
趋化因子受体的结构与信号转导机制
2021-02-07 03:56刘凯雯刘志杰华甜
自然杂志 2021年1期
刘凯雯,刘志杰,华甜
上海科技大学 iHuman研究所,上海 201210
趋化因子受体(chemokine receptor)是七次跨膜G蛋白偶联受体(G protein-coupled receptor, GPCR)超家族中的重要成员,通常表达于中性粒细胞、免疫细胞、内皮细胞等细胞膜上。GPCR是最大的膜蛋白家族,天然配体包括多肽、脂类、生物胺、糖蛋白、核苷酸和离子等,由其介导的信号通路在生理和病理过程中发挥重要作用,市场中约有40%的药物是靶向GPCR的[1-2]。GPCR与配体或GPCR与配体及下游信号转导分子复合物的三维结构解析,以及基于结构设计的细胞功能实验,对于我们揭示GPCR与配体及GPCR下游信号转导的分子机制十分重要。近十年来,GPCR结构生物学技术飞速发展,人们对于趋化因子受体家族的结构与功能研究也投入了大量精力进行探索并获得了丰厚的知识。这些结构对于趋化因子受体的激活和信号转导机制研究,以及相关疾病的药物设计研发提供了非常重要的理论基础。
1 趋化因子及趋化因子受体
趋化因子(chemokine)是一种分子量为8~10 kDa的多肽,是最大的细胞因子家族,主要功能是招募血液中的单核细胞、中性粒细胞、淋巴细胞等进入特定的淋巴器官和组织以及感染发生的部位。大多数趋化因子的氨基酸序列中有4个保守的半胱氨酸(Cys),并形成1或2对保守二硫键稳定趋化因子的核心结构[3-4]。根据保守的半胱氨酸残基的间隔氨基酸个数,通常将它们分为4个亚类:CXC类、CC类、CX3C类和C类。其中CXC 类趋化因子可以根据结构功能区第一个半胱氨酸靠近蛋白的氮(N)端处有无谷氨酸-亮氨酸-精氨酸(Glu-Leu-Arg,ELR_)氨基酸特征性序列分为两类:ELR+和ELR两个亚家族[5]。趋化因子具有保守的二级结构,一般由N端loop、3个反相平行的β折叠和1个碳(C)端α螺旋组成。
当成纤维细胞、表皮细胞、内皮细胞等组织细胞及免疫细胞在受到刺激物,如生长因子、干扰素、病毒产物及细菌产物等的诱导时,可分泌出不同的趋化因子。相应的趋化因子受体受到调控后,会诱导中性粒细胞、淋巴细胞、单核细胞、成纤维细胞等向炎症部位聚集活化,并参与组织损伤修复。研究表明,在急性炎症反应过程中,趋化因子在氨基葡聚糖作用下聚集至内皮细胞表面,通过与表达相应趋化因子受体的白细胞相结合,促进内皮细胞释放大量促炎细胞因子,对炎症反应起到关键作用[6-7]。
趋化因子受体是GPCR家族的重要成员,主要介导趋化因子的信号传导,通常由300多个氨基酸组成。趋化因子受体根据其对应结合的趋化因子配体的分类也可分为4个亚类:XCR、CCR、CXCR、CX3CR。它们都具有类似的拓扑结构,即7个跨膜螺旋区,细胞外的N端、3个细胞外loop、3个细胞内loop和细胞内C端。趋化因子受体被趋化因子等配体激活后,主要通过结合胞内的G蛋白或阻遏蛋白(β-arrestin)等信号分子来进行下游信号转导[8]。趋化因子及其受体在病原体的清除、炎症反应、病原体感染、细胞及器官的发育、创伤的修复、肿瘤的形成及其转移等方面都起着重要的作用。
2 趋化因子受体的结构研究
GPCR的结构生物学研究一直充满挑战,它们在异源系统中表达产量很低,也很难从中提取纯化目的蛋白并保持其稳定性。此外,GPCR一直处于动态平衡的构象状态,很难捕捉解析某一构象状态下的结构。近年来,X-射线晶体学(X-ray crystallography)和冷冻电子显微镜(cryo-electron microscopy, Cryo-EM)技术等的发展,极大加速了GPCR的结构解析。迄今为止,研究者已经发表了27个趋化因子受体的结构,包括13种独特的结构(图1)。其中有5种趋化因子-趋化因子受体复合物晶体或电镜三维结构,包括首次解析的结合拮抗剂以及修饰过的病毒趋化因子的CXCR4[9]、结合CX3CL1的病毒趋化因子受体US28[10]、与拮抗剂5P7-CCL5结合的CCR5[11]、结合内源性配体CCL20的CCR6-Go复合物[12],以及分别与二体和单体状态内源性配体 CXCL8结合的CXCR2-Gi复合物[13]。其中前三种为非激活状态或者类似激活状态的趋化因子受体晶体结构,后两个受体的结构均为结合内源性配体的完全激活态趋化因子受体与下游G蛋白结合的复合物冷冻电镜三维结构。此外,研究者们还解析了一系列小分子拮抗剂结合的趋化因子受体结构,包括CXCR4[14]、CCR5[15]、CCR9[16]、CCR2[17]、CCR7[18]和CXCR2[13](图1)等。
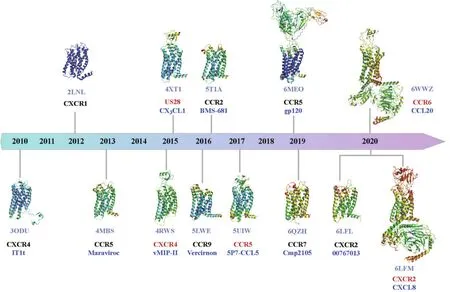
图1 目前为止已解析的独特的趋化因子受体结构(蓝灰色字体为PDB编号,亮蓝色字体表示复合物所结合的配体,红色字体表示结合趋化因子的受体结构。每个结构根据B因子进行着色,2LNL(核磁结构)除外)
2.1 趋化因子类配体与受体的相互作用
通常,趋化因子与受体的结合过程在空间和时间上称为“两步结合模型”[19],第一步主要是趋化因子受体的N端对趋化因子的招募识别并结合其球状核心,第二步主要是趋化因子的N末端插入到受体的跨膜区域口袋处进而激活受体。两步结合模型的位点一般称为趋化因子识别位点(chemokine recognition site, CRS)1和2[20]。研究表明,趋化因子的N端信号传导域在趋化因子受体激活中起着重要作用[19-21]。在趋化因子被招募初期,受体N端初始区域通过表面静电吸附等作用力将趋化因子吸引招募过来,受体N端靠近跨膜区TM1的残基与趋化因子的球状核心(N loop和β折叠)形成静电和疏水相互作用,将趋化因子稳定在受体跨膜区附近。随后,趋化因子的N末端逐渐靠近结合受体正构口袋,与跨膜区螺旋和胞外loop区形成相互作用,引起受体构象变化,从而激活下游信号。结合过程中,CRS1主要负责配体亲和性与识别功能,CRS2则主要与受体激活和信号传导功能相关联。此外,还有提出介于CRS1和CRS2之间起承接关系的CRS1.5的位点,包括趋化因子N端中间保守半胱氨酸残基与受体N端根部附近[9]。通常在CRS1.5处,趋化因子受体N端的Pro-Cys(PC)保守残基中CysN-term与受体的第三个细胞外环上CysECL3形成二硫键,有助于CRS1相互作用的稳定,起到弯折和定位作用[22]。近年来的趋化因子-趋化因子受体结构解析,揭示了趋化因子与受体广泛的结合界面,该结合界面在空间和时间上是连续不可孤立的。
趋化因子受体的正构结合口袋可以分为主要和次要亚腔[23],主要亚腔由靠近TM4(transmembrane helix 4)、TM5和TM6的口袋空腔组成,次要口袋指靠近TM1、TM2一侧的空腔口袋,TM1和TM7则介于两个亚腔之间(图2)。通过结构的对比分析发现,vMIP-II、CX3CL1或5P7-CCL5的N端主要在较深的次要口袋或同时在主、次要口袋亚腔中形成广泛的相互作用。在CXCR2-CXCL8-Gi复合物的结构中,CXCL8在CRS2中的结合位点和特异性相互作用,与已解析的CXCR4-vMIP-II、US28-CX3CL1或CCR5-5P7-CCL5晶体结构存在较大差异。CXCL8的N末端在受体结构中的结合位点口袋较浅,只占据主要口袋亚腔一侧,与ECL2形成氢键和疏水相互作用,并与TM5和TM6的靠近胞外区精氨酸残基(R2786.62、R2085.35和R2125.39)形成盐桥,在激活过程中起着关键作用(图3(a))。同时,CXCL8中的30s loop区域上的“GP”残基也与CXCR2的ECL2有较好的疏水相互作用,稳定了ECL区域构象。此外,CXCR2口袋中TM2和TM4上的极性残基Lys2.64和Arg4.64的大侧链可能会阻止CXCL8的N端更深地进入子口袋,使得CXCL8在浅层口袋结合,而Lys2.64和Arg4.64残基只在CXCR2和CXCR1中存在,而在其他趋化因子受体中相应位置主要为保守的短侧链的Ala2.64和Ser4.64。在CCR6-CCL20-Go结构中,趋化因子CCL20的N末端较短(NH2-ASNFD),结合口袋与CXCL8类似甚至更加浅,十分靠近胞外区域,并同时占据主要和次要亚腔口袋。CCL20的N端主要与受体胞外的ECL区域和N末端形成氢键等相互作用,并和受体ECL2上的可能的激活关键残基E19845.51形成盐桥,与TM1上的R421.28、TM7上的K2987.35形成盐键等相互作用。
此外,CXCL8的结合模式与CXCR4中的vMIP-II的总体架构更为相似(图2),表明该CXC趋化因子亚家族可能具有相似的结合模式。由于CXCR4-vMIP-II和CCR5-5P7-CCL5处于非激活的状态,其跨膜区与CXCR2激活态差异较大,而CX3CL1结合的US28处于近似激活的状态,因此受体结构上与CXCR2激活态更为相似。内源性配体CXCL8和CCL20结合的均是完全激活态的相应受体,它们与受体的作用均属于浅口袋结合模式。
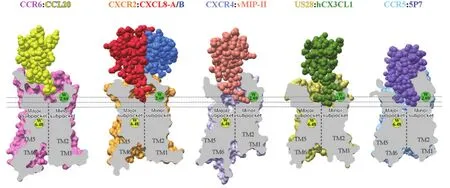
图2 趋化因子-趋化因子受体复合物相互作用口袋横截面示意图(粉色表示CCR6,亮黄色表示CCL20,橙色表示CXCR2,红色和蓝色分别表示二体CXCL8中的两个分子,淡紫色表示CXCR4,三文鱼色表示vMIP-II,浅黄色表示US28,绿色表示hCXCL1,浅蓝色表示CCR5,深紫色表示5P7-CCL5)
2.2 趋化因子受体与G蛋白的相互作用
趋化因子受体家族成员主要通过Gi/o蛋白传递信号,在CXCR2-CXCL8-Gi和CCR6-CCL20-Go复合物结构中,受体主要与G蛋白α亚基相互作用介导信号传递。相互作用界面由受体的TM3、TM5、TM6、ICL2和ICL3以及Gα亚基的α5螺旋、αN螺旋和αN-β1环、β2-β3环组成,α5螺旋结合在受体7TM近胞内区形成的口袋中。主要的相互作用与其他已解析的GPCR-Gi复合物结构相似[24-30]。在CXCL8-CXCR2-Gi结构中,Gαi的α5螺旋的两个大疏水侧链的氨基酸L353和L348指向CXCR2靠近胞内区的疏水口袋。该疏水区域主要由TM3、TM5、TM6和TM7的近胞质末端的疏水残基组成。由于CXCR2的ICL3相对较短,其上的疏水残基与Gαi的α5螺旋残基具有较好的疏水相互作用, 并与α5螺旋的部分侧链形成氢键。相比较而言,在GPCR-Gs的复合物结构中ICL2保守的F/L34.51与Gαs形成疏水相互作用。但在大多数趋化因子受体中,相应位置侧链较短的V/T34.51减弱了这种相互作用。ICL2的氨基酸残基的差异可能有助于分析趋化因子受体对下游 G蛋白的选择性。在CCR6-CCL20-Go结构中,受体上的ICL2形成两圈α螺旋结构,其上的苯丙氨酸侧链也与G蛋白loop区和α5的侧链形成较好的疏水相互作用,通常的34.51保守位点仍然为趋化因子受体家族保守的小侧链的氨基酸V34.51。此外,当将CXCR2-CXCL8-Gi复合物结构与β2AR-Gs结构[31]进行比较时,主要区别在于Gαi和Gαs的α5螺旋的相对位置,以及Gαs偶联受体中TM6的相应位置的偏移。与TM5和TM6相互作用的Gαs的α5螺旋中的C末端残基比Gi更大,因此胞质结合腔中TM5、TM6的开合大小会发生相应改变来容纳不同侧链大小的α5螺旋。总体来说,研究者们已经在结构上阐明了结合内源性趋化因子的激活态受体和下游G蛋白之间的相互作用界面,以及G蛋白偶联至趋化因子受体的分子基础。
3 趋化因子受体激活机制分析
最近CXCR2-CXCL8-Gi和CCR6-CCL20-Go复合物结构的解析为我们打开了探索趋化因子激活机制的大门。由于研究者同时也解析了CXCR2结合别构拮抗剂的非激活态结构,我们主要以CXCR2为例来阐明趋化因子受体家族的激活机制。
白细胞介素8 (CXCL8, Interleukin-8或IL-8)属于CXC类趋化因子家族ELR+亚家族成员,其N末端具有ELR特征序列[32]。在高浓度状态下CXCL8以同源二聚体形式存在,而低浓度下以单体形式存在。研究表明,单体和二聚体状态的CXCL8都能够识别并与CXCR2结合,激活下游信号并参与人体复杂的炎症反应调控[33]。因此CXCL8/CXCR2是抗癌抗炎的重要药物靶标。
在已解析的二聚体CXCL8结合的CXCR2-Gi复合物结构中,将二聚体CXCL8中两个蛋白分子定义为CXCL8-A和CXCL8-B。CXCL8-A分子与CXCR2受体正构口袋结合并激活下游信号,与单体状态的CXCL8结合机制几乎相同,另一侧的CXCL8-B与CXCR2只有较弱的相互作用。研究者通过比较CXCR2的激活态与非激活态发现,CXCL8中的N末端ELR基序在受体激活过程中起着十分关键的作用:当受体识别并结合CXCL8后,在趋化因子正构结合口袋处,CXCL8的N末端氨基酸R6和L5与受体建立的氢键和疏水相互作用分别起锚定点作用,以稳定CXCL8的N端。CXCR2的Y197ECL2与CXCL8的L5和E4的羰基氧形成氢键,以诱导受体ECL2的构象变化并促进激活。在协同作用下,主要亚腔口袋中的E4与R2085.35、R2125.39和R2786.62分别形成关键的盐键相互作用,引起TM5胞外部分向内大幅度摆动,成为CXCR2受体激活的主要驱动力(图3(a、 c))。
CXCL8引起TM5的构象变化后,A类GPCR家族典型的激活特征之一——P5.50I3.40F6.44基序中,P2235.50发生旋转,I1343.40和F2606.44发生协同重排(图3(b)),并伴随经典开关残基W2646.48的侧链构象变化(图3(b))。随后TM6螺旋靠近胞内区的部分向外大幅摆动,TM7则向跨膜区中心靠近TM3和TM5的方向移动,通过氢键等形成螺旋之间的相互作用。此外,CXCR2的激活过程中,还表现出A类家族受体激活的其他特征,包括经典的NPxxY(N3107.49、P3117.50和Y3147.53)和DRY(D1433.49、R1443.50和Y1453.51)保守结构域的构象变化。
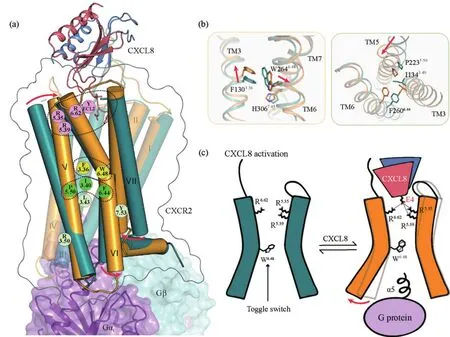
图3 CXCR2-CXCL8-Gi为例展示的趋化因子受体家族激活机制:(a)CXCR2激活态结构展示图;(b)关键特征位点激活示意图;(c)CXCR2受体激活模型图
与其他已解析的与小分子或多肽结合的GPCR结构不同,趋化因子受体的激活触发位点相对较浅,靠近受体的胞外段。这主要是由于趋化因子刺激受体相应口袋关键氨基酸,引起受体TM5的构象变化,进而导致关键的螺旋间相互作用被破坏,如TM3和TM6发生旋转,并影响受体的近胞内端跨膜区和loop区域的构象,从而使得G蛋白α亚基能够结合到胞内区,引起下游信号传递(图3(c))。
4 总结和展望
在过去的几十年里,趋化因子及趋化因子受体因在人体细胞迁移和抵御病原体入侵中起到的重要作用而引起了人们的广泛关注。除了它们在HIV病理机制方面的作用外,人们也更多地认识到它们在自身免疫及炎症、癌症方面的重要影响。趋化因子网络可以说是免疫系统的主要管理调控者。由于趋化因子及其受体功能多样,相互作用错综复杂,一直以来的结构信息不完善,科学家们对趋化因子及其受体的功能机制研究和相关的药物研发受到制约。近年来,随着多个趋化因子受体结构的不断解析,尤其是通过冷冻电镜技术解析的同时结合内源性配体和下游信号分子G蛋白的趋化因子受体复合物三维结构,展示了内源性趋化因子独特的浅口袋结合模式,以及受体与下游G蛋白的相互作用分子机制,使我们对内源性蛋白分子的识别、趋化因子信号转导和激活机制有了更深入全面的理解,也为探索趋化因子受体家族相关多肽、抗体类药物设计研发提供了指导方向。然而,趋化因子与其受体具有广泛复杂的细胞来源以及生物学效应,目前仍有大量未知的过程机制和待解决问题,需要科学家结合不同的研究方法,在分子、细胞、组织、器官和动物等不同尺度深入探索。
猜你喜欢
生物化学与生物物理进展(2022年6期)2022-07-21
中学生数理化(高中版.高考理化)(2021年2期)2021-03-19
中成药(2018年7期)2018-08-04
中成药(2018年3期)2018-05-07
中成药(2017年5期)2017-06-13
池州学院学报(2015年3期)2016-01-05
中国医药生物技术(2015年4期)2015-12-26
天津科技大学学报(2015年2期)2015-08-09
现代检验医学杂志(2015年4期)2015-02-06
现代检验医学杂志(2015年6期)2015-02-06
