsal crus of the pneumotricipital fossa is short, and does not extend as far distal as the bicipital crest.A muscle scar (that widens distally) extends from the distal end of the dorsal crus to the broken end of the specimen.The distal end of the bicipital crest is distal to the apex of the deltopectoral crest, and in cranial view the distal margin of the bicipital crest is straight.The pneumotricipital fossa is partially walled, but has a central pneumatic opening (as in other specimens ofColumba).In ventral view, the ventral rim of the pneumotricipital fossa has a straight margin that is inclined caudoproximal to craniodistal (Fig.2).The caudal rim of the transverse ligamental sulcus adjacent to the capital incisure is a relatively thin ridge.The bicipital crest meets the shaft at an obtuse angle.
The distal end of the humerus is characterized by the ventral condyle extending distal to the dorsal condyle.The dorsal supracondylar tubercle is positioned far proximal relative to the distal end.The tubercle is located 9.3-9.5 mm proximal to the distal end in the fossil specimens (as compared to 9.6-11.3 mm in extant specimens; Table 1).The tubercle is not elongate proximodistally, and is positioned proximal to the proximal end of the brachial fossa.There is a shallow, arcuate fossa around the proximal end of the dorsal condyle that connects ventrally with the brachial fossa.The brachial fossa is a proximodistally elongate ovoid that is deepest ventrally.The proximal end of the dorsal condyle is at the same proximodistal level as the ventral supracondylar tubercle.The facet for the ventral collateral ligament has a very rounded subtriangular outline, is slightly convex on its surface, and faces slightly distally (not entirely cranially directed).The ventral supracondylar muscle scar for one of the pronator muscles is located proximal to the level of the ventral supracondylar tubercle.The area ventral to the ventral supracondylar tubercle and facet for the ventral collateral ligament is concave and directed somewhat cranioventrally.The scapulotricipitalis groove is present and extends proximally to about the same level as the proximal end of the dorsal condyle.The ventral margin of the groove projects more caudally than its dorsal margin.The olecranon fossa is a wide shallow area, but contains a distinctly inset concavity that cuts into the caudal side of the ventral condyle and extends towards the flexor process.The dorsal margin of the bone dorsal to the dorsal condyle is slightly concave.
3 Discussion
There has been a lack of reliable evidence for the origin of the crown group of Columbidae (Shapiro et al., 2002).In addition, the temporal and biogeographic origins ofColumbaand the ubiquitousC.liviaare unknown at present.In part, this is the result of a low level of molecular sampling among the species ofColumbafor phylogenetic analysis (Johnson et al., 2001; Soares et al., 2016).With the exception of the globally distributedC.livia, the species ofColumbaare found exclusively in the Old World with the most species endemic to Africa and Asia (Gibbs et al., 2001).Given that the distribution of the sister species toC.livia(i.e.C.rupestris) is restricted to Asia, and a large number of species ofColumbaare endemic to Asia (14 species), it would appear possible, if not likely, that the cosmopolitanC.livialineage originated in Asia.However, answering that question will require additional phylogenetic and paleontological data.
Records ofColumbaand fossils referred toC.liviahave been published from the Pliocene and Pleistocene across the Old World (Tyrberg, 1998).As yet, those identifications (and that of other fossils attributed to species ofColumba) have not been tested via phylogenetic or other quantitative analyses.As we see in our study, the potential diagnostic features are few and not yet tested within a larger phylogenetic context of the cladeColumba.While there are abundant pigeon fossils from the Pleistocene of Europe (Tyrberg, 1998), the origin ofColumbaremains elusive.Molecular clock estimation places the split betweenC.liviaandC.rupestrislate in the Pleistocene (Soares et al., 2016), and that is much younger than some of the oldest fossils attributed toC.livia(i.e.Pliocene).Either the molecular clock estimates are severely underestimating the time of divergence or the plethora of early fossils identified asC.liviabelong to other lineages.We will be able to clearly address these questions only through greater sampling of species ofColumbaand with the construction of a morphological phylogenetic context for those species (extinct and extant).Our study ofC.congidemonstrates both the problems and the potential outcomes from such a broader research approach.While we dismantled most of the features previously used to recognizeC.congias a separate species, we uncovered others that may withstand further sampling and testing and documentC.congias a valid species.In addition, the great age of these fossils along with their apparent morphological differences could, along with a study of the younger specimens identified asC.liviafrom Zhoukoudian, narrow hypotheses about the evolutionary origin ofC.livia, and pigeons in Asia.
The sediments preserved in locality 12 represent a cave deposit.At present, there is not much sediment left at locality 12, and in recent years, the site was backfilled (Cheng, 2002).No additional specimens ofC.congiwill be found there.Columba congiis only known from locality 12 with its fauna representing a hot and humid climate, with abundant surface water and well-developed forests (Cheng, 1997).That hypothesized Early Pleistocene habitat contrasts with the preferences of the extantC.liviaandC.rupestriswhich prefer more open environments (Goodwin et al., 1983).The occurrence ofC.congiin a wet forest habitat along with an extinct monkey and leopards could indicate that its phylogenetic (and biogeographic) relationships might be closer to those columbid species present in southern China (and Asia) today.Hou (1993) identifiedC.congionly at locality 12 and designated all specimens from all other (younger) cave sites at Zhoukoudian asC.livia.The proposed difference in the inferred habitats of locality 12 may relate to the presence ofC.congiduring that time period alone, and its absence in other caves (and younger times) that represent more open habitats preferred by the extant species.A study of stable isotopes from the fossils possibly could help to resolve any habitat or ecological differences among the specimens ofColumbapresent through the Pleistocene at Zhoukoudian.Furthermore, we recommend that future study examine the geologically younger fossil columbid material, particularly with reference to its potential placement withinC.rupestris, the sister species ofC.liviaand a current resident species in Beijing.
AcknowledegmentsWe thank Song Gang and Liu Di for access to modern skeletons at Institute of Zoology, Chinese Academy of Sciences (Beijing) and the Beijing Museum of Natural History Collection.We also thank the reviewers for valuable comments on an earlier draft of the manuscript.

- 古脊椎动物学报(中英文)的其它文章
- A new species of Pteronisculus from the Middle Triassic (Anisian) of Luoping, Yunnan, China, and phylogenetic relationships of early actinopterygian fishes
- Taxonomic revision of the holotype of Proboselaphus watasei Matsumoto, 1915 (Bovidae, Artiodactyla) from Chuanyu area, China
- A juvenile skull of the longirostrine choristodere (Diapsida: Choristodera), Mengshanosaurus minimus gen.et sp.nov., with comments on neochoristodere ontogeny
- Exceptional preservation of an extinct ostrich from the Late Miocene Linxia Basin of China
- 《古脊椎动物学报(中英文)》编委会