
不同生长期薰衣草枝叶功能性状及其与环境因子的相关性1)
2021-02-10 11:17李玲孙宇婷任杏莫丽娜侯一航苑泽宁
东北林业大学学报 2021年12期
李玲 孙宇婷 任杏 莫丽娜 侯一航 苑泽宁
(哈尔滨师范大学,哈尔滨,150025)
植物多为构件生物,植株是由叶构件、枝构件等组成的构件系统[1],植物通过构件在数量、形状和空间上的变化实现对环境的适应[2]。构件及其功能性状是连接植物与环境的桥梁,能反应植物对环境的适应能力以及环境对植物的影响程度[3-5]。枝条与叶片作为植物地上构件重要组成部分,在养分和水分运输、机械支撑[6]、光合同化[7]等生理活动中发挥重要作用。构件生物量是物质和能量积累的基本体现[3],植物在生长季的生物量积累能力是评价其适应性的指标之一[8]。叶面积与植物的蒸腾、光合及呼吸作用等生理活动密切相关,与叶面积相关的性状均可反应植物对环境的适应[9-14]。所以对植物枝叶功能性状进行研究有助于揭示植物对环境的适应机制。
薰衣草(LavandulaangustifoliaMill.)为唇形科小灌木,精油品质极佳,是我国生产精油的主要栽培品种。目前,薰衣草在我国主要种植于新疆,在甘肃、北京、黑龙江等地也有分布[15-18]。关于薰衣草的引种栽培已有较多研究成果[17],主要目的是增加多年生花卉资源,丰富花卉优良品种,美化城市居住环境;但关于薰衣草在北方高寒地区生长的生态适应性方面的研究较少。为此,本研究在黑龙江省哈尔滨市松北区设置3个10 m×10 m的样方,选取薰衣草30株,于不同生长期每株随机选取10个当年生枝条,测量每个枝条的叶片数量、总叶面积、枝条和叶片质量,计算枝条和叶片功能性状;分早晨、中午、傍晚3个时段,在样方区域按5点法监测环境因子。依据试验统计监测数据,应用方差分析法、显著性差异法、相关性分析法、冗余分析法等,分析薰衣草枝叶功能性状在不同生长期期间的差异显著性、功能性状变异性、枝叶性状间相关性及其与环境因子的关系,探索薰衣草在哈尔滨地区的生长规律及其对环境因子的适应性;旨在为寒冷的东北地区引种栽培薰衣草提供技术参考。
1 研究区域概况
研究区域位于黑龙江省哈尔滨市松北区(126°32′49″~126°33′11″E,45°51′50″~45°52′N),该地区属于温带大陆性季风气候,地势平坦,四季分明;全年平均气温5.44 ℃,集中降水期为7—8月份,集中降雪期为11月份至次年1月份。春季平均气温7.06 ℃,气温回升快,易出现波动;夏季平均气温21.39 ℃,炎热湿润多雨;秋季平均气温6.39 ℃,降水明显减少,昼夜温差较大;冬季平均气温-13.06 ℃,漫长而寒冷。试验地以黑钙土为主,土壤中含有丰富的有机质,土壤透气性和保水性强。
2 材料与方法
薰衣草(L.angustifolia)在该地区的生长物候期:每年4月下旬至5月上旬开始返青,5月中旬至6月末为营养生长期,7月上旬进入花期,盛花期为7月中旬至9月下旬,种子成熟于9月份至10月份,11月份至次年3月份为休眠越冬期。取材时间分别是生长初期(6月上旬)、旺盛期(8月中旬)、生长末期(11月下旬)。
在研究区域设置3个10 m×10 m的样方,选取长势相近、健康的薰衣草30株作为样本。在每株随机选取10个无叶面损伤、长势相近的当年生枝条,所取枝条分别装入纸袋,带回实验室,每次取样时进行叶片叶绿素相对吸光值[19]监测。
以枝条为单位,统计每个枝条的叶片数量(片),用植物叶面积仪(YMJ-OH)测量植物总叶面积/枝,用分析天平(FA2004)分别测枝条和叶片鲜质量;然后,将枝条、叶片分别置于烘箱(WGL-125B)中70 ℃烘干48 h至恒质量,再分别测枝条、叶片干质量。
功能性状的计算:①枝条相对含水量=(枝条鲜质量-枝条干质量)/枝条鲜质量;②枝条总含水量=(枝条鲜质量-枝条干质量)/枝条干质量;③叶片相对含水量=(叶片鲜质量-叶片干质量)/叶片鲜质量;④叶片总含水量=(叶片鲜质量-叶片干质量)/叶片鲜质量;⑤单叶面积=总叶面积/叶片数量;⑥单叶干质量=叶片干质量/叶片数量;⑦叶面积比=总叶面积/枝条干质量;⑧比叶面积=总叶面积/叶片干质量;⑨比叶质量=叶片干质量/总叶面积;⑩叶干物质量=叶片干质量/叶片鲜质量;叶枝质量比=叶片干质量/枝条干质量[12,20-21]。
环境因子测定:每次取样时,在样方区域按5点法进行环境因子监测,分早晨、中午、傍晚3个时段进行。采用土壤pH计(KS-06)测土壤pH,采用土壤ZD-EC仪测土壤电导率,采用土壤温度计(TPJ-21-6)测土壤温度,采用温湿度计(8828)测空气温度、空气湿度,采用照度仪(TPJ-22-1)测光照强度。每个区域按5点法每点取土样,带回实验室,用分析天平测土壤鲜质量;然后置于烘箱(WGL-125B)中70 ℃烘干48 h至恒质量,测土壤干质量,计算土壤相对含水量=(土壤鲜质量-土壤干质量)/土壤鲜质量、土壤绝对含水量=(土壤鲜质量-土壤干质量)/土壤干质量。
采用SPSS25.0中单因素方差分析法分析薰衣草枝条叶片功能性状在生长初期、旺盛期、生长末期期间的差异显著性,运用最小显著性差异法进行多重比较分析(P<0.05)。用Excel 2010进行功能性状变异性分析,用皮尔逊(Pearson)相关性分析法进行枝条叶片性状间相关性分析(P<0.05,P<0.01),运用生态学数据处理软件(Canoco 5)对环境因子与植物功能性状进行冗余分析(RDA)。
3 结果与分析
3.1 薰衣草枝条叶片生长性状
薰衣草枝条鲜质量、干质量、相对含水量、总含水量,在不同生长阶段间均具有显著性差异(见表1)。由表1可见:鲜质量在生长旺盛期约是初期的19倍、末期的1.3倍。干质量,随生长阶段的延续呈上升趋势,在生长末期达到最大值,是初期的近36倍。相对含水量和总含水量的变化趋势相似,均在生长旺盛期达到最大值,总含水量在各时期均高于相对含水量。

表1 薰衣草不同生长阶段枝条生长性状
由表2可见:叶片叶绿素相对吸光值随生长时间延续呈下降趋势,生长初期和旺盛期是末期的近2倍。总叶面积、单叶面积、比叶面积、叶鲜质量、干质量、相对含水量、总含水量,在生长旺盛期均为最大值。在生长旺盛期:叶鲜质量是初期和末期的6倍,干质量是初期的近4倍、末期的2倍,相对含水量是末期的近2倍,总含水量是末期的5倍,总叶面积是初期和末期的近5倍,比叶面积是末期的2倍。

表2 薰衣草不同生长阶段叶片生长性状

注:表中数据为“平均值±标准差”;数据后,同列不同小写字母,表示差异显著(P<0.05)。
单叶干质量随生长时间延续呈上升的趋势,在生长末期为最大值,是初期的近2倍。叶面积比、叶枝质量比均在生长初期较高,分别是生长末期的37倍、21倍,比叶质量和叶干物质量的变化趋势相似,均在生长旺盛期最小,在末期最大。
3.2 薰衣草枝条叶片功能性状相关性
由表3、表4可见:薰衣草生长初期和旺盛期,枝条叶片功能性状相关性较为密切。在这两个时期,枝条鲜质量,与枝条干质量、总叶面积、叶鲜质量呈正相关,与叶面积比、叶枝质量比呈负相关。枝条干质量,与总叶面积、叶鲜质量、叶干质量、单叶干质量呈正相关,与叶相对含水量、叶总含水量、叶面积比、叶枝质量比呈负相关。枝条相对含水量,与枝总含水量、叶相对含水量、叶总含水量、比叶面积呈正相关,与叶片干质量、比叶质量、叶干物质量呈负相关。枝条总含水量,与叶相对含水量、叶总含水量、比叶面积呈正相关,与叶干质量、比叶质量、叶干物质量呈负相关。总叶面积,与叶鲜质量、叶干质量、单叶面积呈正相关。叶鲜质量与叶干质量呈正相关。叶干质量,与单叶干质量、比叶质量、叶干物质量呈正相关,与叶片相对含水量、叶片总含水量、比叶面积呈负相关。叶含水量与比叶面积呈正相关,与比叶质量、叶干物质量呈负相关。单叶干质量,与比叶质量呈正相关,与比叶面积呈负相关。叶面积比,与叶枝质量比呈正相关。比叶面积,与比叶质量、叶干质量呈负相关。比叶质量与叶干质量呈正相关。

表3 生长初期薰衣草枝条叶片功能性状Pearson相关性

续(表3)

表4 生长旺盛期薰衣草枝条叶片功能性状Pearson相关性
由表5可见:生长末期性状之间的相关性有所下降,与前两个时期相比也表现为正相关的有:总叶面积与叶片干质量、叶片鲜质量,叶片鲜质量与干质量。表现为负相关的有:枝条鲜质量与叶面积比、叶枝质量比,枝条干质量与叶面积比、叶枝质量比,叶片相对含水量、叶片总含水量与叶干物质量,比叶面积与比叶质量。

表5 生长末期薰衣草枝条叶片功能性状Pearson相关性
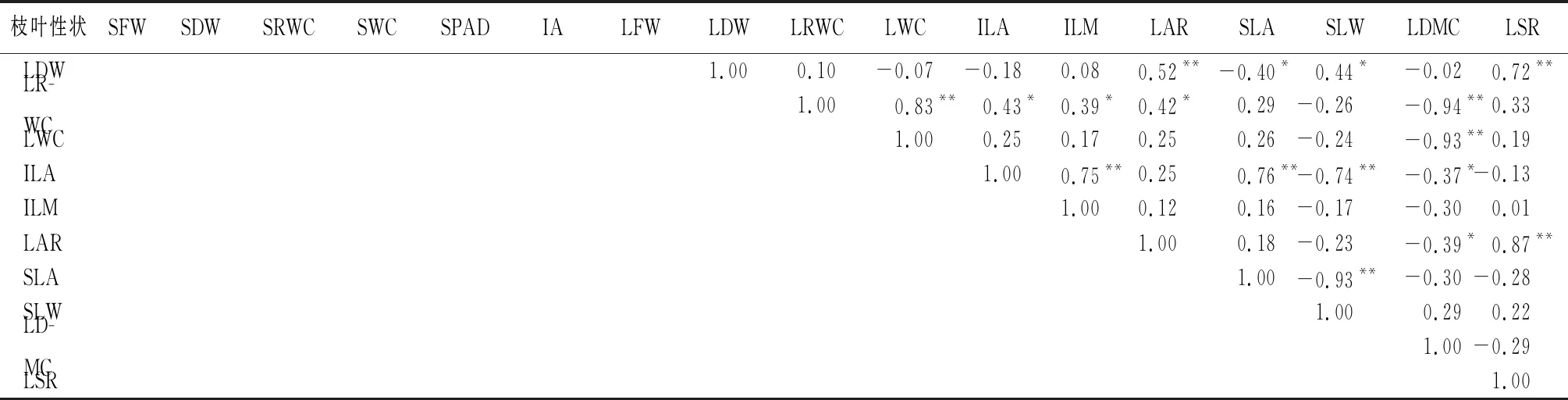
续(表5)
3.3 薰衣草枝条叶片功能性状变异性
由表6可见:薰衣草枝条功能性状变异系数在1.82%~33.48%。其中:干质量和鲜质量变异系数,由大到小依次为生长初期、末期、旺盛期;相对含水量和总含水量变异系数,由大到小依次为生长末期、初期、旺盛期。
由表6可见:叶片功能性状变异系数在3.06%~53.49%。其中:叶片叶绿素相对吸光值、总叶面积、鲜质量、干质量、相对含水量、单叶面积、单叶干质量、叶面积比、叶枝质量比、枝条总含水量的变异系数,由大到小依次为生长末期、初期、旺盛期;叶片总含水量、比叶面积、叶干物质量变异系数,由大到小依次为生长末期、旺盛期、初期;比叶质量变异系数,由大到小依次为生长旺盛期、末期、初期。

表6 不同生长阶段薰衣草枝条叶片功能性状变异性
3.4 薰衣草枝条叶片生长与环境因子相关性
根据冗余分析,对所得的枝条、叶片功能性状及环境因子进行筛选,将筛选出的6个环境因子、4个枝条性状、13个叶片性状进行冗余分析。由表7可见:排序轴1、排序轴2的特征值分别为0.831 0和0.139 2;薰衣草枝叶生长与环境因子的相关性,在第1排序轴、第2排序轴分别为0.991 7和0.993 8。冗余分析前4轴方差累计贡献率为97.65%,其中前2轴方差累计贡献率为97.02%,对薰衣草枝叶生长和环境因子关系方差累计贡献率为99.35%,提取的生态信息量大,具有显著的生态意义。由此可知,前2轴能很好地反应薰衣草枝叶生长与环境因子的关系,且主要由第2排序轴决定。

表7 薰衣草枝叶生长与环境因子的相关性和各排序轴方差累计贡献率
为研究环境因子影响薰衣草枝叶生长的重要程度,对6个环境因子分别进行前向选择排序,得到环境因子重要性排序(见表8)。由表8可见:环境因子对薰衣草枝叶生长影响重要性,由大到小依次为电导率、土壤温度、最低气温、空气湿度、土壤绝对含水量、平均气温。其中:电导率对薰衣草枝叶生长的影响呈极显著水平(P<0.01),贡献率为81.5%;土壤温度、最低气温对薰衣草枝叶生长的影响呈显著水平(P<0.05),贡献率分别为44.4%、41.8%。

表8 环境因子对薰衣草枝叶生长的重要性
冗余分析可以同时在1个图上表示植物生长的时期(质心)、环境因子(空心箭头)、植物生长指标(实心箭头)之间的排序,更能直观地显示三者之间的关系。环境因子与排序轴之间的正负相关性,用箭头所在的象限表示,箭头连线的长度代表着某个环境因子或植物生长指标与植物相关程度的大小,连线越长,代表此环境因子对植物的生长影响越大。箭头之间的夹角越小代表着环境因子的相关性越大,箭头与排序轴的夹角代表着某个环境因子与排序轴的相关性大小[22]。由图1可见:在生长初期,薰衣草枝叶生长与平均气温的相关性最大,旺盛期的枝叶生长与空气湿度、电导率的相关性最大,末期的薰衣草枝叶生长与土壤绝对含水量相关性最大。功能性状与环境因子的关系为:土壤绝对含水量与枝条干质量相关性最大,最低气温与叶片叶绿素相对吸光值相关性最大,空气湿度与叶鲜质量、总叶面积相关性较大,电导率与叶总含水量相关性最大。
4 讨论
4.1 不同生长阶段薰衣草枝条叶片生长适应性
已有研究表明,在水分亏缺情况下,高粱鲜质量累积减少,反之,鲜质量增加[23]。本研究中,薰衣草枝条鲜质量、含水量在生长旺盛期达到峰值,且枝条鲜质量与土壤含水量呈正相关(见表1、图1),表明该时期植物体内水分充盈,代谢旺盛,有利于物质运输,促进植株生长。较高的叶片相对含水量可使植物增强抗失水能力[24]。已有研究表明,枝条相对含水量与植物的抗寒性有关,枝条相对含水量越低,植物的抗寒性越强[25]。本研究中,生长末期的薰衣草枝条相对含水量最低(见表1),此时温度呈下降趋势,植物体内含水量的下降有利于增强其抗寒性,为越冬做准备。枝条干质量与叶面积的变化,可揭示植物应对逆境的生长策略[26],枝条干质量越大,总叶面积越小,植物的抗逆性越强。薰衣草在生长末期,枝条干质量达到最大,而总叶面积为最小(见表1、表2),枝条干物质积累增大,叶面积减小以降低消耗均有利于植物抗寒性的提高。
薰衣草叶片叶绿素相对吸光值在生长初期、旺盛期处于较高水平,总叶面积和单叶面积在旺盛期达到最大值(见表2),叶片光合作用强,有利于积累有机物。已有研究表明,叶面积比随枝条质量的增加而减小,对于优化叶片、枝条的资源分配具有重要意义[27]。本研究中,叶面积比在生长初期最大,随植物生长到末期,叶面积比显著下降(见表2),使枝条积累的光合产物增多,为植物抵御低温奠定物质基础。比叶面积和叶干物质量反映植物对资源获取和转化利用间的平衡能力[28-29],植物比叶面积值大是适应环境的体现[20]。薰衣草的比叶面积在生长旺盛期时最大(见表2),表明其对环境的适应性较好。伴随着植物叶片干物质量的增加,比叶面积减小,使叶片内的水分向表面扩散阻力增大,进而降低植物蒸腾作用丧失的水分,增强在胁迫的环境中适应性[20,30]。本研究中,生长末期的薰衣草,比叶面积显著下降(见表2),有利于增强植物对低温胁迫环境的适应。
单叶干质量和叶干物质量,反映植物生物量的累积[31],植物对环境资源的利用能力[32]和对体内养分的保持能力[33]。本研究中,薰衣草在生长末期,叶片中积累的有机物最多,叶干物质量达到最高(见表2),为种子成熟和植株越冬提供养分。已有研究表明,较高的比叶质量有利于减缓叶片衰老,维持较高的光合速率,促进干物质的积累[34]。薰衣草比叶质量,在生长末期显著升高,与单叶干质量和叶干物质量在末期显著升高相吻合(见表2),为植物应对寒冷的冬季奠定物质基础。植物干物质分配是植物适应逆境胁迫的重要策略之一[35]。在胁迫条件下,许多植物通过调整自身的物质分配格局,提高对胁迫的适应性[36]。本研究中,薰衣草叶枝质量比,随生长时期变化呈下降趋势,在生长末期达到最低(见表2),叶生物量减少的同时,枝条生物量增多,营养物质的分配格局发生了变化,为植物越冬提供一定的资源和能量。
4.2 不同生长阶段薰衣草枝条叶片生长相关性与变异性
枝条与叶片功能间的平衡,是植物协调生长的重要生物学特性[37],反映植物对生境的适应能力[38-39]。本研究中,薰衣草在生长的初期和旺盛期,枝条叶片生长相关性高于末期(见表3、表4),表明在植株形态建成及生殖生长时期,枝条叶片协同生长。植物在枝条和叶片的生物量投入和水分含量上表现出一定的规律性[40]。水分含量影响植物光合作用、蒸腾作用、净初级生产和物质运输等生理生化过程[41],也表征植物代谢速率[42]。总叶面积大,生物量的生产和运输相对较大,需要植物体内较多的水分来参与光合作用和物质运输等生理生化过程[43-44]。本研究中,生长旺盛期,枝条含水量与总叶面积呈正相关(见表4),与上述结论相近,有利于提高植物的光合作用和物质运输速率,促进有机物积累。
变异系数反映数据离散程度,变异系数≤0.2为弱变异,介于0.2~0.5为中等变异,变异系数≥0.5为强变异[45]。本研究中,生长旺盛期,各个功能性状的变异性为弱变异,普遍低于初期和末期(见表6)。除薰衣草在生长末期的叶片总含水量为强变异外,枝条叶片的其他功能性状均为弱变异到中等变异(见表6),表明植株生长良好。在植物生长初期,易受外界环境影响,性状变异系数偏大,自然选择的压力促使植物各性状间沿最优化的方向发展[46]。薰衣草在生长初期具有较强可塑性,这种特性可促使植物进行最佳的资源分配,以提高自身生存的适合度[26]。到生长旺盛期,薰衣草枝条生长趋于稳定,枝条叶片功能性状变异性下降,对于植物的开花结实具有重要的保障。至生长末期,植物面临冬季到来,枝条叶片的物质代谢发生变化以应对低温环境,导致各功能性状的变异性有所升高(见表6)。
4.3 薰衣草枝条叶片生长与环境因子的关系
已有研究表明,春季气温越高,植物物候发生期越早;反之,物候发生期推后[47]。本研究中,生长初期的薰衣草,主要受平均气温的影响(见图1),薰衣草在哈尔滨地区的返青时间为4月下旬,对当地的环境温度表现出较好的适应性,是完成生活史周期的重要前提。处于生长旺盛期的植物,主要受土壤电导率的影响,电导率是判断土壤中盐离子是否限制植物生长的基础[48],电导率越高,土壤中的盐类离子越多,对植物体生长的促进作用越强。本研究中,在生长旺盛期的薰衣草,枝条叶片与电导率呈正相关,表明薰衣草生长对当地的土壤较适应。在生长末期,植物主要受土壤绝对含水量的影响[49],土壤含水量增加,降低土壤盐度,提高植被对养分的利用效率[50]。薰衣草在生长末期,与土壤绝对含水量呈正相关,并且相关性最大(见图1),有利于该阶段养分的积累和植株越冬。
植物各构件在适应环境过程中协调发展,形成相互依赖和制约的关系,反映了植物适应环境的生存策略和自然选择的作用[51-52]。分析植物生长发育期间各构件的生长状况,有助于揭示植物的生长策略,为植物的适应与进化提供依据[53]。本研究通过对分布在哈尔滨地区的薰衣草枝条叶片生长的研究表明,薰衣草对当地的生态环境具有良好的适应性,研究结果可为拓展薰衣草的引种栽培区域提供参考。
猜你喜欢
农学学报(2022年8期)2022-11-16
农业与技术(2022年16期)2022-09-01
热带农业工程(2021年5期)2021-12-06
恋爱婚姻家庭·养生版(2021年10期)2021-10-28
少儿科学周刊·少年版(2021年20期)2021-01-17
农家科技(2020年3期)2020-05-11
中国化妆品(2019年12期)2019-04-01
作文周刊·小学三年级版(2018年2期)2018-02-06
哈尔滨理工大学学报(2016年2期)2016-09-12
海外星云 (2001年15期)2001-07-02
