
缺铁胁迫下外源亚精胺对番茄幼苗生长及生理特性的影响
2021-01-18 05:23姚棋赵鑫陈浩婷石玉侯雷平张毅
山东农业科学 2020年12期
姚棋,赵鑫,陈浩婷,石玉,侯雷平,张毅
(山西农业大学园艺学院/山西省设施蔬菜提质增效协同创新中心,山西太谷 030801)
铁(Fe)是植物必需的微量元素之一,对于促进植物的光合作用、物质和能量的代谢起着重要作用[1]。缺铁严重影响番茄幼苗的生长发育,导致植株叶片黄化、叶绿体结构发育不完整、叶片捕获光激发能转化为化学能的能力降低,光合能力下降[2,3]。严重缺铁时叶绿体被解离或液泡化,从而影响叶绿素的形成。
虽然大多数土壤中全铁含量较高,但由于受不同土壤因素和植物基因型的影响,植物吸收到体内的可利用铁含量往往不足。土壤有效铁含量不足成为作物生长发育最大的限制因子之一,严重影响植物正常的生长发育[4]。
多胺是广泛存在于植物体内的一类低分子含氮碱,是一种重要的植物生长调节物质,包括腐胺(Put)、精胺(Spm)、亚精胺(Spd)等[5]。Spd由于具多价阳离子特性,其生理功能更强,与植物抗逆境胁迫的关系密切,不仅可直接作为胁迫保护物质,而且还在胁迫信号转导中作为信号分子促进胁迫抗性机制的构建[6]。研究表明,喷施Spd能提高高温胁迫下生菜植株的生长量、根系活力、叶绿素含量,缓解高温对生菜的危害[7]。外源Spd可显著提高高温下番茄幼苗的地上部鲜重和生物量积累,缓解幼苗徒长和叶片黄化,促进光合色素合成,显著增加叶绿素a、叶绿素b和类胡萝卜素的含量[8,9]。多胺除参与种子萌发、植物生长发育、延缓植物叶片衰老外,还可显著提高植物对盐、干旱、低温、重金属等非生物胁迫的耐受性[10]。
目前关于亚精胺(Spd)在植物逆境胁迫方面的研究大多集中在高温胁迫方面,而在重金属等非生物胁迫耐受性等方面的研究还较少,铁胁迫下的番茄抗逆情况还未有研究。因此本试验采用水培法,实施外源喷施Spd处理,研究其对缺铁胁迫和正常生长下番茄幼苗生物量、光合特性及铁素含量的影响,以期为亚精胺调控番茄缺铁胁迫技术的应用提供理论参考。
1 材料与方法
1.1 试验材料与处理
本试验供试番茄品种为‘Micro-tom’。选其饱满种子,浸种催芽、播种。幼苗2片真叶完全展开时定植于日光温室内盛有1/2个剂量的日本山崎番茄配方营养液的水培槽中,缓苗5天后进行试验处理。
试验前两天先进行Spd叶面喷施预处理,以叶片正反两面均匀喷施但无液体滴下为宜。4个处理为:CK(正常铁浓度:100μmol/L EDTAFe);LF(缺铁浓度:10μmol/L EDTA-Fe);Spd(CK+喷施 0.25 mmol/L Spd);LF+Spd(10 μmol/L EDTA-Fe+喷施0.25 mmol/L Spd)。试验处理均用山崎番茄配方营养液栽培,每5天更换1次营养液,每2天进行1次Spd叶面喷施,处理10 d时取样,于-80℃超低温冰箱中保存。
1.2 测定指标及方法
使用皮尺、游标卡尺测量株高、茎粗。地上部和地下部分开后洗净擦干、称其鲜重,后置于烘箱中烘干称干重。取番茄幼苗叶片采用乙醇提取法进行色素含量测定;采用Li-6400光合仪测定净光合速率(Pn);参照何明明[4]的方法测定相对电导率;用火焰原子吸收光度计法测量植株Fe含量,重复3次。
1.3 数据处理
采用Microsoft Excel 2016对数据进行分析和作图,用SPSS 21.0软件进行方差分析,用Duncan’s新复极差法进行多重比较(P<0.05)。
2 结果与分析
2.1 缺铁胁迫下外源Spd对番茄生物量的影响
由表1可知,与CK相比,缺铁胁迫(LF)下番茄幼苗生物量生长受到明显抑制,单株地上部鲜重、地下部鲜重、总鲜重分别降低 45.39%、70.65%和51.79%,株高降低28.14%;与LF相比,LF+Spd处理的单株地上部鲜重、地下部鲜重、总鲜重分别显著增加42.57%、62.96%和45.71%,但株高、茎粗无明显差异;而单喷Spd对幼苗生物量影响较小,较CK未达到显著差异。
2.2 缺铁胁迫下外源Spd对番茄叶片光合色素含量的影响
由表2可知,与CK相比,缺铁胁迫下叶片色素含量均显著降低,LF处理叶绿素a、叶绿素b、类胡萝卜素、叶绿素(a+b)、叶绿素 a/b分别降低27.89%、15.15%、41.67%、23.94%和 14.80%;LF+Spd处理叶绿素a、叶绿素b、类胡萝卜素、叶绿素(a+b)显著高于 LF,分别增加23.58%、12.50%、28.57%和19.75%;与 CK相比,单喷Spd的叶绿素a、叶绿素(a+b)分别增加11.56%和9.86%。
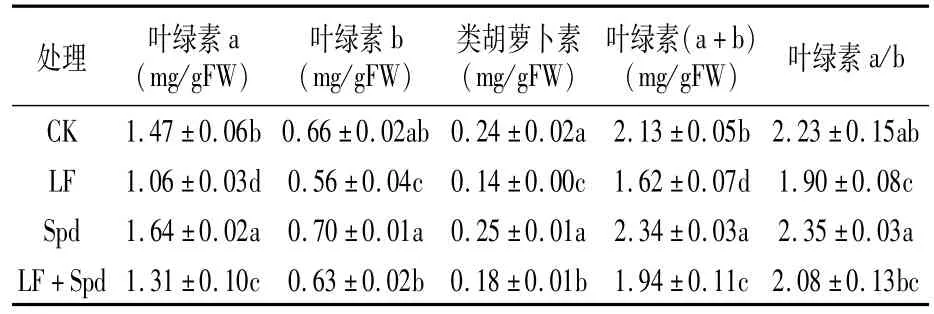
表2 缺铁胁迫下外源Spd对番茄叶片色素含量的影响
2.3 缺铁胁迫下外源Spd对番茄叶片净光合速率的影响
由图1可知,缺铁胁迫下,番茄叶片净光合速率显著降低,LF处理较CK降幅达49.12%;LF+Spd处理较LF净光合速率显著升高,达22.91%;单喷Spd处理较CK净光合速率增加17.89%。
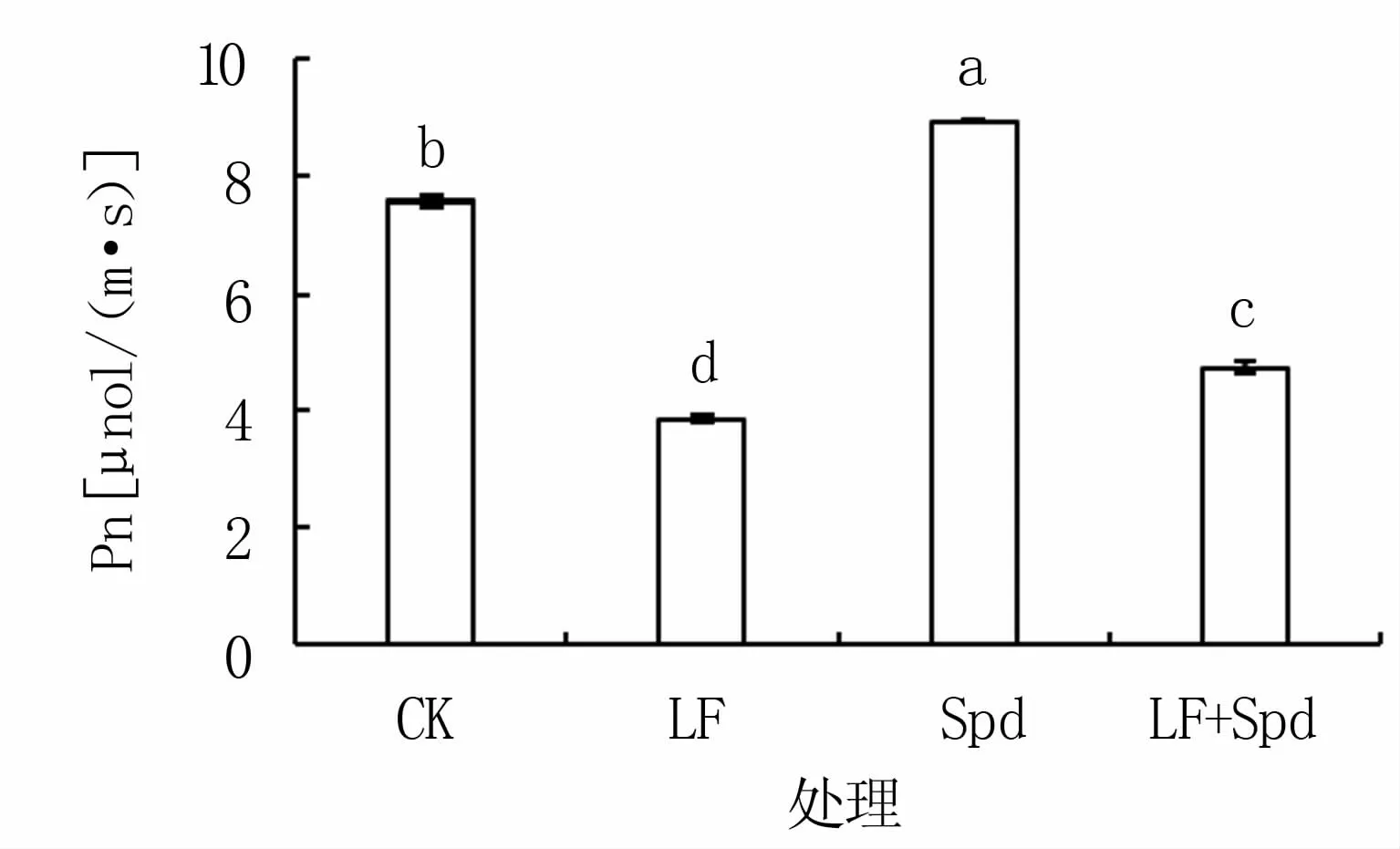
图1 铁胁迫下外源Spd对番茄叶片净光合速率(Pn)的影响
2.4 缺铁胁迫下外源Spd对番茄相对电导率的影响
由图2可以看出,缺铁胁迫下番茄叶片和根系的电导率均显著增加,比 CK 分别增加141.91%和171.09%;LF+Spd处理较LF,其叶片及根系的电导率分别降低15.33%和37.31%,可见根系对缺铁胁迫更加敏感;单喷Spd处理的电导率与CK无显著差异。
2.5 缺铁胁迫下外源Spd对番茄不同部位Fe含量的影响
由表3可知,与CK相比,LF处理植株各部位Fe含量均显著降低;与LF相比,LF+Spd处理叶、茎Fe含量分别降低11.36%和17.96%,根Fe含量增加87.04%;与CK相比单喷Spd处理叶、茎、根Fe元素含量均降低,降幅分别为2.07%、19.25%和45.51%。
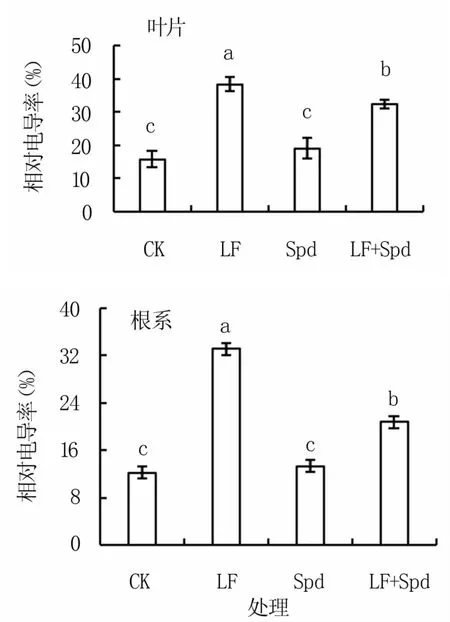
图2 缺铁胁迫下外源Spd对番茄相对电导率的影响

表3 缺铁胁迫下外源Spd对番茄不同部位 Fe含量的影响 (mg/kg)
3 讨论
植物缺铁时,植株矮小、叶片黄化,严重影响其正常生长发育。植株通过改变体内物质的分配,以积累更多的能量应对胁迫,所以生物量是植株生长发育状况最直接的表现[11]。本试验表明,番茄幼苗生长受缺铁胁迫影响显著,苗高明显降低、鲜重减少。这与龙文靖等[12]在玉米干物质积累中的研究结果一致。多胺作为一种生物活性物质,被认为在生物体内信号传递中起“第二信使”的作用,与植物的生长、形态建成和应对胁迫的反应密切相关[13]。喷施Spd后的番茄幼苗生长状况与对照差异不大,这可能是由于试材为矮化品种所致,鲜重虽有所增加,但均未形成显著差异。
叶绿素作为光合作用的物质基础,可将光能转化为化学能,对植物的生长发育至关重要。叶绿素主要包括叶绿素a和叶绿素b,其含量与植物的营养状况密切相关,直接影响光合效率。铁虽然不是叶绿素的组分,但却参与叶绿素的合成[14]。叶绿素含量是判断植物缺铁的标准,逆境胁迫下叶绿素a/b值能反映植物捕获和转化光能的能力。本试验缺铁胁迫下,番茄叶绿素a、叶绿素b、类胡萝卜素含量均显著降低,叶绿素a/b值减小,说明叶绿素合成受到抑制,叶片捕获和转化光能的能力下降;喷施外源Spd后,植株叶片光合色素增加、叶绿素a/b值增大、捕获和转化光能的能力得以恢复。李秀等[15]研究表明,添加外源0.5 mmol/L的Spd可有效保护生姜叶片的(亚)细胞结构,降低高温胁迫对生姜叶片的损伤,提高生姜的耐热性。外源多胺可快速进入完整的叶绿体参与逆境条件下光合元件的保护,说明多胺可能调控叶绿体的发育及衰老进程。
光合作用是植物合成有机物质和获得能量的根本源泉,其强弱对于植物生长、产量及抗逆性等都具有重要影响[16]。光合作用的强弱可以通过光合速率大小来反映,光合速率越大,说明植物光合作用的水平越高。植物在逆境胁迫环境中,净光合速率可能会下降。研究认为,与叶绿素b相比,叶绿素a是更为重要的光合色素,参与光合作用的光化学反应,其含量的减少通常伴随着光合速率的降低[17]。本研究也发现,缺铁胁迫处理显著降低番茄叶片的净光合速率。考虑到铁素在光合作用中的影响机制,将净光合效率作为植物抗缺铁性基因型选择的首选光合参数指标更为合理[18]。张帆等[19]研究表明,外源 Spd可以有效增强低温弱光下辣椒幼苗的光合作用。苏晓琼等[20]研究亦证明,添加外源Spd可以稳定高温胁迫下番茄的光合系统,优化能量在PSⅡ反应中心的分配,进而缓解高温胁迫对番茄光系统的损伤。本研究喷施外源Spd处理的叶片净光合速率显著升高。这是由于外源施加的Spd可快速进入完整的叶绿体,并保护光合机构免受环境胁迫的不利影响;低浓度Spd在体内和体外均增强或恢复光化学活性[21]。
缺铁胁迫与其它逆境伤害一样,最先受到攻击的都是作物的膜系统,产生膜脂过氧化分解和脱脂化作用,膜的完整性被破坏,电解质及某些小分子有机物大量渗漏,破坏细胞物质交换平衡,进而导致一系列生理生化代谢紊乱,使植物受到伤害[22]。研究表明,缺铁胁迫下,番茄幼苗生长受到明显抑制,植株体内的Fe含量和叶绿素含量降低,电解质渗漏率升高[4]。而一定浓度的Spd可显著提高高温胁迫下黄瓜幼苗的膜系统稳定性,缓解叶片MDA的进一步积累,降低叶片相对电导率,提高抗性[5]。本研究中,外源 Spd处理同样降低胁迫下番茄叶片和根系的相对电导率,提高植物抵抗逆境的能力。
总铁含量是植物体内含铁量的重要指标之一,可以体现出植物的铁吸收和分配情况[23]。本试验各处理根系Fe含量均显著高于叶和茎,根与叶、茎中积累的差异,可能与其实际受到的胁迫时间有关,因为植物液泡中含有大量的铁元素[24]。与其它元素相比,铁在植物中的移动性较差,由根系转运到叶枝比较困难,因此在植物体内铁的利用和再利用率很低,也容易在根系中滞留积累[25]。本试验中,缺铁条件下番茄各部位Fe含量均显著降低,喷施Spd显著提高根系Fe元素含量,能有效缓解缺铁胁迫产生的逆境作用,大大提高其耐受性。
4 结论
缺铁胁迫下植株生长受到显著抑制,番茄叶片色素含量大幅度降低,光合作用受到抑制,净光合速率减小,电导率升高,且植株各部位Fe元素含量降低;外源Spd处理有效缓解缺铁胁迫给植株带来的伤害,提高叶绿素a、叶绿素b含量,增大叶绿素a/b比值,增加叶片捕获和转化光能的能力,提高净光合速率,增加根系Fe含量,并通过降低叶片和根系的相对电导率进一步缓解细胞膜脂过氧化程度,提高植物耐受性。
猜你喜欢
舰船科学技术(2022年11期)2022-07-15
农业工程学报(2022年4期)2022-04-24
中国土壤与肥料(2021年5期)2021-12-02
农业与技术(2020年12期)2020-07-04
新课程·下旬(2017年7期)2017-08-14
农业知识·百姓新生活(2014年5期)2015-01-20
食品与健康(2014年4期)2014-06-06
食品工业科技(2014年21期)2014-03-11
意林(2009年16期)2009-05-14
中学生物学(2008年6期)2008-08-29
