
棉花种子活力与其植株停留期间气象因子的关系研究
2021-02-05 09:38:50徐敏李憬霖叶福民朱鹤金路路单莹王子胜
棉花学报 2021年1期
徐敏,李憬霖,叶福民,朱鹤,金路路,单莹,王子胜*
(1.辽宁省经济作物研究所,辽宁 辽阳111000;2.中国农科院棉花研究所,河南 安阳455000)
1976 年,国际种子检验协会(ISTA)将种子发芽和出苗期间活性的综合表现,定义为种子活力[1],并以之作为衡量种子质量的主要指标。在种子萌发过程中, 会发生一系列生理生化变化,如信号和激素物质被激活, 种子的休眠被打破,大分子物质的修复[2],氧化还原酶(Peroxidase,POD,过氧化物酶;superoxide dismutase, SOD 超氧化物化酶;catalase,CAT,过氧化氢酶等)被激活等,从而决定了种子活力。 大量的研究表明,种子活力与种子成熟度密切相关[3]。 种子所含成分的差异也会对种子活力造成一定影响[4-5]。 棉花生长发育期间的气象条件必然会影响到种子的发育。 张旺峰[6]等通过比较不同生态区棉花铃重变化与气象因子之间的关系, 认为新疆棉区日照时间长、平均温度高、日温差大,有利于光合物质的产生及在棉铃中的积累,是新疆棉区获得高产的生理基础。张丽娟等[7]、马富裕等[8]研究认为,棉花纤维比强度是对气候变化最为敏感的品质指标,而温度是其最主要的影响因素。王友华等[9]研究认为,不同棉花品种纤维比强度的稳定性存在较大差异,变异的主要来源在于其对温度变化敏感性的差异。 可见,关于棉花生长与气象条件的关系,前人更多关注棉铃发育、纤维品质等方面。 另一方面,关于种子和种子活力的研究,林业和其他经济作物的研究受到一定的重视, 而有关大田作物种子的研究较少。 有关棉花种子活力的研究更多集中在种子引发[10-14]、老化和劣变等方面[15-17]。陈莉等[13]研究认为,褪黑素能够影响棉花种子的萌发;刘瑞显等[14]研究发现,小麦秸秆的浸提液和腐解液具有极强的化感效应,能够抑制棉花种子的萌发和幼苗生长。 棉花种子作为重要的生殖器官,其生长发育的过程必然会受到外界条件的影响。 张文强等[18]研究了不同果位和结铃期对棉花种子活力的影响,认为高活力种子生产以采收中部果位的伏桃种子为宜。
那么, 气象因子是如何通过影响种子的发育,进而影响种子活力呢? 本试验通过比较不同成熟度棉花种子的特征以及萌发过程中的生理生化指标的变化,力图发现其中的规律,为生产实践提供指导。
1 材料与方法
1.1 供试棉花品种
本试验选择辽棉25、辽棉31 两个棉花品种。这两个品种都是辽宁省经济作物作物研究所选育的,遗传差异较大。 其中:辽棉25 号于2011 年审定,编号为:辽审棉[2011]27 号。 生育期122 d左右,长势较旺,植株塔形、松散,叶片较大,叶色深绿;铃卵圆形,较大。种子较大。衣分40.20%。抗病性强,高抗枯萎病,抗黄萎病。 棉絮洁白,纤维品质优良。 辽棉31 号于2014 审定,编号:辽审棉2014002。 生育期130 d 左右;长势中等,植株塔形、较松散;叶片较小,叶色较深;铃卵圆形,较大。种子大小中等。衣分40.98%。抗病性强,抗枯萎病,抗黄萎病。 棉絮乳白色,纤维品质优良。
1.2 不同植株停留时间棉花种子群体的获得
2016 年, 在辽宁省经济作物研究所试验地,于棉花花期(7 月12 日、7 月19 日、7 月26 日、8月2 日、8 月9 日)对当日所开花挂牌标记。 在棉花吐絮期(9 月19 日、9 月29 日、10 月12 日)分别收获。 将所收获的棉花分别轧花收取种子,构成不同成熟度的棉花种子群体。 两个品种共包含13份样品。 详见表1。
1.3 棉花种子特征指标的测定
从不同成熟度棉花种子群体中取适量种子,硫酸脱绒后,检测百粒重,并进行发芽试验。
1.3.1 种子百粒重测定。 选择大小均匀、无破损的种子,测量百粒种子质量,每样品重复3 次,取平均值为种子百粒重。
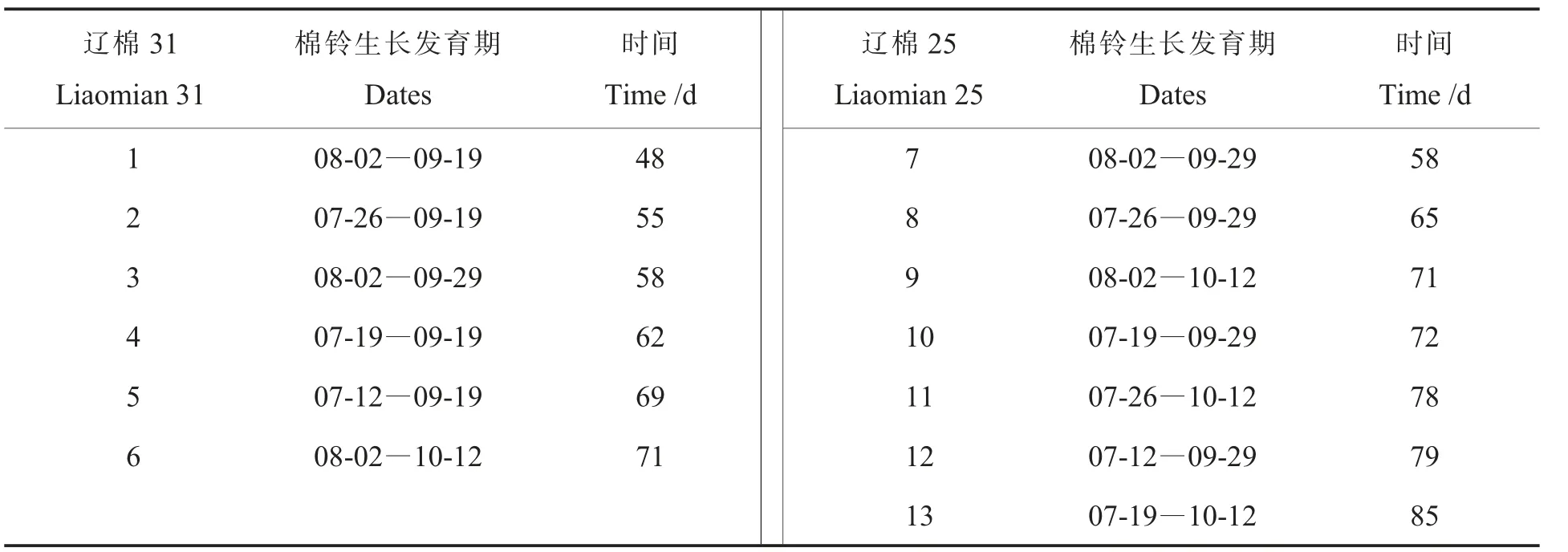
表1 不同成熟度棉花种子群体Table 1 Cotton seed population with different maturity levels
1.3.2 种子发芽试验。 每样品取100 粒种子,使用发芽盒,在温度28℃、湿度40%的条件下,利用培养箱进行发芽试验。 在第7 天时计算种子发芽率,3 次重复,取平均值。
1.4 棉花种子营养成分含量的测定
利用索氏提取法测定种子的粗脂肪含量,所用仪器为FOSS Soxtec 8000 索氏提取仪。 用蒽酮显色法测定种子的可溶性糖含量和淀粉含量[19]。
1.5 棉花种子萌发期间酶水平的测定
各样品取10 粒种子(约1 g),使用发芽盒在在温度28 ℃、湿度40%的条件下,利用培养箱进行发芽试验。 分别在发芽第1 天、第4 天、第7 天测定种子丙二醛 (MDA) 浓度及过氧化物酶(POD)、过氧化氢酶(CAT)和超氧化物歧化酶(SOD)活性。 所有指标的测定均重复3 次,最终结果取平均值。 具体检测方法:采用TBA 比色法测定种子丙二醛(MDA)浓度,采用愈创木酚法测定种子过氧化物酶(POD)活性,采用紫外吸收法测定种子过氧化氢酶(CAT)和超氧化物歧化酶(SOD)活性[20]。
1.6 气象数据收集整理
本文所有气象数据皆采自中国气象数据网。
1.7 数据整理与分析
利用MS Excel 2007 软件进行数据的初步整理和作图,利用DPS 软件进行分析和比较。
2 结果与分析
2.1 种子植株停留期间气象因子汇总
经过检索中国气象数据网, 选择了降水量X1、相对湿度X2、平均气温X3、≥12 ℃积 温X4、日最高温X5、日最低温X6、日温差X7和日照时间X8共8 个因子。 根据各样本的开花日期和植株停留时间,对各指标分别取值。 如表2。
由表2 可见, 随着种子植株停留时间的增加,降水量、日照时间和≥12℃积温3 个累计指标和日温差共4 个指标呈现不稳定的上升趋势,与预期相符。 相对湿度以及日均气温、日最高气温、日最低气温4 个平均指标与种子的挂牌时间表现一定的同步趋势,即:挂牌时间越晚,各指标值越低。 总体来看:第一次挂牌时(7 月12 日)正值伏季, 为一年中高温高湿季节;7 月底开始,温度下降、降水减少,同时日照增加、日温差加大,综合气象条件有利于植株的干物质积累和成熟。本试验正是基于这个出发点而设计和进行的。
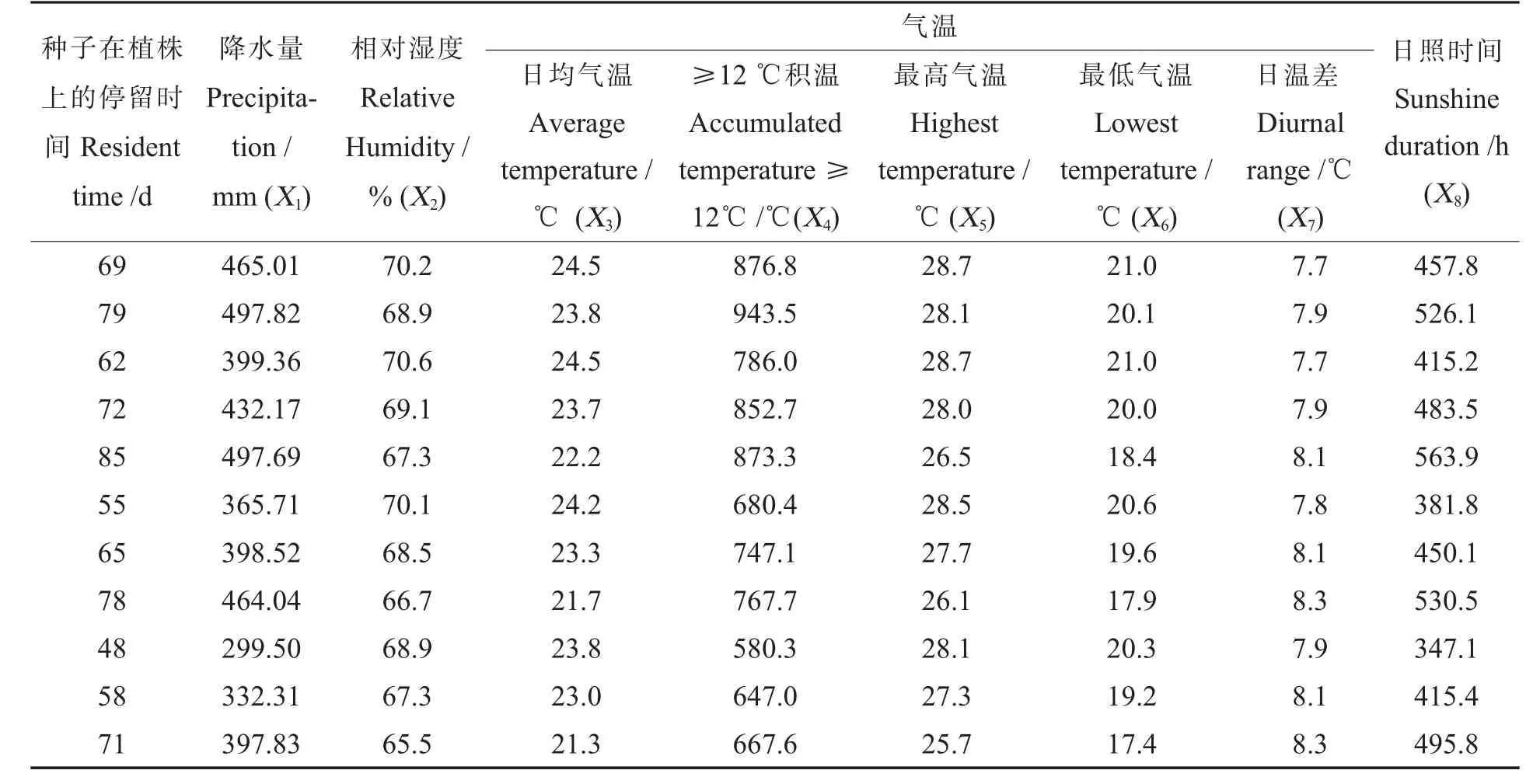
表2 种子植株停留期间的主要气象因子Table 2 Main meteorological factors during seed resident on the plants
2.2 种子百粒重和发芽率
由表3 可见:样本间百粒重差异显著,并且与种子的植株停留时间显著负相关(r=-0.584 9,α=0.05 时相关系数临界值r=0.552 9): 种子的植株停留时间在50~70 d 之间时, 百粒重可达10 g 以上;低于50 d 或高于70 d 都会下降。 各样本种子的7 d 发芽率差异较小, 与种子的植株停留时间无显著相关。 无论是百粒重还是7 d 发芽率,辽棉31(10.01 g,82.28%)都稍高于辽棉25(9.79 g,80.19%)。
2.3 种子主要营养成分含量
由图1 可见, 棉花种子中粗脂肪含量最高,达到20%左右,因此,棉花种子被归类为脂肪类种子[21];其次是可溶性糖,5%左右;淀粉含量最低,3%左右。总体看来,样本间粗脂肪含量差别较大,淀粉含量和可溶性糖含量差别较小。辽棉31的粗脂肪含量(19.56%)高于辽棉25 号(18.42%),两个品种间差异显著(P=0.001 6);辽棉31 的可溶性糖含量和淀粉含量(5.14%,2.99%)稍低于辽棉25(5.32%,3.33%),两个品种间差异不显著。各营养成分中,只有可溶性糖含量与种子的植株停留时间表现显著相关(r=0.650 1,α=0.05 时相关系数临界值r=0.552 9)。
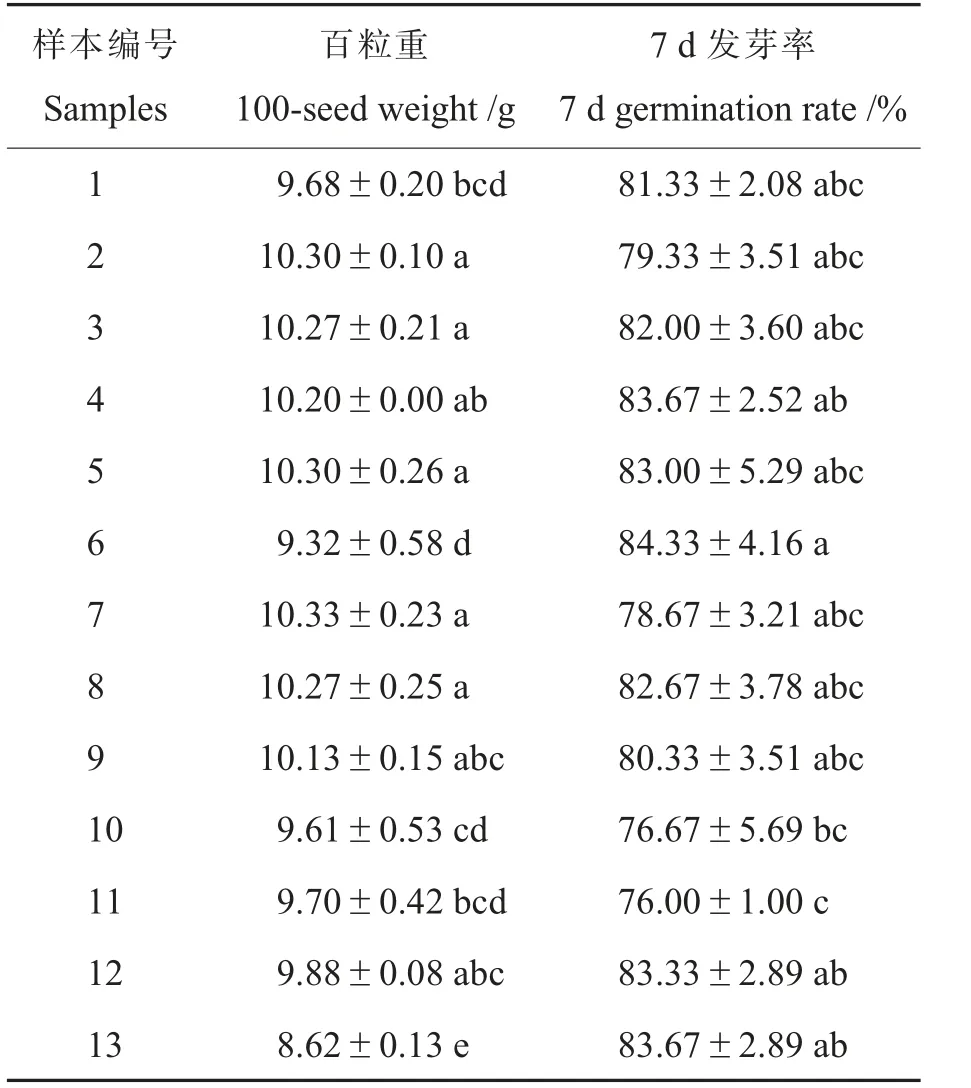
表3 种子百粒重和发芽率统计表Table 3 100-seed weight and germination rate
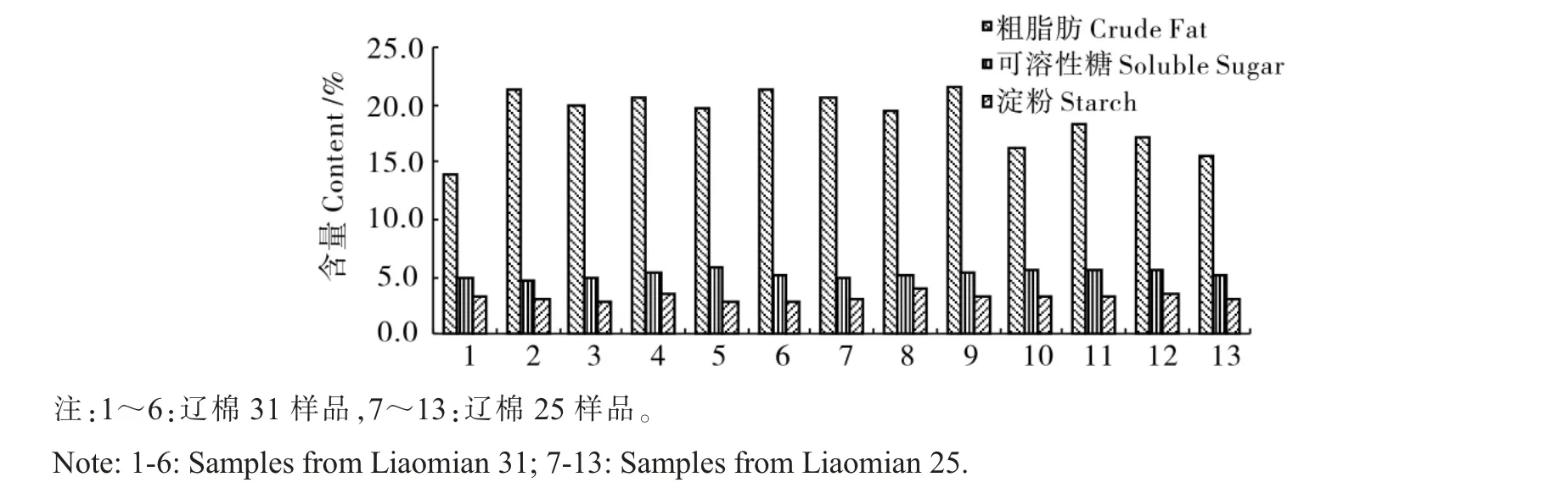
图1 种子主要成分含量Fig. 1 Content of main nutrients in seeds
2.4 种子发芽期间丙二醛(MDA)浓度变化情况
由图2 可见, 种子发芽期间MDA 浓度呈先升高后下降趋势。 发芽1 d 时,种子的生理活性较小,MDA 浓度较低(7.57 μmol·g-1)且样品间差异较大。发芽4 d 时,种子被充分激活,进入萌发旺盛期,MDA 浓度上升明显(21.35 μmol·g-1)且样品间差异缩小。 发芽7 d 时,已经到发芽末期, 种胚的活性降低,MDA 浓度也随之降低(18.19 μmol·g-1),样品间差距继续缩小。 总体来看,辽棉25 的MDA 浓度(9.79 μmol·g-1)低于辽棉31(10.01 μmol·g-1),两个品种间差异不显著。
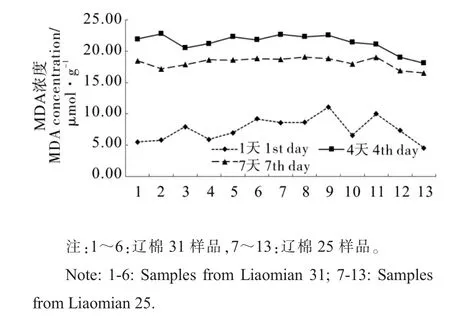
图2 MDA 浓度变化情况Fig. 2 Change of MDA concentration
2.5 种子发芽期间酶活性变化情况
2.5.1 过氧化物酶(POD)活性变化情况。 由图3可见, 种子发芽期间POD 活性呈先升高后下降趋势,下降较明显,且样品间差别明显。 发芽1 d时, 平均POD 活性为0.055 △OD·(min·g)-1;4 d时为0.109 △OD·(min·g)-1,几乎是发芽1 d 时的2 倍;7 d 时迅速下降为0.046 △OD·(min·g)-1,比1 d 时还要低。 总体来看, 辽棉25 的POD 活性(0.13 △OD·(min·g)-1) 高于辽棉31 (0.08 △OD·(min·g)-1),但两个品种间差异不显著。
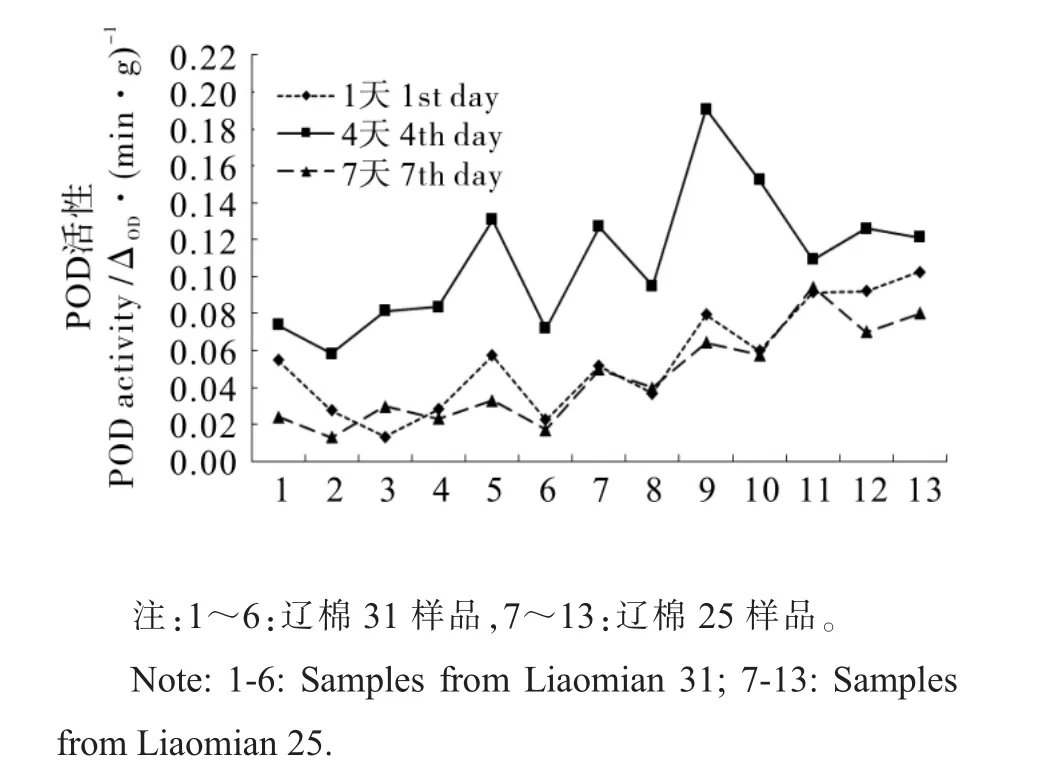
图3 POD 活性变化情况Fig. 3 Changes of POD activity
2.5.2 过氧化氢酶(CAT)活性变化情况。 由图4可见, 种子发芽期间CAT 活性呈先下降再升高趋势,下降明显,但升高有限;且样品间差别明显。 发芽1 d 时,CAT 活性(以蛋白质含量计)为663.40 μmol·(min·mg)-1;4 d 时为375.31 μmol·(min·mg)-1,几乎下降了50%;7 d 时微有上升,达到439.21 μmol·(min·mg)-1。 辽棉25 的CAT活性(437.96 μmol·(min·mg)-1)稍高于辽棉31(302.22 μmol·(min·mg)-1) ,但两个品种间差异不显著。
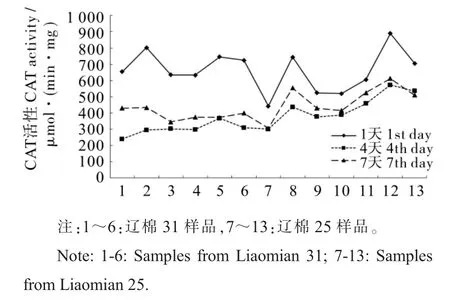
图4 CAT 活性变化情况Fig. 4 Changes of CAT activity
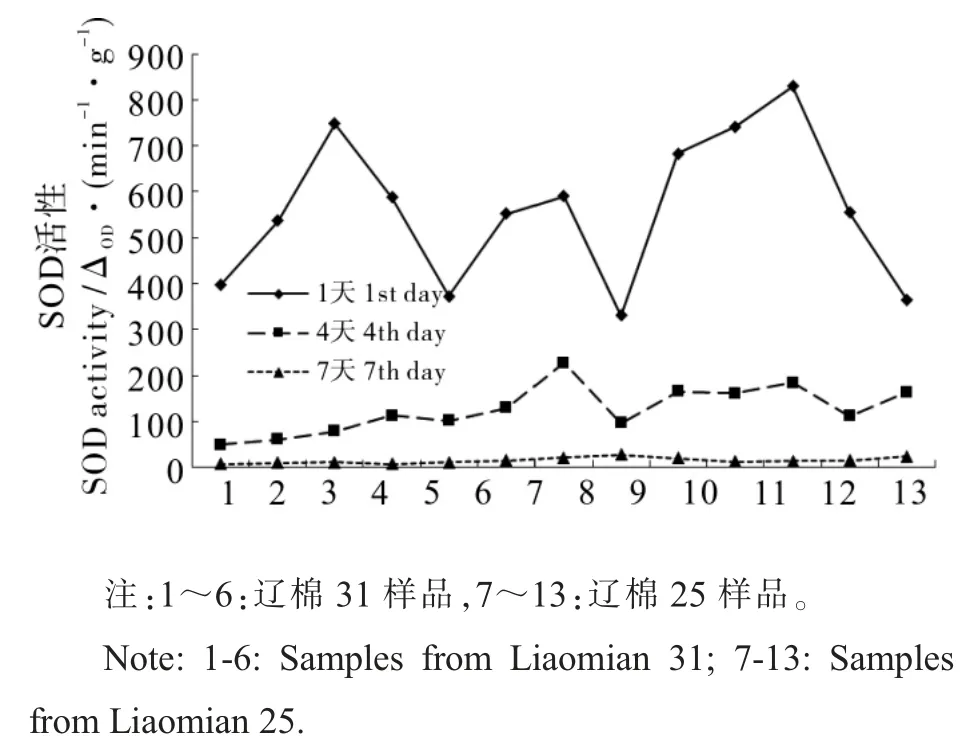
图5 SOD 活性变化情况Fig. 5 Changes of SOD activity
2.5.3 超氧化物歧化酶(SOD)活性变化情况。 由图5 可见,种子发芽期间SOD 活性呈明显下降趋势, 且样本差别较小。 发芽1 d 时,SOD 活性为560.49 △OD·(min·g)-1,4 d 时 为126.30 △OD·(min·g)-1,7 d 时为14.94 △OD·(min·g)-1,辽棉25的SOD 活性158.51 △OD·(min·g)-1高于辽棉31 的88.71 △OD·(min·g)-1,但两个品种间差异不显著。
2.6 棉花种子基本特征与发芽期间生理生化指标与气象因素的相关分析
根据徐敏等[21]研究的结果,种子发芽4 d 时为发芽盛期,其各项指标更具有代表性,因此,以发芽4 d 时数据为基础, 计算种子的特征指标和气象因子之间的相关关系(表4)。
由表4 可见:可溶性糖含量与降水量和日温差呈现极显著正相关, 与≥12 ℃积温和日照时间显著正相关。MDA 含量与降水量、≥12 ℃积温、日温差和日照时间呈现显著负相关。 CAT 活性与降水量、≥12 ℃积温、日温差和日照时间呈现极显著正相关。 此外,百粒重与日照时间呈现显著负相关。
由此可见,从气象因子方面来说,降水量、≥12 ℃积温、日温差和日照时间与种子的部分生理生化指标显著或极显著相关;从种子的生理生化指标来说, 可溶性糖含量、MDA 浓度和CAT 活性与部分气象因子的相关达到显著或极显著水平。 因此,以下主要围绕百粒重、可溶性糖含量、MDA 浓度和CAT 活性4 个种子的特征指标和降水量、≥12℃积温、 日温差和日照时间4 个气象因子进行分析。
2.7 棉花种子特征指标与气象因子的数学模型模拟
在以上分析的基础上,将相关显著或极显著的指标进行二次多项式逐步回归分析,以气象因子为自变量, 以棉花的生理生化特征值为因变量,建立模拟方程。 由于本试验选用2 个品种,因此,选用两品种平均值及辽棉31、辽棉25 各自的数据分别进行模拟, 其中25 个模型模拟成功(r2≥0.5,P≤0.05),详见表5。
所有模拟成功的25 个模型中, 所表现的趋势与相关分析的结果基本相符, 选择其中符合y=ax2+bx+c 的二次曲线的方程, 进一步利用边缘分析方法考察气象因子对各指标的边际效应(表6)。
边际效应值反映了种子基本指标对气象因子的反应特征。 表中所有指标中, 除CAT 活性与≥12 ℃积温、日照时间的边际效应与预期相差较大外,其余指标基本符合预期。参考张旺峰等[6]的研究,从逐步回归模型和边际效应的数学模型可以看出,气象因子对棉花种子特性影响的特定特征值都有一个最适值(Optimum value,OV),也就是所说的边际效应。 当OV=xi=b/2a 时,y'=2axi-b=0,即是气象因子的最适值。 当OV<xi时,由于y' 变化率不同,表现出不同的趋势:y'>0,边际效应值为正,即随着各气象因子数值的提高, 种子的特征值会上升;y'<0, 边际效应值为负,即随着各气象因子数值的提高,种子的特征值则会下降。当OV>xi时,情况刚好相反。同时,y' 变化率绝对值也能代表y' 值的稳定程度,即y'值对x 值的利用效率,变化率越大则y'值稳定程度越低。 由表中可见,降水量的波动对种子各指标的影响不明显,日照时间波动对各指标的影响稍大,而日温差的波动对各指标的影响最大。
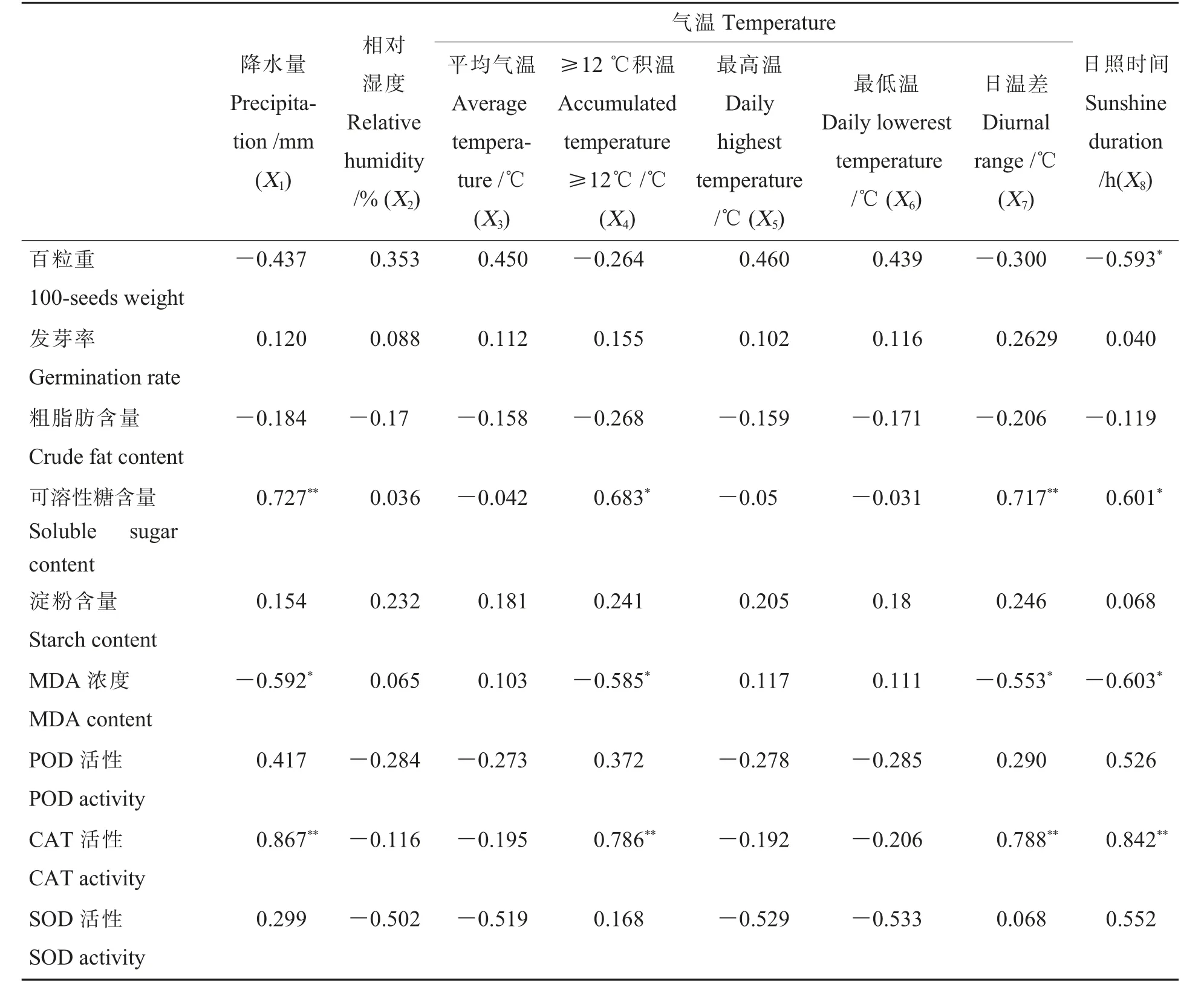
表4 棉花种子特征指标与气象因素的相关系数Table 4 Correlation coefficients among samples
所选择的棉花种子特征值并非越大或者越小越好, 同时各指标之间还有一定的互作效应。综合各指标的边际效应认为,降水量的最适值为305~480 mm,日温差的最适值7.3~7.83 h,日照时间的最适值为421~508 h。 由此推断种子在植株的最佳停留时间为65~72 d。
3 讨论
3.1 棉花种子形成、发育和萌发过程中各项指标互相影响
棉花种子的形成、发育和萌发是一个完整的生理过程,尤其是萌发出苗的过程,受到外部条件和内在因素的共同作用。 仅以本试验所选择的4 个指标来说,同种作物种子,百粒重大代表种子成熟度好,储藏丰富,有利于种子萌发;可溶性糖作为能量物质,一般在幼苗能够自主光合作用以前,为种子萌发和出苗提供能量;MDA 是种子萌发期间膜脂过氧化的产物, 但作为有害物质,种子会通过自身的生理作用对其进行分解以消除影响;CAT 是一种活跃的过氧化物酶, 是种子自身保护系统的重要组成成分。 综合来看,MDA 含量增加必然会引起CAT 活性的提高,CAT 的活动又会消耗更多的能量,进而影响其他的生理过程。 因此,所有的指标并非简单的升高或下降就有利于提高棉花种子的活力。
3.2 MDA 来自于膜脂过氧化, 环境胁迫并非必要条件
有观点认为,MDA 的形成是由于环境胁迫造成的,在环境条件良好的情况下,种子能够顺利萌发,不会形成MDA 的积累。 也有研究认为,MDA 来源于膜脂的过氧化, 其产生是种子发芽过程中不可避免的生理活动[4]。 本试验的结果更倾向于第二种观点。 另一方面,有关种子的试验研究一般采用人工发芽的形式进行,提供理论上最适的温度和湿度条件,因此认为种子是在最适条件下萌发。 但人工环境与自然环境存在差异,即使提供了理论最适的条件,种子仍然需要通过一系列自我调节,能够顺利萌发,这一点需要重新加以认识。
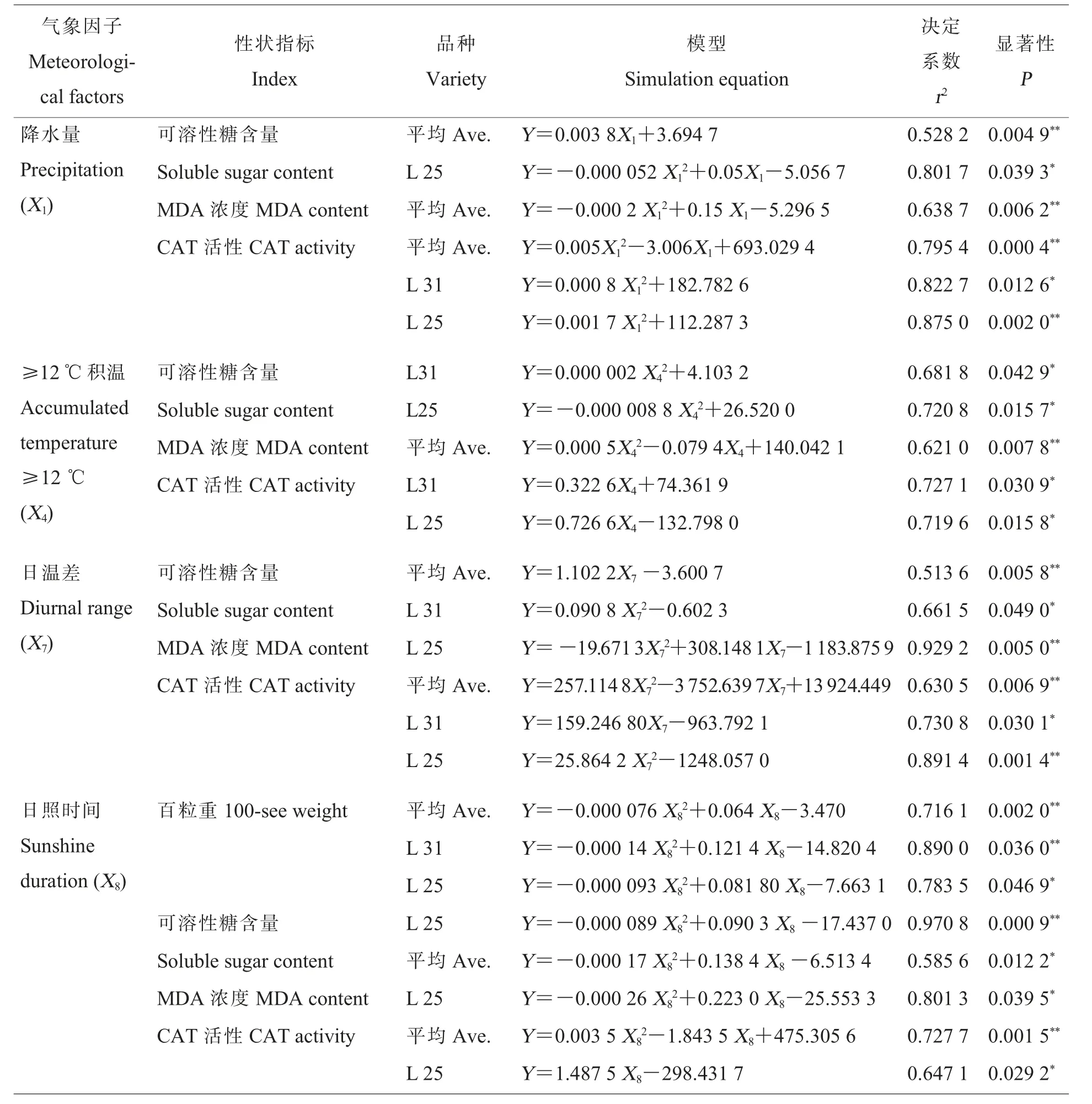
表5 种子生理生化指标与气象因子的二次多项式逐步回归模型Table 5 Quadratic polynomial stepwise regression models of seed characteristic indices and meteorological factors
3.3 种子自身的抗氧化系统能够起到自我调节和保护的能力
根据El-Maarouf-Bouteau 等[22]的研究,在种子萌发过程中,活性氧(ROS)作为信号分子参与调控种子的休眠释放、胚乳的松弛和储存物质的动员。 但是,活性氧的过量积累又会起到毒害作用,抑制种子的萌发。随着O2-自由基的积累,种子自身具有的抗氧化系统将被激活,以清除O2-自由基,起到自我保护的作用[5]。 本试验所检测的过氧化物酶(POD)、过氧化氢酶CAT、超氧化物歧化酶(SOD)就是种子抗氧化系统的重要组成成分。

表6 种子特征指标与气象因子的边际效应模型Table 6 Marginal effect model of seed characteristic indices and meteorological factors
本试验中,POD、CAT 活性都在发芽4 d 时达到顶峰,此时也是发芽的高峰,种子的生理活动活跃,O2-自由基产生较多。 1 d 时种子刚刚萌动、7 d 时种子发芽活动接近尾声, 所产生的O2-自由基浓度较低,酶活性也相应较低,以减少能量的消耗。总之,种子发芽期间O2-自由基含量变化会直接影响酶活性的变化,并且与发芽的趋势基本一致,这一点与预期相符。
3.4 部分气象因子通过影响种子部分特征指标进而影响种子活力
棉花起源于热带和温带地区,属喜温无限生长类型植物[9]。随着棉花种植范围的扩大,气象因子对棉花生长发育及产量形成的影响逐渐成为人们研究的热点,棉花生产标准化、机械化水平的提高,更加重了对这方面研究的需求。
本试验最初选择了8 个气象因子,其中相对湿度、日最高温、日最低温和日均气温4 个指标由于与种子的特征指标相关不显著而剔除,而降水量、≥12 ℃积温、 日温差和日照时间4 个气象因子通过影响种子的百粒重、 可溶性糖含量、MDA 浓度以及CAT 活性, 进而影响种子活力。随着4 个气象因子指标的上升, 可溶性糖含量、CAT 活性随之上升, 而MDA 浓度呈下降趋势。可见,充足的日照、降水、积温及较大的日温差有利于种子发育,营养物质积累增加,酶活性提高,促使种子顺利萌发。 边际效应分析发现日温差和日照时间的变化更容易引起可溶性糖含量和CAT 活性的变化,即其对种子的发育和成熟影响较明显,这一点与预期相符。
3.5 关于种子后熟对种子活力影响的研究
后熟作用是指成熟种子离开母体后,经过一系列的生理生化变化,完成生理成熟而具备发芽能力的生理过程。 后熟期长短因植物种类而异,同时,外部条件对种子的后熟有一定的影响。 有研究表明,棉花种子有很明显的后熟过程,陆地棉的棉籽在棉铃刚吐絮时就有发芽能力,但发芽率很低,仅有14%~18%;经晒干、储藏2~4 个月,棉籽完成后熟作用,能够显著提高发芽率[23]。关于棉花种子的后熟,前人进行了一些研究。 李艳萍等[24]研究了棉花种子后熟过程中活力变化的规律,认为棉花种子后熟期主要决定于种子干燥脱水的程度。 张晓洁等[25]的研究结果与之相似。
本试验仅比较了种子在植株上停留期间气象条件对其活性的影响。 试验所需种子皆于棉铃正常吐絮后收获,于当年10 月底轧花,后即入库统一保存。 第二年4 月初统一硫酸脱绒,6~7 月统一进行发芽试验和检测。 关于后熟对种子活力的影响,将另外安排试验。
4 结论
本试验通过综合比较各气象因子的边际效应,认为棉花种子在辽河流域的最适植株停留时间为65~70 d,也就是开花时间为7 月中旬。 该类种子大部分来自早伏桃以及部分伏前桃。
猜你喜欢
今日农业(2022年6期)2022-07-05 15:13:21
作文周刊·小学一年级版(2022年24期)2022-06-18 13:11:03
内蒙古气象(2021年2期)2021-07-01 06:19:58
科学导报(2020年8期)2020-03-12 04:51:04
心声歌刊(2019年5期)2020-01-19 01:52:52
中国食品学报(2020年2期)2020-01-17 07:59:08
领导决策信息(2018年46期)2018-04-20 04:00:42
南方农业·下旬(2018年1期)2018-02-23 09:07:19
潍坊学院学报(2016年2期)2016-12-01 12:59:44
军事文摘·科学少年(2015年6期)2015-05-30 20:39:04
