
KRTAP36-1基因在子午岭黑山羊中的鉴定及其与羊绒性状的关联分析
2021-01-14 06:52宋宜泽赵孟丽郝志云罗玉柱李少斌沈继源王继卿
中国农业大学学报 2021年2期
宋宜泽 赵孟丽 郝志云 罗玉柱 刘 秀 李少斌 沈继源 柯 娜 王继卿
(甘肃农业大学 动物科学技术学院/甘肃省草食动物生物技术重点实验室/甘肃省牛羊基因改良工程实验室,兰州 730070)
山羊绒纤维由次级毛囊产生,被称为“软黄金”[1-2]。羊绒纤维主要由角蛋白(Keratins,Ks)和角蛋白相关蛋白(Keiatin associated proteins,KAPs)构成[3],这2种蛋白的重量占羊绒纤维重量的90%以上。Ks组装成角蛋白中间丝(Keratin intermediate filaments, KIFs)蛋白,构成了羊绒纤维骨架,KAPs通过二硫键填充于KIFs周围[4]。KAPs最显著的特征是含有丰富的半胱氨酸(Cysteine,Cys)、或者甘氨酸(Glycine,Gly)和酪氨酸(Tyrosine,Tyr)[5]。根据氨基酸的组成和含量,KAPs可以分为3大类,分别是:高甘氨酸/酪氨酸KAPs(HGT-KAPs,含有35~60 mol%的Gly/Tyr)、高硫KAPs(HS-KAPs, 含有≤30 mol%的Cys)和超高硫KAPs(UHS-KAPs, 含有>30 mol%的Cys)[6]。根据序列相似性,KAPs可被进一步划分为不同的家族。例如,KAP6~8、KAP18~22和KAP36同属于HGT-KAPs[7-8]。
KAP蛋白的编码基因标识为KRTAPs,其长度较小,一般只包含1个单独的外显子[9]。研究表明,KRTAPs基因的核苷酸序列变异与山羊的羊绒性状密切相关。例如,Wang等[7]在陇东绒山羊上发现,KRTAP20-1等位基因A的存在与较大的绒层高度和产绒量相关(P<0.05),等位基因B的存在与较小的产绒量和平均纤维直径的相关(P<0.05);Zhao等[19]发现KRTAP15-1等位基因A能显著降低陇东绒山羊的平均纤维直径(P=0.006);Wang等[20]发现KRTAP20-2等位基因A的存在与陇东绒山羊较大的产绒量相关(P<0.001),而等位基因B的存在与较小的产绒量相关(P<0.01)。
KRTAP36家族只包含KRTAP36-1基因一个成员。在绵羊上已发现,KRTAP36-1基因位于绵羊的1号染色体上,在新西兰罗姆尼羊等3个群体中共检测到3个等位基因和4个单核苷酸多态性(Single nucleotide polymorphisms,SNPs)。相关性分析表明,等位基因A的存在与较大的产绒量相关(P=0.035),等位基因C的存在与较大的平均纤维直径相关(P=0.025)[8]。目前有关KRTAPs基因的研究主要集中在人类和绵羊上。例如,在人类基因组中,共鉴定出来自25个家族的80多个KRTAPs基因[10-11];在绵羊基因组中,共发现来自13个家族的29个KRTAPs基因。但在山羊基因组中目前只发现来自12个基因家族的15个KRTAPs,包括KRTAP1-1[12]、KRTAP1-4[13]、KRTAP6-2[14]、KRTAP7-1[15]、KRTAP8-1[16]、KRTAP8-2[15]、KRTAP9-2[17]、KRTAP11-1[18]、KRTAP13-1、KRTAP13-3[12]、KRTAP15-1[19]、KRTAP20-1[7]、KRTAP20-2[20]、KRTAP24-1[21]和KRTAP28-1[22]。因为KRTAPs基因序列具有种间保守性,说明山羊基因组上还有大量的KRTAPs基因有待进一步鉴定。
子午岭黑山羊是我国一个历史悠久的地方品种,以盛产黑猾皮和紫绒著称,毛黑而光亮,有优美的波浪形卷曲,花穗美观,羊绒纤维细,是高档纺织原料。因此,本试验以子午岭黑山羊为研究对象,采用生物信息学和比较基因组学方法,在山羊基因组中鉴别出KRTAP36-1基因。采用已广泛应用于KRTAPs基因研究的单链构象多态性(Single strand conformation polymorphism,SSCP)分析和测序法,检测该基因的SNPs。最后分析该基因的核苷酸序列变异对子午岭黑山羊羊绒性状的影响。
1 材料与方法
1.1 试验材料
在甘肃省环县钰生绒山羊繁育有限公司,选择11只无亲缘关系的子午岭黑山羊种公羊,采用人工授精技术对子午岭黑山羊母羊进行配种。在出生的后代羔羊中,随机选择342只健康无病和生长发育正常的子午岭黑山羊作为研究对象。当这些羔羊生长至12月龄时,现场测定产绒量,背中线处测定绒层高度。同时在背中线处采集山羊绒样本,用于实验室测定羊绒细度。对于测定了羊绒性状的所有子午岭黑山羊,颈静脉采血8 mL,加入酸性柠檬酸葡萄糖(Acid-citrate-dextrose,ACD)抗凝,用苯酚-氯仿法提取血液基因组DNA。
1.2 引物设计与PCR反应
以绵羊KRTAP36-1基因的编码区序列(GenBank登录号:MK770620)作为模板,用GenBank的BLAST功能,在山羊基因组GCF_001704415.1(www.ncbi.nlm.nih.gov/assembly/GCF_001704415.1)中进行同源性搜索。在搜索结果中,与绵羊KRTAP36-1序列相似性最高的山羊序列被假定为子午岭黑山羊的KRTAP36-1基因。据此序列,设计特异性引物(上游引物:5′-GGTTTACCACACCCACAATG-3′,下游引物:5′-GTAGCATAGCAAGAGTGAAG-3′),扩增367 bp的山羊假定的KRTAP36-1基因。引物由上海生工生物工程有限公司合成。
采用20 μL反应体系进行PCR扩增,包括:10 μL Taq预混酶,0.25 μmol/L的上、下游引物各0.8 μL,0.8 μL DNA模板和7.6 μL去离子水。反应条件为:94 ℃预变性5 min;94 ℃变性30 s,57 ℃退火30 s,72 ℃延伸30 s,共35个循环;72 ℃终延伸9 min;4 ℃低温保存。用1.5%的琼脂糖凝胶电泳检测PCR扩增效果。
1.3 PCR-SSCP检测
在1 μL PCR扩增产物中加入9 μL变性缓冲剂(0.025%溴酚蓝、98%去离子甲酰胺、0.025%二甲苯氰和10 mmol/L EDTA)。混合物在105 ℃变性 9 min 后立即放入冰水混合物中,上样于16×18 cm、12%(Acr∶Bis=37.5∶1)的聚丙烯酰胺凝胶中,在0.5×TBE、6 ℃(恒温)、350V电压下电泳18 h。电泳结束后,根据Byun等[23]描述的方法对聚丙烯酰胺凝胶进行银染显色。
1.4 核苷酸序列测定及相关性分析
核苷酸序列测定方法因纯合子和杂合子而异。若个体的基因型是纯合子,直接对PCR扩增产物测序;若个体全部为杂合子(无纯合子个体),则根据Gong等[24]建立的方法切胶测序。序列测定在北京奥科鼎盛生物科技有限公司进行。使用MEGA(V.5.0)进行序列比对和分析,并用最大简约法构建系统发育树,进化树的置信度以1000次自展分析进行重复检验。
采用SPSS(V.20.0)软件的一般线性混合效应模型(General liner mixed-effect models,GLMMs),分析KRTAP36-1核苷酸序列状态(存在或缺失)和基因型对子午岭黑山羊3个羊绒性状(产绒量、羊绒平均纤维直径和绒层高度)的影响。为确定建模因素,分析了性别、父本(配种公羊)和出生等级(单羔和双羔)对子午岭黑山羊羊绒性状的影响。结果表明,性别和父本对羊绒性状有极显著影响(P<0.01),出生等级(单羔和双羔)对羊绒性状没有显著影响(P>0.05)。因此在GLMMs模型中,将核苷酸序列(或基因型)和性别作为固定效应,父本作为随机效应。由于所有的子午岭黑山羊均饲养于同一条件,且都在周岁时测定羊绒性状,因此在模型中没有考虑环境和年龄因素。具体模型如下:
核苷酸序列对羊绒性状影响的研究模型:
Y=μ+Variant+Gender+Sire+e
(1)
基因型对羊绒性状影响的研究模型:
Y=μ+Genotype+Gender+Sire+e
(2)
其中,Y为羊绒性状表型值,μ为群体均值,Variant为核苷酸序列状态(用0和1分别表示缺失和存在),Genotype为基因型,Gender为性别,Sire为父本(配种公羊),e为随机误差。
2 结果与分析
2.1 子午岭黑山羊KRTAP36-1基因鉴定
以绵羊KRTAP36-1基因的编码区序列(GenBank登录号:MK770620)作为模板,应用BLAST程序在山羊基因组GCF_001704415.1中进行同源性搜索。结果显示,在山羊1号染色体上发现了一个174 bp(nt3675662-nt3675835)的开放阅读框,这个片段与绵羊KRTAP36-1基因的编码区序列有98.28%的同源性。该片段周围分布有已经鉴定出的11个山羊KRTAPs基因,它们分别是KRTAP11-1、KRTAP7-1、KRTAP8-1、KRTAP8-2、KRTAP6-2、KRTAP20-2、KRTAP20-1、KRTAP15-1、KRTAP13-1、KRTAP13-3和KRTAP24-1。
经PCR-SSCP分析,在342只子午岭黑山羊中,共检测到A、B和C3条核苷酸变异序列(图2),组成AA、BB、AB和AC4种基因型。

图2 子午岭黑山羊KRTAP36-1基因的PCR-SSCP检测
基于人类、绵羊和山羊中已鉴别的所有HGT-KAPs的氨基酸序列,以及本研究获得的3条山羊序列,构建了系统进化树。结果显示,这3条序列与绵羊的KAP36-1首先聚为一支,显示出了最高的序列相似性。这表明本研究发现的3条序列来自于山羊KRTAP36-1基因。

竖线表示KRTAPs基因的位置,箭头表示基因的转录方向,竖条下面的数字代表KRTAPs基因的名称(如36-1代表KRTAP36-1)。
2.2 子午岭黑山羊KRTAP36-1基因的核苷酸序列变异
测序结果表明,子午岭黑山羊KRTAP36-1基因的3条核苷酸序列中存在2个SNPs,其中一个SNP(c.-18T/C)位于KRTAP36-1基因的5′UTR区域。另一个SNP(c.119A/G)为错义突变,导致第40位的酪氨酸变为半胱氨酸(p.Tyr40Cys)。
2.3 山羊KAP36-1蛋白的氨基酸组成
山羊KRTAP36-1的3条核苷酸序列都编码一条由57个氨基酸残基组成的多肽链。这些多肽链含有高含量的Gly(36.84 mol%)和Tyr(26.32~28.07 mol%),也含有中等含量的Ser(12.28 mol%)以及较低含量的Leu(8.77 mol%)、Phe(7.02 mol%)、Arg(3.51 mol%)、Pro(1.75 mol%)、Met(1.75 mol%)和Cys(0~1.75 mol%)。值得注意的是,在核苷酸序列A和B翻译生成的多肽链中均不含Cys,只在核苷酸序列C翻译生成的多肽链中发现了一个Cys残基。
2.4 子午岭黑山羊KRTAP36-1的基因型频率和核苷酸序列频率
在342只子午岭黑山羊中,核苷酸序列A、B和C的频率分别为68.13%、31.73%和0.15%。基因型AA、BB、AB和AC的频率分别为40.06%、3.80%、55.85%和0.29%。
2.5 KRTAP36-1基因的核苷酸序列变异对子午岭黑山羊羊绒性状的影响
在子午岭黑山羊上发现的3条核苷酸序列中,由于序列C的频率低于5%,故将此序列排除在核苷酸存在/缺失模式之外。结果表明,核苷酸序列A的存在与较小的产绒量相关(P<0.001),B的存在与较大的产绒量相关(P=0.004)。A和B的存在(或缺失)对羊绒平均纤维直径和绒层高度无影响(P>0.05)(表1)。
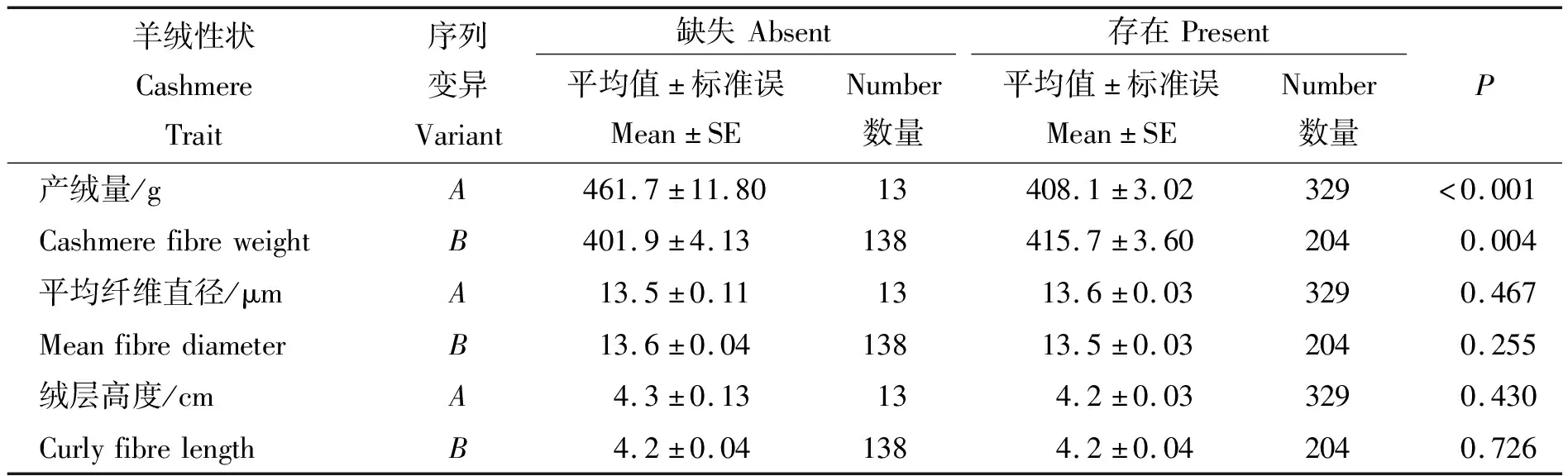
表1 KRTAP36-1核苷酸序列变异与子午岭黑山羊羊绒性状间的相关性
对于AA、AB和BB3种优势基因型,分析了基因型对子午岭黑山羊羊绒性状的影响。结果发现,基因型影响了山羊的产绒量。BB型山羊的产绒量极显著高于AB和AA型山羊(P<0.001)(表2)。
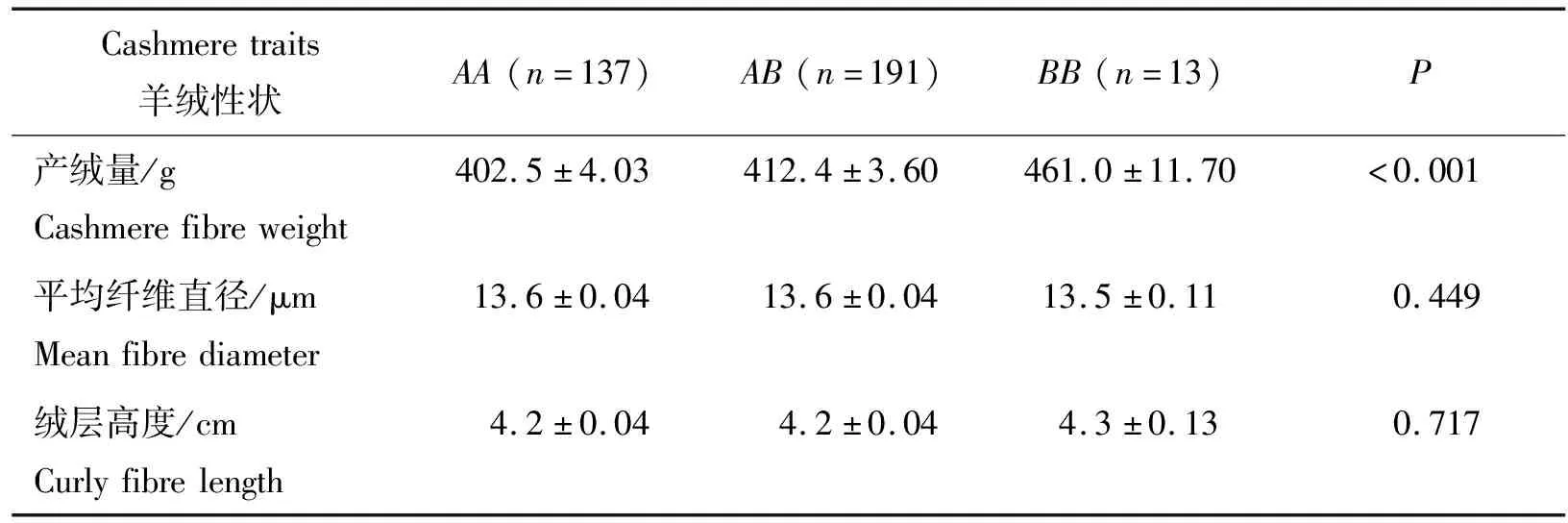
表2 KRTAP36-1基因型与子午岭黑山羊羊绒性状间的相关性
3 讨 论
本研究从子午岭黑山羊基因组中鉴定出了一个编码HGT-KAP蛋白的KRTAP36-1基因,并分析了该基因的遗传变异及其与子午岭黑山羊羊绒性状的相关性。山羊KRTAP36-1基因分布于1号染色体上,位于KRTAP20-1与KRTAP15-1之间,这与KRTAP36-1基因在绵羊染色体上的位置一致[8]。在山羊、绵羊和人类已鉴定的所有HGT-KAP序列中,山羊的KRTAP36-1序列与绵羊的KRTAP36-1序列有最高的同源性,可达97%。

人类、绵羊和山羊的KAPs序列分别用“h”、“s”和“g” 表示。本研究鉴定的山羊KAP36-1序列用方框显示。山羊、绵羊和人类HGT-KAPs的GenBank登陆号为:NM 001193399(sKAP6-1),KT725832(sKAP6-2),KT725837(sKAP6-3),KT725840(sKAP6-4),KT725845(sKAP6-5),X05639(sKAP8-1),KF220646(sKAP8-2),MH243552(sKAP 20-1),MH071391(sKAP 20-2)KX377616(sKAP 22-1),MK770620(sKAP36-1),NM 181602(hKAP 6-1),NM 181604(hKAP 6-2),NM 181605(hKAP 6-3),AJ457063(hKAP 7-1),AJ457064(hKAP8-1),AJ457067(hKAP19-1),NM 181608(hKAP19-2),NM 181609(hKAP19-3),NM 181610(hKAP19-4),NM 181611(hKAP19-5),NM 181612(hKAP19-6),NM 181614(hKAP19-7),AB096964(hKAP19-8),NM 181615(hKAP 20-1),NM 181616(hKAP 20-2),NM 181619(hKAP 21-1),NM 181617(hKAP 21-2),NM 181620(hKAP 22-1),AY510121(gKAP7-1),AY510122(gKAP8-1),AY510123(gKAP8-2),MG742218(gKAP20-1),MF973462(gKAP20-2)。

引物序列用横线标示,“-”表示与gKRTAP36-1*A的序列相同,大写字母表示KRTAP36-1的编码区序列,小写字母表示非编码区序列,2个SNPs显示在序列上方(包括1个错义突变),起始密码子ATG和终止密码子TGA用方框显示,SNPs的顺序和书写格式遵循了HGVS的命名规则(http:∥varnomen.hgvs.org/)。
在山羊KAP36-1中发现了高比例的Gly和Tyr(Gly含量为36.84 mol%,Tyr含量为26.32~28.07 mol%),这符合HGT-KAPs蛋白质含有35~60 mol%的Gly/Tyr这一特征。在山羊中,目前已研究了KAP6家族、KAP7-1、KAP8-1、KAP8-2、KAP20-1和KAP20-2这些HGT-KAPs蛋白的氨基酸组成。与以上HGT-KAPs蛋白相比,山羊KAP36-1中Gly+Tyr的含量与KAP6家族[25]、KAP20-1[7]和KAP20-2[20]接近,但显著高于KAP7-1的34.52%、KAP8-1的40.30 mol%和KAP8-2的46.78 mol%[25]。研究发现,Gly和Tyr在HGT-KAP的结构和性质中发挥了重要作用。Gly是20种氨基酸中侧链最小的氨基酸,它可以影响KAP螺旋结构的形成,最终影响KAP和KIFs的交互作用[22]。Tyr是包含苯环结构的芳香族氨基酸,它可以通过π的交互作用调节KIFs的排列,最终影响羊绒纤维的结构[7]。通过以上Gly+Tyr含量比较结果表明,不同的HGT-KAPs蛋白,含有不同含量的Gly+Tyr,这也意味着这些HGT-KAPs蛋白在羊绒纤维形成过程和理化性质中发挥了各自不同的作用。同时,本研究还发现山羊KAP36-1蛋白含有极少量的Cys(0~1.75 mol%),这远远低于山羊KAP20-1的7.94~9.52 mol%[7]、KAP20-2的14.52 mol%[20],也低于KAP7-1的5.95 mol%和KAP8-1的6.50 mol%[25]。Cys是羊绒和羊毛纤维合成的第一限制性氨基酸,它形成了KAPs和KIFs之间交联的二硫键。山羊KAP36-1中只有极少量的Cys,这表明KAPs和KIFs之间还存在其它形式的交联作用。本研究在山羊KRTAP36-1核苷酸序列A和B编码的多肽链中未发现Cys,在C编码的多肽链中发现了一个Cys,这与绵羊KRTAP36-1的研究结果一致。Gong等在绵羊KRTAP36-1中共发现了3个等位基因,这3个等位基因编码的多肽链中均没有Cys。
本研究在山羊KRTAP36-1基因中发现了2个SNPs,少于绵羊上发现的4个SNPs[8],这可能与所用的试验群体有关。本研究的试验对象是342只纯种的子午岭黑山羊,而绵羊KRTAP36-1研究中使用了46只新西兰罗姆尼羊、48只美利奴羊和321只美利奴羊X无角短毛羊的杂种后代。这表明,如果以后的研究中使用更多的山羊品种,有可能在山羊KRTAP36-1基因上发现更多的核苷酸序列和SNPs。本研究在子午岭黑山羊上发现的2个SNPs中,其中一个位于5′UTR区域,另一个为非同义突变,这与绵羊KRTAP36-1中发现的SNPs类型一致。Gong等[8]在绵羊KRTAP36-1中发现了1个5′UTR区域的SNP和3个非同义突变SNPs。表明这2个物种中KRTAP36-1基因的多态性有可能产生于同一种机制。但是在本研究中并未发现绵羊KRTAP36-1中的SNPs。
在本研究子午岭黑山羊中发现的2个SNPs中,c.119A/G导致了多肽链上的第40位Tyr变为Cys,鉴于Tyr和Cys在羊绒纤维中的重要作用,山羊KRTAP36-1基因的这个SNP有可能对羊绒纤维性状有重要的影响。然而,由于本研究中选用的342只子午岭黑山羊中,这个SNP产生的核苷酸序列C的频率仅为0.15%,故无法通过相关性分析研究这种影响。建议将来在其他山羊品种和更多的山羊样本中,进一步研究c.119A/G对山羊羊绒性状的影响。另外一个SNP(c.-18T/C)处于非编码区,虽然不能导致编码氨基酸的改变,但它的功能同样不能被忽视。因为这个SNP有可能与KRTAP36-1基因重要区域的其它SNPs连锁,也有可能影响基因的转录结合位点、转录效率和mRNA的稳定性,最终导致基因的功能发生变化[22]。序列比对发现,c.-18T/C导致了核苷酸序列B与A和C之间的序列差异,结合核苷酸序列B影响了产绒量这一相关性分析结果,说明c.-18T/C这个SNP影响了羊绒纤维的结构和性质。
在山羊1号染色体上分布的12个KRTAPs基因中,前人已在相同试验群体中研究了KRTAP15-1、KRTAP20-1、KRTAP20-2和KRTAP24-1对产绒量、羊绒纤维直径和绒层高度这3个性状的影响。结果发现,KRTAP15-1和KRTAP24-1影响了羊绒纤维直径[19,21],KRTAP20-1和KRTAP20-2影响了产绒量和绒层高度[7,20]。本研究中发现同样位于1号染色体的KRTAP36-1基因影响了产绒量。在山羊1号染色体上,与KRTAP36-1基因距离最近的是KRTAP20-1基因,它们之间的距离约为50 kb,而KRTAP20-1基因的核苷酸序列变异也影响了山羊的产绒量(P<0.001)。由此推断,KRTAP36-1基因对山羊产绒量的影响有可能是与KRTAP20-1基因相连锁的结果。
相关性分析结果表明,核苷酸序列A的存在与较小的产绒量相关,B的存在与较大的产绒量相关。在342只子午岭黑山羊中,核苷酸序列A和B的频率分别为68.13%和31.73%。这表明在本试验群体中,对产绒量这一性状的人工选择并不强烈。因此,在以后的育种过程中,应该有意识地选留含有序列B的个体,淘汰含有序列A的个体,以提高子午岭黑山羊的产绒量。除了以上描述的KRTAPs基因以外,Qiao等[26]用全基因组关联分析发现AKT1、ALX4、HK1和NT-3等基因参与了毛囊发育,与绒山羊细度有关。Li等[27]用全基因组重测序发现FGF5、SGK3、IGFBP7、OXTR和ROCK1基因与绒长、产绒量等绒山羊的羊绒性状有关。
4 结 论
本研究首次在山羊1号染色体上鉴定出KRTAP36-1基因,在342只子午岭黑山羊的该基因中检测到3条变异核苷酸序列和2个SNPs。相关性分析表明,KRTAP36-1的核苷酸序列变异和基因型显著影响了子午岭黑山羊的产绒量。该研究丰富了山羊基因组数据,筛选出了提高产绒量的分子标记,为子午岭黑山羊羊绒性状的分子选种提供了科学依据。
猜你喜欢
云南畜牧兽医(2022年4期)2022-08-19
云南畜牧兽医(2021年6期)2021-12-24
学苑创造·A版(2021年10期)2021-10-30
云南畜牧兽医(2021年5期)2021-10-27
南方农业·下旬(2021年3期)2021-07-28
农家科技中旬版(2020年4期)2020-05-25
现代农业科技(2018年14期)2018-10-31
流行色(2018年5期)2018-08-27
兽医导刊(2016年6期)2016-05-17
质量探索(2014年12期)2014-02-04
