
小分子RNA干涉技术在农作物基因沉默中的应用现状
2021-01-18 08:20焦健王义春
农业与技术 2021年1期
焦健王义春
(1.中国科学院科技战略咨询研究院,北京 100190;2.山东滨州阳信县综合检验检测中心,山东 滨州 256600)
引言
近年来,随着生命科学领域研究的日益深入,长度约为20~30个核苷酸(20~30nucleotide [nt])的非编码小RNA(small noncoding RNA sRNA)分子被科研工作者发现。通过对sRNA特征和功能的研究,发现这些sRNA参与了生物体基因和基因组各个信号通路的调控过程,其重要性不言而喻。根据sRNAs的起源、结构特征、相关的效应蛋白和生物学功能,可将sRNAs分为2个主要的类别,即siRNAs(short interfering RNAs)和miRNAs(microRNAs)。这2类sRNAs的发现,向人们揭示出,动植物的基因组非编码区亦蕴含着极其重要的作用和功能。siRNA和miRNA参与调控了动植物的染色质结构的变化、染色体的迁移、转录重编程、蛋白质的翻译以及RNA加工和稳定性保持等重要的生物学过程。
1 sRNA的起源和合成机制
1.1 siRNA和miRNA的起源
miRNA最早是由Ambros及其同事,于1993年在研究控制线虫幼虫时序性发育过程中发现的,当时被命名为一个内源调控基因lin-4[1]。5a后,Mello和Conte报道了有关外源双链RNA分子(double-stranded RNA,dsRNA)通过RNA干涉(RNA interference,RNAi)的机制,可以导致基因的特异沉默[2]。1999年,Tomariand和Zamore证明了植物中的基因沉默机制是通过长度约为20~25nt的siRNAs与靶标基因的碱基互补配对实现的[3]。随后不久,研究证明dsRNA分子可以直接转换为长度约21~23nt的siRNAs;在多种动植物物种中,miRNA介导的生物学作用被广泛报道。至此人们认识到:miRNA作为内源基因的调控子,而siRNA则通过阻止外源核苷酸的干扰(如病毒的感染、转座子的活动和转基因的影响)来保护生物基因组的完整性。
1.2 siRNA和miRNA的合成机制
sRNA的生物合成是在多种基因元件的精细调控下完成的。通过对其生物合成调控网络的详细研究,发现了参与其中的许多主要组分,如,RNA聚合酶Ⅱ(RNA polymerase Ⅱ)、Dicer蛋白酶家族、依赖于RNA的RNA聚合酶(RNA-dependent RNA polymerase,RdRP)、Argonaute蛋白、HEN1以及HST蛋白[4]。
siRNA和miRNA在生物合成机制中,存在以下3个不同点。miRNA是来源于生物体本身内源基因的转录物,并且是受时空调节、有目的性表达的;siRNA主要来自生物体外部,如外源病毒基因组和外来转基因,只有少部分来自于内源的转座子基因。miRNA是从不完全互补配对的双链茎环结构的miRNA前体切割产生;siRNA则来自较长的完全互补配对的双链RNA(double-stranded RNAs,dsRNAs)分子。成熟的siRNA分子主要以双链形式存在,并且以碱基完全互补配对的方式与其靶标基因结合;成熟的miRNA分子均为单链,与靶标基因的结合也并不是完全互补的,存在1~3个碱基的错配。
尽管存在上述差异,但siRNA和miRNA由于在产物大小方面和以序列特异的沉默方式方面极其相似,预示着两者的生物合成和作用机制存在必然联系。已有研究证明,siRNA和miRNA在生物合成过程和作用机制中,均依赖于两类蛋白家族成员——Dicer酶和Argonaute蛋白。其中,Dicer酶负责siRNA和miRNA前体的剪切,最终形成成熟的sRNA分子;AGO蛋白(Argonaute protein)在这两类小RNA的沉默发生机制中,与siRNA和miRNA形成RNA诱导的沉默复合体(RNA-induced silencing complex,RISC),即siRNA-RISC和miRNA-RISC,并发挥核心作用。因此,Dicer酶、AGO蛋白与长度约为20~23nt的双链sRNAs形成了RNA干涉机制中的核心结构特征(图1)[5]。
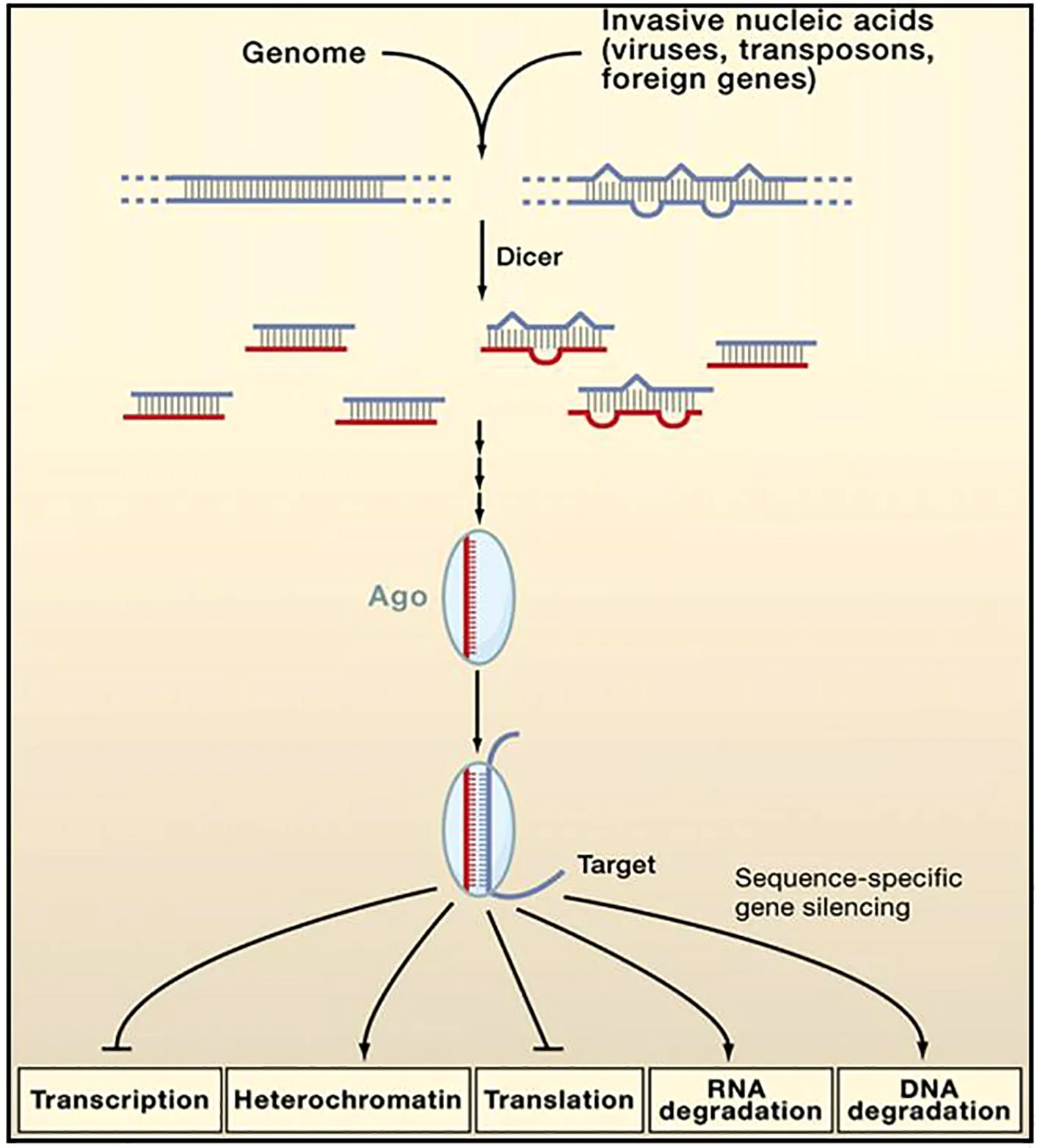
图1 siRNA和miRNA相似的合成和干涉机制[5]
2 sRNA在作物中的生物学功能
2.1 siRNA在作物中的生物学功能研究简介
siRNA参与了农作物生长发育、生物胁迫、非生物胁迫及代谢途径等多种生物学过程,具有重要的生物学功能。在转基因烟草中表达dsRNA以产生siRNA,可有效抑制PMMoV (Pepper mild mottle virus)的侵染,表明siRNA在提高植物抗病毒方面起重要作用[6]。siRNA还可以通过降解卫星RNA(Satellite RNAs)或缺陷干涉RNA(Defective interfering RNAs),显著提高本生烟的抗性[7]。还有报道证实,烟草黄瓜花叶病毒(Cucumber mosaic virus,CMV)可利用病毒卫星RNA产生siRNA,能有效降解寄主烟草中与RNAi相关的基因元件,以防止被寄主RNAi机制干扰病毒自身的生长发育[8]。此外,Wesley等利用RNAi载体转化禾本科作物大麦,通过产生siRNA使新转基因大麦品种对黄矮病毒产生持久抗性[9]。因此,通过siRNA介导的沉默技术结合其它育种技术,可为改良作物品种提供一种有效的技术方法。
2.2 miRNA在作物中的生物学功能研究简介
miRNA介导的基因沉默是农作物体内天然存在的一种防御体系,利用这一体系,可有效抵抗病毒、细菌或真菌病害的侵染。方荣祥研究报道,利用amiRNA靶向黄瓜花叶病毒(CMV)的沉默抑制因子2b,可以有效提高转基因烟草对CMV抗性[10]。另外,对大豆Glyma18g02680.1基因通过amiRNA沉默,导致了对大豆线虫病害抗性的明显增强[11]。孙其信实验室通过对普通小麦进行白粉病接种处理或热激处理,结合Solexa高通量测序,鉴定出了24个与小麦白粉病反应相关及12个与热激反应相关的miRNA。而这些miRNA是否真正参与了小麦白粉病抗病过程及非生物热激胁迫过程,仍需进一步深入研究[12]。此外,通过利用高通量测序法,预测到了58个品质较好的面包小麦miRNA和70个候选的普通小麦保守miRNA,其中23个通过比对得到互补配对的miRNA*序列,可以确定属于小麦的天然miRNA。对于大麦miRNA的研究方法与小麦方面几乎一样,都是通过预测、高通量测序或深度测序再比对的方法,目前已经得到175个大麦miRNA和234个小麦miRNA,有些是植物不同物种间相对保守的,而有些是麦类作物或者单子叶植物特有的miRNA[13]。miRNA也同样参与了农作物生长发育调节的过程。如,水稻中利用miRNA抑制不同基因的表达,如Phytoene desaturase(PDS Os03g08570)、Spotted leaf 11(Spl11 Os12g38210)和Elongated uppermost internode1/CYP714D (Eui1 Os05g40384),导致了水稻叶片光漂白表型[14]。
3 VIGS技术及其在作物功能基因组学中的应用
3.1 VIGS技术简介
由于农作物突变体的收集获得较难,基因沉默已经成为一种有效的研究功能基因组学的反向遗传学工具。病毒诱导的基因沉默(Virus-induced gene silencing,VIGS)作为一种PTGS现象,是作物基因组功能分析的重要技术手段。即在已改造、优化的病毒载体中,插入适当大小的目的基因cDNA片段,随着病毒侵染寄主并在作物组织中大量繁殖后,病毒载体中的目的cDNA片段在作物体内利用siRNA的生物合成途径产生针对靶标基因的多种siRNA,继而通过siRNA干涉机制,降解靶标基因的mRNA,最终实现对目的基因转录水平的沉默或翻译水平的抑制[15]。如图2所示[16],VIGS的基本过程包含病毒载体的构建、寄主作物的病毒侵染、产生sRNA并启动对目标基因的干涉机制。

图2 VIGS的流程示意图[16]
3.2 VIGS病毒载体在作物中的应用
瞬时VIGS在实际应用时需要高侵染效率、高表达、易操作且稳定的病毒载体。经过近年来相关领域的不断发展,研究者成功开发出了多种病毒载体并相继对其进行了合理改造和优化,形成了一系列可分别在单、双子叶作物中应用的VIGS病毒载体。如,大麦条纹花叶病毒(Barley stripe mosaic virus,BSMV)可在单子叶植物中有效实现靶标基因的沉默[14,15]。对于单子叶作物而言,基因功能抑制的研究手段不多且较困难,VIGS作为一种反向遗传学的手段极大地促进了单子叶作物功能基因组的研究,为麦类、水稻和玉米等重要经济作物基因功能分析提供了有效的方法。再如,烟草花叶病毒(Tobacco mosaic virus,TMV)、烟草脆裂病毒(Tobacco rattle virus,TRV)、甘蓝叶卷曲病毒(Cabbage leaf curl virus,CaLCuV)、马铃薯X病毒(Potato virus X,PVX)和卫星病毒诱导的沉默系统(Satellite virus-induced silencing system,SVISS)等可在双子叶植物(烟草、拟南芥和马铃薯等)中对目的基因实现有效的抑制[17]。
因此,VIGS实验时应选择操作简单、侵染性好、寄主发病症状轻以及病毒症状能在植株系统和各个组织中传播的病毒载体。清华大学刘玉乐教授实验室于2010年,报道了一种改造后的甘蓝叶卷曲病毒(CaLCuV)载体,通过农杆菌介导的方式可以有效侵染本生烟植株。该CaLCuV载体不但可以携带目的基因的cDNA片段以引发siRNA介导的基因沉默,而且还可以通过拟南芥AtmiR319前体作为骨架,携带人工微小RNA(artificial microRNAs amiRNA)以引发miRNA介导的基因沉默。并且该系统还可以高表达内源miRNA,内源AtmiRNA156和AtmiRNA165高表达后,其相应的靶标基因在转录水平的表达量明显下调。该载体携带的amiRNA是由WMD3平台设计并选择的,以AtmiR319前体作为骨架,采用Overlap PCR的方法,用适当的amiRNA∶amiRNA*替换内源的miRNA∶miRNA*,并保持原有前体的二级茎环结构。通过对本生烟八氢番茄红素脱氢酶PDS基因的沉默,研究了siRNA和amiRNA所介导VIGS的异同及优缺点,发现amiRNA所介导VIGS沉默效果相当于传统的插入cDNA片段大于300bp所介导沉默的效果,证明CaLCuV∶amiRNA介导的基因沉默更加有效[17]。可见,不论是传统的表达cDNA片段的沉默方法,还是表达sRNA起到沉默效果,最终都在农作物体内表达出小分子RNA以实现对目标基因的沉默。
4 结语
当前,农作物改良和育种进入后基因组时代,解读、编辑与改造农作物基因组显得尤为重要。小分子RNA介导的干涉技术作为一种传统的反向遗传学技术,是农作物基因组改造与基因功能研究必不可少的手段之一,并且相信可以与基因编辑技术、分子设计育种等新兴技术一起,为改良农作物品种、保持作物稳产增产、保障粮食安全等诸多涉及国家战略性的基础领域和产业领域,提供有力科技支撑。
猜你喜欢
当代水产(2022年1期)2022-04-26
军民两用技术与产品(2021年10期)2021-03-16
世界农药(2019年3期)2019-09-10
光学精密工程(2016年4期)2016-11-07
上海农业学报(2016年2期)2016-10-27
西南农业学报(2016年5期)2016-05-17
西南农业学报(2016年6期)2016-04-16
肿瘤影像学(2015年3期)2015-12-09
华东理工大学学报(自然科学版)(2015年4期)2015-12-01
动物营养学报(2015年10期)2015-12-01
