
Skp2通过调控Snaill水平对上皮性卵巢癌细胞EMT的作用研究
2021-01-26 04:13徐小丽
国际检验医学杂志 2021年2期
徐小丽,李 青,陈 彬
安徽省安庆市立医院妇产科,安徽安庆 246001
卵巢癌致病机制复杂,在疾病早期难以诊断,超过70%的卵巢癌患者在晚期(Ⅲ或Ⅳ期)被发现,同时,卵巢癌细胞通常对化学治疗药物具有抗性,导致患者整体生存率急剧下降[1-2]。卵巢癌是上皮性肿瘤,具有高度转移性,与其他肿瘤细胞不同,由于没有解剖屏障,卵巢癌细胞可以直接扩散到腹膜腔,而不通过血液扩散[3]。在卵巢癌的发生和发展过程中,上皮间质转化(EMT)是其中的关键过程[4]。E-钙黏蛋白(E-cadherin)的表达抑制被认为是EMT过程中的一个基本事件,许多细胞因子如转录因子Snaill、锌指E框结合同源框1/2(ZEB1/2)和样转录因子8(KLF8)等可直接与E-cadherin启动子区结合而抑制其转录[5-6]。Snaill在EMT的调控过程中起关键作用,包括E-cadherin的水平降低,以及波形蛋白(Vimentin蛋白)和纤连蛋白(Fibronectin)等生物标志物水平的升高。S期激酶相关蛋白2(Skp2)是近年发现的一种F-box蛋白,可以直接降解Snaill,在许多晚期癌症(包括卵巢癌)中发现其低表达[7]。体外研究显示,Skp2过表达可抑制腺癌或鳞状细胞癌细胞系的迁移和侵袭[8]。同时,卵巢癌中Skp2的低表达与E-cadherin的丢失有关,表明发生了EMT[9]。然而,Skp2在EMT中的分子机制尚不清楚。因此,本研究探讨Skp2通过调控Snaill水平在抑制上皮性卵巢癌细胞EMT方面的作用,以期为卵巢癌的治疗提供新的策略。
1 材料与方法
1.1材料 分别收集15例上皮性卵巢癌组织和正常卵巢组织。所有研究对象均未接受放疗或化疗,切除后立即将标本放液氮中保存。所有标本均经病理学确诊,所有研究对象对本研究均知情同意并签署知情同意书,同时本研究获得本院医学伦理委员会批准。
1.2仪器与试剂 上皮性卵巢癌SKOV3细胞(中国科学院上海细胞研究所),胎牛血清和RPMI-1640培养基(美国Gibco公司),脂质体Lipofectamine2000(美国Invitrogen公司),Skp2干扰表达载体(5′-CCGGGATAGTGTCATGCTAAAGAATCTCGAG ATTCTTTAGCATGACACTATCTTTTTG-3′,广州锐博公司设计与合成)和Skp2过表达载体(5′-CCGGGCCTAAGCTAAATCGAGAGAACTGAGT TCTCTCGATTTAGCTTAGGCTTTTTG-3′,广州锐博公司设计与合成),Lipofectamine2000转染试剂盒(美国Invitrogen公司),TRIzol(美国Invitrogen公司),RNA反转录试剂盒,实时荧光定量聚合酶链反应(RT-PCR)试剂盒(宝生物工程有限公司)。PCR引物序列由大连TaKaRa公司设计及合成:Skp2上游引物为5′-ATAAGCTTATGCACAGGAAGCA-CCCTCCAGG-3′,下游引物为5′-AAGAATTCTCATAGACAACGGGCTTTTGC-3′;Snaill上游引物为5′-TCCAGAGTTTACCTTCCAGCA-3′,下游引物为5′-CTTTCCCACTGTCCTCATCTG-3′;E-cadherin上游引物为5′-GCTGCTCTTGCTGTTTCTTCG-3′,下游引物为5′-CCGCCTCCTTCTTCATCATAG-3′;Vimentin上游引物为5′-AAGTTTGCTGACCTCTCTGAGGCT-3′,下游引物为5′-CTTCCATT TCACGCATCTGGCGTT-3′;U6上游引物为5′-CTCGCTTCGGCAGCACA-3′,下游引物为5′-AACGCTTCACGAATTTGCGT-3′。RIPA裂解液(上海生工生物工程有限公司),Skp2、Snaill、E-cadherin、Vimentin、β-actin鼠抗和辣根过氧化物酶(HRP)标记山羊抗鼠IgG二抗(美国Santa Cruz公司),电化学发光(ECL)试剂盒(美国GE公司),二氧化碳细胞培养箱(美国Ther-mo Revco公司),24孔Transwell小室(美国Corning科技有限公司),NanoDrop2000c型蛋白核酸检测仪(美国Thermo公司),实时荧光定量PCR仪和BIO-RAD垂直电泳仪(美国BIO-RAD公司),凝胶成像仪(美国UVP公司)。
1.3方法
1.3.1细胞分组及转染 将实验对象分为阴性对照组(只加脂质体)、Skp2干扰表达组(加Skp2干扰表达载体和脂质体)、Skp2过表达组(加Skp2过表达载体和脂质体)。将含10%胎牛血清的RPMI-1640培养基在37 ℃、5%CO2环境中培养SKOV3细胞,选取对数生长期的细胞进行实验。对SKOV3细胞进行离心后接种于6孔板中,待细胞生长后更换为无血清培养液(RPMI-1640),分别用Lipofectamine法将脂质体、Skp2干扰表达载体和Skp2过表达载体转染到SKOV3细胞中,转染6 h后更换为含血清培养液(RPMI-1640)继续培养48 h收获细胞,进行相关检测。
1.3.2RT-PCR法检测组织中Skp2、Snaill mRNA,以及细胞中Skp2、Snaill、E-cadherin和Vimentin mRNA的表达 阴性对照组、Skp2干扰表达组和Skp2过表达组细胞转染成功后以5×104/孔接种于6孔板内(3毫升/孔),继续培养48 h收获细胞,使用TRIzol提取总RNA,测定mRNA的水平和纯度,根据RNA反转录试剂盒说明书将提取的总RNA反转录为cDNA,根据RT-PCR试剂盒说明书,制备20 μL反应体系。在RT-PCR仪中进行扩增,反应条件为预变性95 ℃ 30 s、变性95 ℃ 5 s、60 ℃ 44 s、40个循环,61 ℃时采集荧光,用RT-PCR仪进行检测并对其表达量进行分析。以U6作为内参,采用2-ΔΔCt法计算Skp2、Snaill、E-cadherin和Vimentin mRNA的相对表达量(正常卵巢组织和卵巢癌组织标本只进行Skp2和Snaill mRNA的检测)。每组试验重复3次。
1.3.3Transwell法检测细胞体外侵袭、迁移能力 各组细胞转染成功后以5×105/mL接种在24孔Transwell小室中(200微升/孔),下室加入含10%胎牛血清的RPMI-1640培养液,继续培养48 h,取出小室,用无菌棉签擦拭去除上室内的细胞,用4%甲醛固定10 min,用0.1%结晶紫染液染色30 min,冲洗干净后于倒置显微镜下观察细胞并拍照。计数染成紫色的穿膜细胞(代表细胞侵袭和迁移能力)。每组试验重复3次。
1.3.4免疫印迹(western blot)法检测Skp2、Snaill、E-cadherin和Vimentin蛋白表达 各组细胞转染成功后以5×104/孔接种于6孔板内(3毫升/孔),继续培养48 h收获细胞,加入RIPA裂解液,冰浴2 h。离心取上清液进行蛋白定量。调整蛋白水平,沸水进行变性。进行电泳(每孔上样量为20 μg),将膜与Skp2(1∶200)、Snaill(1∶500)、E-cadherin(1∶400)、Vimentin(1∶500)和β-actin(1∶1 000)抗体进行孵育,4 ℃过夜,将IgG二抗(1∶5 000)在室温下孵育30 min。进行显色,采集图像,通过Image LabTM软件(Bio-Rad)获取信号并估计强度,以β-actin作为内对照。每组试验重复3次。
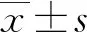
2 结 果
2.1正常卵巢组织和卵巢癌组织中Skp2和Snaill mRNA的表达情况 Skp2 mRNA在卵巢癌组织中的表达水平明显低于正常卵巢组织、Snaill mRNA在卵巢癌组织中的表达水平明显高于正常卵巢组织,差异均有统计学意义(P<0.05)。见表1。
表1 正常卵巢组织和卵巢癌组织中Skp2和 Snaill mRNA的表达情况
2.23组细胞中Skp2、Snaill、E-cadherin和Vimentin mRNA表达情况 与阴性对照组相比,Skp2干扰表达组细胞中Skp2和E-cadherin mRNA表达水平降低,Snaill和Vimentin mRNA表达水平升高,差异均有统计学意义(P<0.05);与Skp2干扰表达组相比,Skp2过表达组细胞中Skp2和E-cadherin mRNA表达水平升高,Snaill和Vimentin mRNA表达水平降低,差异均有统计学意义(P<0.05)。见表2。
2.33组细胞侵袭、迁移能力 与阴性对照组相比,Skp2干扰表达组侵袭细胞数和迁移细胞数增加,差异有统计学意义(P<0.05);与Skp2干扰表达组相比,Skp2过表达组侵袭细胞数和迁移细胞数降低,差异有统计学意义(P<0.05)。见图1~2、表3。
表2 3组细胞中Skp2、Snaill、E-cadherin和Vimentin mRNA表达情况
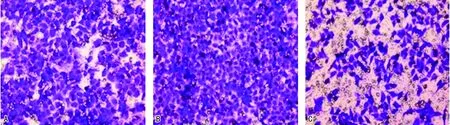
注:A为阴性对照组;B为Skp2干扰表达组;C为Skp2过表达组。
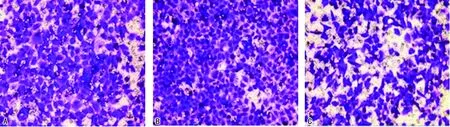
注:A为阴性对照组;B为Skp2干扰表达组;C为Skp2过表达组。
表3 3组细胞侵袭、迁移能力
2.43组细胞中Skp2、Snaill、E-cadherin和Vimentin蛋白表达情况 与阴性对照组相比,Skp2干扰表达组细胞中Skp2和E-cadherin蛋白表达水平降低,Snaill和Vimentin蛋白表达水平升高,差异有统计学意义(P<0.05);与Skp2干扰表达组相比,Skp2过表达组细胞中Skp2和E-cadherin蛋白表达水平升高,Snaill和Vimentin蛋白表达水平降低,差异有统计学意义(P<0.05)。见表4、图3。
表4 3组细胞中Skp2、Snaill、E-cadherin和Vimentin蛋白表达情况
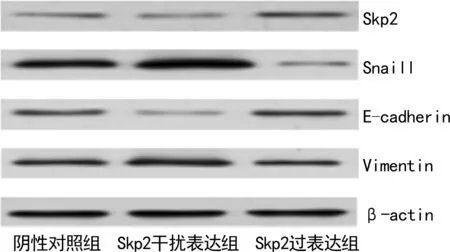
图3 western blot法结果
3 讨 论
上皮性卵巢癌的高致死率很大程度上是由于高侵袭性和易于局部及远端转移造成的,其中EMT起着实质性的作用。在肿瘤进展过程中,卵巢癌细胞EMT表现为上皮细胞特异性标志物如E-cadherin、闭锁蛋白1(ZO-1)等表达下调,而间充质细胞特异性标志物如Vimentin蛋白、N-钙黏蛋白(N-cadherin)等表达增加为特征,这意味着EMT是卵巢癌进展的先决条件[10]。E-cadherin表达下调是EMT的特征性改变,主要受转录因子Snaill的调控,是癌细胞侵袭的前提条件。Snaill可以与E-cadherin启动子中的E-BOX结合,抑制其转录[11]。因此,调控EMT相关因子可能为卵巢癌的治疗提供新的线索。
Skp2是一种E3泛素连接酶[12],调控多种肿瘤抑制因子水平,包括p27和叉头状转录因子O1(FOXO1)[12]。Skp2低表达在包括卵巢癌在内的多种人类癌症中经常被观察到,然而,Skp2与EMT的相关性尚不清楚[13]。本研究探讨了Skp2通过调控Snaill水平在抑制上皮性卵巢癌细胞EMT方面的作用。结果表明,Skp2在卵巢癌组织中表达明显降低,且在其卵巢癌中水平升高则会抑制Snaill的表达,从而抑制卵巢癌细胞的侵袭性。同时,有研究显示,Skp2对Snaill表达有抑制作用,而Snaill对Skp2表达无明显影响[14],表明Snaill是Skp2的下游因子。Snaill被认为是EMT中的转录因子。为了进一步支持上述结果,笔者进一步探讨了Skp2在SKOV3细胞侵袭、迁移中的作用,结果表明,Skp2抑制了SKOV3细胞的侵袭和迁移。
有研究者提出了一个有关EMT的全面监管网络,以强调Skp2的关键作用[14]。然而尚不清楚EMT过程中Skp2发挥作用的确切机制,在某些情况下,Skp2低表达与E-cadherin低表达相关[15]。有研究报道,Skp2主要充当肝癌细胞中与EMT相关的抑制因子,并与基质金属蛋白酶结合抑制其侵袭性,但这些因子与EMT之间的关系尚未确定[16]。此外,磷脂酰肌醇3-激酶(PI3K)/蛋白激酶B(AKT)信号通路的激活导致E-cadherin表达下调和间充质标志物表达的上调。糖原合成酶激酶3β(GSK-3β)通过磷酸化的机制下调Snaill来抑制EMT,而PI3K/AKT通过GSK-3β失活,稳定Snaill来促进EMT[17-18]。本研究观察到促进Skp2表达可以使Snaill蛋白水平降低,抑制卵巢癌细胞EMT,而抑制Skp2表达则作用相反。上述结果对于在EMT中建立Skp2的功能网络非常有意义,但本研究没有对Skp2相关机制进行研究,说服力较欠缺。
综上所述,Skp2在上皮性卵巢癌组织中低表达,它在卵巢癌细胞迁移、侵袭和EMT的调控中起重要作用。通过基因干预技术促进Skp2的表达可以降低SKOV3细胞中Snaill表达,抑制EMT,而抑制Skp2表达则作用相反。
猜你喜欢
昆明医科大学学报(2022年3期)2022-04-19
昆明医科大学学报(2022年1期)2022-02-28
中成药(2018年2期)2018-05-09
中成药(2018年2期)2018-05-09
中成药(2017年3期)2017-05-17
安徽医科大学学报(2016年12期)2017-01-15
中国癌症杂志(2015年4期)2015-12-09
肿瘤预防与治疗(2015年2期)2015-09-26
中国医疗美容(2015年1期)2015-07-12
医学研究杂志(2015年4期)2015-06-10
