
影响长芒苋种子萌发因素的研究
2021-01-17 00:57吉美静崔海兰陈景超于海燕张金鹏李香菊
杂草学报 2021年3期
关键词:发芽率
吉美静 崔海兰 陈景超 于海燕 张金鹏 李香菊


摘要:长芒苋是我国1985年发现的外来入侵植物,明确长芒苋种子萌發特性,将为其有效防控提供数据支撑。采用室内生物测定方法,对温度、光照、水势、发芽深度等影响长芒苋萌发的关键因子进行研究。结果表明,变温处理可有效提高长芒苋种子的发芽势和萌发率,最适萌发温度为35 ℃/15 ℃变温处理;在35 ℃/15 ℃培养条件下,光照对长芒苋种子的萌发影响不显著;当渗透势为-0.4~-0.2 MPa时,长芒苋种子的萌发率可达82.2%~100%,当渗透势为-0.8 MPa时,仍有36.7%的种子萌发;长芒苋种子的出苗率受播种深度的影响,在播种深度为0.5~1.0 cm时,萌发率最高。由此可见,长芒苋萌发具有广泛的环境适生范围,这也可能是其在我国大面积入侵的生态学基础。
关键词:长芒苋;入侵性杂草;发芽势;发芽率;变温处理
中图分类号:S451 文献标志码:A 文章编号:1003-935X(2021)03-0008-07
Abstract:Amaranthus palmeri,an invasive plant,was first recorded in China in 1985. Clarifying its seed germination characteristics will provide data support for its effective control. Laboratory experiments were conducted to examine the effect of key factors including temperature,light,osmotic stress and burial depth on the seed germination of A. palmeri. The germination potential and germination rate of A. palmeri could be effectively improved in changing temperature,and under the optimum temperature of 35 ℃/15 ℃ day/night,its seed germination was not affected significantly by light. Seed germination rate reached 82.2%~100% when the osmotic potential was -0.4~- 0.2 MPa and 36.7% of the seeds still germinated when the osmotic potential was -0.8 MPa. The optimum burial depth for highest germination rate was 0.5~1.0 cm then decreasing with burial depth. Therefore,A. palmeri was strongly adapted to different environments providing it with the ability to its large-scale invasion in China.
Key words:Amaranthus palmeri;invasive weed;germination potential;germination rate;changing temperature
杂草种子萌发与环境条件的关系是杂草生态学研究的重要组成部分。温度、光照、水分、播种深度是影响种子萌发和出苗的重要因素。由于杂草长期适应自然环境下温度的波动变化,大部分杂草种子在变温条件下更易萌发。如矢车菊[1]和棒头草[2]等在变温条件下,萌发率更高。有报道认为,苋属杂草反枝苋、绿穗苋、长芒苋、西部苋和糙果苋等经过变温处理后,可以有效提高其萌发率[3]。光照也是种子萌发过程中的重要环境因子。光暗交替有利于一些杂草种子的萌发。例如,在昼夜交替的条件下,稗草[4]、藜[5]、野西瓜苗[6]种子的萌发率均显著提高。但也有报道称,在有光的条件下,某些杂草种子发芽受到抑制,从而导致萌发率降低[7]。苋属杂草种子为需光种子,反枝苋在自然光照和全光照下萌发率较高,全黑暗下萌发率显著降低[8]。Taylorson等报道,当温度到达35 ℃时,反枝苋种子在无光的情况下也能萌发[9]。Gallagher等发现,绿穗苋种子萌发在 20 ℃ 下比在30 ℃下对光的需求更高[10]。土壤水分匮乏将延缓种子萌发、降低种子萌发率甚至抑制种子萌发[11]。苋属杂草如反枝苋、长芒苋等作为全球恶性杂草,对环境有较强的适应能力,因此可以耐受超强的干旱胁迫。Ward等报道,苋属杂草种子个体较小,一般在浅土层萌发出苗,主要原因是浅土层的温度、湿度、光照可以满足其种子萌发出苗的要求[12]。除此之外,苋属杂草的萌发与其采样时期及亲本所受外界环境等有密切关系[13-14]。
长芒苋(Amaranthus palmeri),属苋科苋属异株苋亚属,原产于墨西哥西北部和美国的南加州至新墨西哥州到得克萨斯一带,后入侵至世界各地,现欧洲、大洋洲、亚洲等国均有分布[12]。长芒苋具有植株生长迅速、适应性广泛、与作物竞争性强的特性,可导致大豆、棉花、玉米等作物大幅度减产[15-17]。长芒苋繁殖系数高,单株结实量达20万~60万粒[18]。长芒苋种子个体较小,极易通过风力或人类活动、粮谷调运等途径扩散传播[12]。1985年,我国首次在北京市丰台区南苑乡发现长芒苋[19-20]。目前,北京、天津、山东、江苏等地有长芒苋入侵报道,耕地、荒地、河滩、沟渠边、村边、铁路与公路边、仓库周围、加工厂、工地、港口、垃圾场和饲养场周围等地均能成为长芒苋的栖息地[19],并且有多地蔓延的趋势[21-22]。个别长芒苋种群还对除草剂咪唑乙烟酸产生了较高水平的抗性[23-24],给其化学防控带来了一定难度。
我国对于长芒苋生物学、生态学的研究主要集中在分类鉴定[25]、适生性分析[26]及扩散传播[21-22]等方面,对其萌发生物学研究少有报道。基于此,本试验通过设定不同的温度、光照、渗透势、播种深度参数,对长芒苋萌发和出苗与上述参数的关系进行研究,以明确影响长芒苋种子萌发和出苗的环境因素,为其有效治理提供理论依据。
1 材料与方法
1.1 试验材料
2020年8月于中国农业科学院植物保护研究所温室播种长芒苋种子,在控制条件(温度30 ℃/25 ℃,8 h光照/16 h黑暗)下扩繁,10月采收,种子通风晾干后,室内储藏。
1.2 试验方法
1.2.1 温度对种子萌发影响的研究方法 采用培养皿滤纸法:在直径为9 cm的培养皿上垫双层定性滤纸作为发芽床,加入5 mL蒸馏水将滤纸润湿,均匀摆放大小一致、饱满的长芒苋种子30粒。(使用前用5%盐酸浸泡种子1 h,用清水冲洗3遍,并晾干)。
设置6组温度处理,分别为35 ℃/15 ℃、30 ℃/10 ℃、25 ℃/5 ℃、25 ℃、20 ℃、15 ℃。所有处理置于12 h光照/12 h黑暗培养。
1.2.2 光照对长芒苋种子萌发影响的研究方法 采用培养皿滤纸法(同“1.2.1”节)。设光暗交替(12 h光照/12 h黑暗)、全光照(24 h光照/0 h黑暗)和全黑暗(0 h光照/24 h黑暗)3组处理。黑暗处理时间段种子放置在特殊暗箱内。培养皿置于温度35 ℃/15 ℃下培养。
1.2.3 水势对长芒苋种子萌发影响的研究方法 采用PEG-8000溶液模拟干旱胁迫处理。设置4个浓度梯度,分别为-0.2、-0.4、-0.6、-0.8 MPa,以蒸馏水作对照。置于温度35 ℃/15 ℃,12 h光照/12 h黑暗培养。
1.2.4 播种深度对长芒苋种子出苗影响的研究方法 采用盆钵法进行:对田间采集的土壤进行高温灭菌(120 ℃,40 min),作为培养土。设7个播种深度处理,分别为0(土壤表面)、0.5、1.0、20、4.0、6.0、8.0 cm。每钵按照设定的播种深度均匀播种30粒种子后覆土。所有盆钵土壤总质量一致(土壤质量=体积×容重),然后加入蒸馏水使其充分吸水后再进行称重。置于温度 35 ℃/15 ℃、12 h光照/12 h条件下黑暗培养。每天定量补充水分。
上述所有处理置于光照度为5 000 lx的光照培养箱内培养,每日记录长芒苋发芽数。每个处理重复3次。
1.3 数据分析及处理
分别采用下列公式计算长芒苋种子的发芽势和发芽率:
发芽势=萌发前5 d发芽种子数/供试种子总数×100%;
发芽率=发芽种子总数/供试种子总数×100%。
采用SPSS 26.0软件对不同处理的发芽势和发芽率进行统计分析,用邓肯氏新复极差法进行多重比较,利用Sigmaplot 12.5软件进行绘图。
2 结果与分析
2.1 温度对长芒苋种子萌发的影响
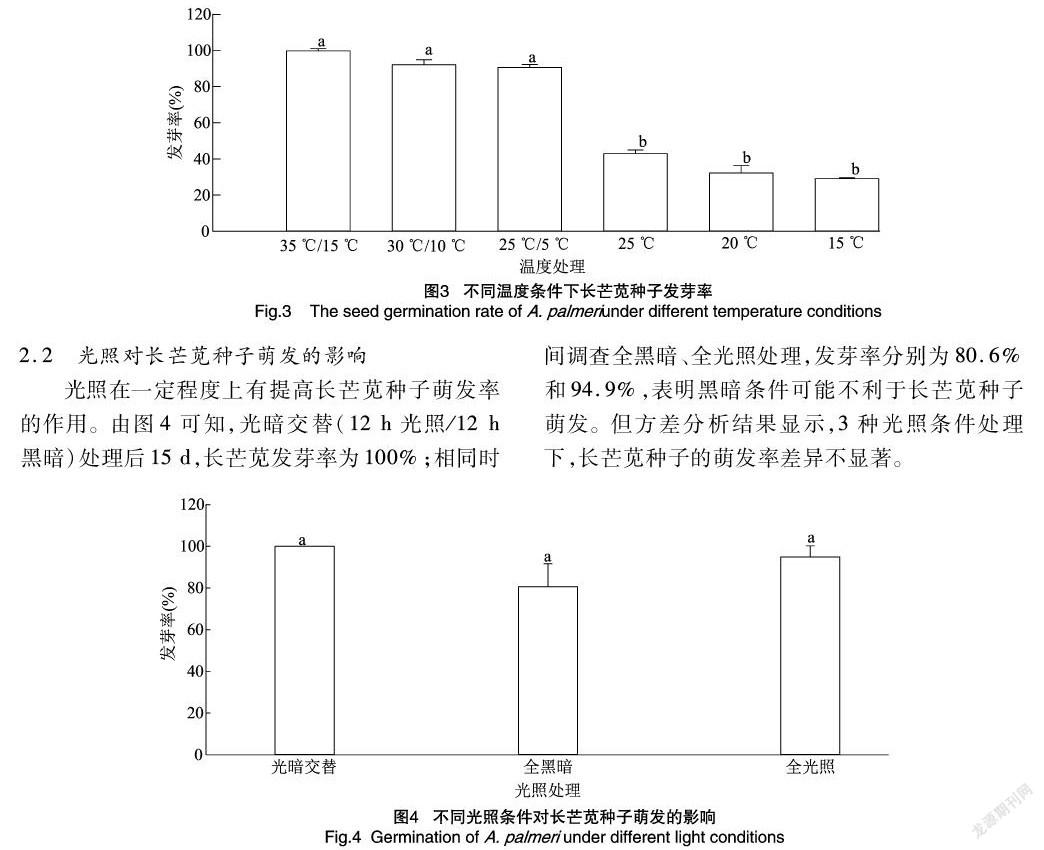
由图1可知,变温对长芒苋种子的发芽有显著促进作用。由图2可知,在培养5 d后,35 ℃/15 ℃变温处理下长芒苋种子发芽势为76.7%,而恒温25 ℃处理下其发芽势仅为38.3%;30 ℃/10 ℃变温处理后,发芽势为67.5%,相应的恒温20 ℃处理下种子发芽势为26.1%;25 ℃/5 ℃变温处理后发芽势为52.1%,其15 ℃恒温对照发芽势仅为153%。由图3可知,培养15 d后,3组变温处理种子发芽率分别为99.7%、92.0%和90.5%,35 ℃/15 ℃ 与30 ℃/10 ℃和25 ℃/5 ℃处理相比,种子发芽率差异不明显;而变温处理与相应的3个恒温处理(25、20、15 ℃)相比,长芒苋种子发芽率在 0.05 水平上差异显著,恒温处理的发芽率分别为429%、32.2%、29.1%。
2.2 光照对长芒苋种子萌发的影响
光照在一定程度上有提高长芒苋种子萌发率的作用。由图4可知,光暗交替(12 h光照/12 h黑暗)处理后15 d,长芒苋发芽率為100%;相同时间调查全黑暗、全光照处理,发芽率分别为80.6%和94.9%,表明黑暗条件可能不利于长芒苋种子萌发。但方差分析结果显示,3种光照条件处理下,长芒苋种子的萌发率差异不显著。
2.3 水分胁迫对长芒苋种子萌发的影响
长芒苋种子萌发的渗透势范围较大。在 PEG-8000 溶液-0.2~-0.8 MPa供试范围内,长芒苋种子均可萌发,但随着PEG-8000溶液渗透势的增大,长芒苋种子的萌发率呈逐渐下降的趋势。从发芽势来看(图5),当渗透势为-0.2、-0.4 MPa 时,发芽势分别为97.8%、67.8%,与其他供试渗透势处理(-0.6、-0.8 MPa)相比,均差异显著。观察15 d后,从发芽率来看(图6),当渗透势为-0.2、-0.4 MPa时,发芽率分别为100%、82.2%,与清水对照相比,差异不显著。当渗透势大于-0.4 MPa时,发芽率显著降低,但当渗透势为-0.8 MPa时,还有大量种子萌发,发芽率达36.7%。
2.4 不同播种深度对长芒苋种子出苗及萌发的影响
不同播种深度影响长芒苋种子的萌发和出苗(图7)。播种深度为0.5~1.0 cm时,长芒苋种子出苗率最高,分别为42.2%和36.7%,随着播种深度的增加,长芒苋种子出苗率降低。播种后 15 d 调查,在0(土壤表面)、2、4、6、8 cm播种深度时,长芒苋种子出苗率分别为12.2%、14.4%、11%、2.2%、0。因此,长芒苋种子最适的播深为05~1.0 cm。
3 结论与讨论
研究结果表明,与恒温处理相比,变温处理长芒苋种子的发芽势及发芽率更高,35 ℃/15 ℃变温处理下长芒苋发芽率可达99.7%。与黑暗处理相比,有光条件下长芒苋种子的萌发率稍高,可能是因为长芒苋种子发芽需要一定的光刺激。随着PEG-8000溶液渗透压的增大,长芒苋种子的萌发率均呈逐渐下降的趋势,在渗透压为-0.2~-0.4 MPa 时,其种子便可达到较高的发芽率,渗透压为-0.8 MPa时,长芒苋种子仍能萌发,萌发率为36.7%。长芒苋种子出苗受播种深度的影响,在播种深度为0.5~1.0 cm时,出苗率最高。
种子萌发受环境因子及自身生理条件的影响,不同种类的杂草萌发需要特定的环境条件[27]。大多数研究表明,交替温度处理能提高种子的萌发率。魏雨其等报道,在变温25 ℃/15 ℃、30 ℃/15 ℃、30 ℃/20 ℃处理下,圆囊苔草、膨囊苔草和异穗苔草的发芽率显著提高[28]。王江涛等研究发现,垂穗披碱草的最适萌发温度为25 ℃/10 ℃;三毛草和草地早熟禾的最适萌发温度为25 ℃/15 ℃[29]。从本研究结果来看,长芒苋种子最适萌发温度为 35 ℃/15 ℃,比恒温25 ℃处理发芽率显著提高,长芒苋变温处理发芽率高可能是其适应自然环境温度变化的结果。
光照对杂草种子萌发影响因素的研究有很多。按照发芽对光照的要求,杂草种子分光敏型和非光敏型2类。如鳢肠[7]、胜红蓟[30]等种子发芽需要光刺激,而播娘蒿等种子萌发受可见光的抑制[31],光敏色素可能是其萌发受抑制的内在原因。除此之外,Zhao等报道光照对看麦娘种子的萌发似乎没有任何影响[32]。Singh等发现,圆叶牵牛花萌发不需要光照[33]。光敏型杂草种子可能由于弱光或光的波长变化,导致发芽率降低,因此,生产实践中对于光敏型杂草种子采取遮阴或植物秸秆覆盖等措施,可为杂草的非化学防治提供新思路。温度和光照是苋属种子萌发过程中的重要因子[20,34],两者的相互作用也影响种子萌发率[35],例如在低温条件下,黑暗处理有利于牛筋草种子的萌发,而随着温度的升高,光照和黑暗处理条件下种子萌发差异不显著[36]。表明光照和温度可协同作用影响种子萌发。关于长芒苋是否存在光照与温度的互作而造成萌发率差异,尚需进一步试验研究。
水分是杂草种子萌发的重要因子。本研究随着PEG-8000溶液渗透势的增大,长芒苋种子的萌发率呈逐渐下降的趋势,这在其他杂草中也存在类似情况,如看麦娘[35]、节节麦[37]、菵草[38]、鳢肠[17]等也会随着渗透压的增大,萌发率逐渐降低。然而有的杂草如旱雀麦[39]对极端干旱胁迫生境也能适应。杂草出苗率与其播种深度有关。一般来讲,禾本科杂草在较深土层也能出苗,如节节麦、牛筋草、看麦娘等[40-42]。而长芒苋种子较小,顶土能力较差,在浅土层易萌发出苗[1],在较深土层的长芒苋种子出苗率低。Keeley等报道,长芒苋种子播种深度小于1.3 cm时,萌发和出苗高于播种深度5 cm处理[5]。本研究长芒苋种子在播种深度为0.5~1.0 cm时,出苗率高于较深土层,与前人报道结果一致。生产中,可以利用较深土层的长芒苋种子不能出苗这一特性,采用深翻措施将表土层长芒苋种子翻耕至较深土层,减少其出苗率。
长芒苋作为外来入侵性杂草,具有较强的适应性。其种子的萌发和出苗对温度、光照、水分及播种深度要求不严格,增加了其传播扩散的能力,也对未来我国长芒苋防除与治理增加了难度。
参考文献:
[1]Clements C D,Harmon D,Young J A.Diffuse knapweed (Centaurea diffusa) seed germination[J]. Weed Science,2010,58(4):369-373.
[2]Wu X,Li J,Xu H L,et al. Factors affecting seed germination and seedling emergence of Asia minor bluegrass (Polypogon fugax) [J]. Weed Science,2015,63(2):440-447.
[3]Steckel L E,Sprague C L,Stoller E W,et al. Temperature effects on germination of nine Amaranthus species[J]. Weed Science,2004,52(2):217-221.
[4]Boyd N,van Acker R. Seed germination of common weed species as affected by oxygen concentration,light,and osmotic potential[J]. Weed Science,2004,52(4):589-596.
[5]Jursik M,Soukup J,Holec J. Seed dormancy and germination of shaggy soldier (Galinsoga ciliata Blake.) and common lambsquarter(Chenopodium album L.) [J]. Plant, Soil and Environment,2011,49(11):511-518.
[6]Chachalis D,Korres N,Khah E M. Factors affecting seed germination and emergence of venice mallow (Hibiscus trionum) [J]. Weed Science,2008,56(4):509-515.
[7]Chauhan B S,Johnson D E. Influence of environmental factors on seed germination and seedling emergence of eclipta (Eclipta prostrata) in a tropical environment[J]. Weed Science,2008,56(3):383-388.
[8]程偉霞,陈双臣,李文亮. 不同处理对反枝苋种子萌发特性的影响[J]. 河南农业科学,2009,38(11):94-96,104.
[9]Taylorson R B,Hendricks S B. Action of phytochrome during prechilling of Amaranthus retroflexus L. seeds[J]. Plant Physiology,1969,44(6):821-825.
[10]Gallagher R S,Cardina J. Phytochrome-mediated Amaranthus germination Ⅰ:Effect of seed burial and germination temperature[J]. Weed Science,1998,46(1):48-52.
[11]Javaid M M,Tanveer A. Germination ecology of Emex spinosa and Emex australis,invasive weeds of winter crops[J]. Weed Research,2014,54(6):565-575.
[12]Ward S M,Webster T M,Steckel L E,et al. Palmer amaranth (Amaranthus palmeri):A review[J]. Weed Technology,2013,27(1):12-27.
[13]Cristaudo A,Gresta F,Luciani F,et al. Effects of after-harvest period and environmental factors on seed dormancy of Amaranthus species[J]. Weed Research,2007,47(4):327-334.
[14]Jha P,Norsworthy J K,Riley M B,et al. Shade and plant location effects on germination and hormone content of palmer amaranth (Amaranthus palmeri) seed[J]. Weed Science,2010,58(1):16-21.
[15]Webster T M,Everest J,Brecke B,et al. Broadleaf crops subsection (cotton,peanut,soybean,tobacco,and forestry) [J]. Proceedings-Southern Weed Science Society,2001,58:291-306.
[16]Bensch C N,Horak M J,Peterson D. Interference of redroot pigweed (Amaranthus retroflexus),Palmer amaranth (A. palmeri),and common waterhemp (A. rudis) in soybean[J]. Weed Science,2003,51(1):37-43.
[17]Massinga R A,Currie R S,Horak M J,et al. Interference of Palmer amaranth in corn[J]. Weed Science,2001,49(2):202-208.
[18]Keeley P E,Carter C H,Thullen R J. Influence of planting date on growth of palmer amaranth (Amaranthus palmeri) [J]. Weed Science,1987,35(2):199-204.
[19]李振宇. 长芒苋——中国苋属一新归化种[J]. 植物学通报,2003,38(6):734-735.
[20]车晋滇. 外来入侵杂草长芒苋[J]. 杂草科学,2008,26(1):58-60.
[21]马金双. 中国入侵植物名录[M]. 北京:高等教育出版社,2013.
[22]莫训强,孟伟庆,李洪远. 天津3种外来植物新记录——长芒苋、瘤梗甘薯和钻叶紫菀[J]. 天津师范大学学报(自然科学版),2017,37(2):36-38,56.
[23]吉美静,黄兆峰,李香菊,等. 靶标基因突变导致入侵性杂草长芒苋对咪唑乙烟酸产生抗性[J/OL]. 植物保护:1-7. (2020-12-11)[2021-01-02]. https://doi.org/10.16688/j.zwbh.2020639 .DOI.org/10.16688/j.zwbh.2020639.
[24]吉美靜,黄兆峰,崔海兰,等. 一个抗咪唑乙烟酸长芒苋种群的发现[J]. 杂草学报,2020,38(3):23-27.
[25]徐 晗,赵彩云,刘勇波,等. ITS序列及其SNP位点在外来入侵杂草长芒苋、西部苋和糙果苋物种鉴定中的应用[J]. 植物保护,2017,43(5):128-133.
[26]曹晶晶,王 瑞,李永革,等. 外来入侵植物长芒苋在中国不同地区的表型变异与环境适应性[J]. 植物检疫,2020,34(3):25-31.
[27]Baskin J M,Baskin C C. Physiology of dormancy and germination in relation to seed bank ecology[M]. San Diego:Academic Press,1989:53-65.
[28]魏雨其,王显国,孔令琪,等. 变温以及酸、碱处理对打破3种苔草种子休眠的作用[J]. 草业与畜牧,2016,37(2):3-9.
[29]王江涛,杨 军,王向涛. 变温处理对藏北高原3种禾本科牧草种子萌发的影响[J]. 高原农业,2019,3(6):670-674.
[30]杜丽思,李 铷,董玉梅,等. 胜红蓟种子萌发/出苗对环境因子的响应[J]. 生态学报,2019,39(15):5662-5669.
[31]李怡慧,徐 琼,尹梦婕,等. 环境因素对播娘蒿种子萌发的影响[J]. 山东农业科学,2020,52(9):45-48.
[32]Zhao N,Li Q,Guo W L,et al. Effect of environmental factors on germination and emergence of shortawnfoxtail (Alopecurus aequalis)[J]. Weed Science,2018,66(1):47-56.
[33]Singh M,Ramirez A H M,Sharma S D,et al. Factors affecting the germination of tall morningglory (Ipomoea purpurea)[J]. Weed Science,2012,60(1):64-68.
[34]Gallagher R S,Cardina J. Phytochrome-mediated Amaranthus germination Ⅱ:Development of very low fluence sensitivity[J]. Weed Science,1998,46(1):53-58.
[35]Butler T J,Celen A E,Webb S L,et al. Temperature affects the germination of forage legume seeds[J]. Crop Science,2014,54(6):2846-2853.
[36]马亚杰,马小艳,陈全家,等. 环境因素對不同地区牛筋草种子萌发的影响[J]. 中国农学通报,2019,35(17):60-74.
[37]Fang F,Zhang C X,Wei S H,et al. Factors affecting Tauschs goatgrass (Aegilops tauschii Coss.) seed germination and seedling emergence[J]. Journal of Agricultural Science,2011,4(1):114-121.
[38]Rao N,Dong L Y,Li J,et al. Influence of environmental factors on seed germination and seedling emergence of American sloughgrass (Beckmannia syzigachne) [J]. Weed Science,2008,56(4):529-533.
[39]Guo W L,Liu W T,Li L X,et al. Molecular basis for resistance to fenoxaprop in shortawn foxtail (Alopecurus aequalis) from China[J]. Weed Science,2015,63(2):416-424.
[40]王晓阳,于海燕,杨 娟,等. 不同环境因素对节节麦萌发的影响[J]. 植物保护,2019,45(3):196-200.
[41]Mahmood A H,Florentine S K,Chauhan B S,et al. Influence of various environmental factors on seed germination and seedling emergence of a noxious environmental weed:Green galenia (Galenia pubescens) [J]. Weed Science,2016,64(3):486-494.
[42]Nosratti I,Amiri S,Bagheri A,et al. Environmental factors affecting seed germination and seedling emergence of foxtail sophora (Sophora alopecuroides) [J]. Weed Science,2018,66(1):71-77.
猜你喜欢
天津农业科学(2022年6期)2022-07-19
果农之友(2020年4期)2020-12-23
农业与技术(2019年19期)2019-11-11
热带农业科学(2019年2期)2019-04-28
农民致富之友(2018年9期)2018-06-27
江苏农业科学(2017年1期)2017-02-27
现代农业科技(2016年20期)2016-12-20
天津农业科学(2016年4期)2016-04-20
江苏农业科学(2015年11期)2016-01-27
天津农业科学(2015年11期)2015-12-03
