
宿根黄化对甘蔗主要农艺性状及内源激素的影响
2021-01-13 05:05:15罗霆邓宇驰丘立杭陈荣发范业赓周慧文闫海锋黄杏周忠凤吴建明
热带作物学报 2021年12期
关键词:农艺性状
罗霆 邓宇驰 丘立杭 陈荣发 范业赓 周慧文 闫海锋 黄杏 周忠凤 吴建明



摘 要:为了探明宿根黄化对甘蔗主要农艺性状及内源激素变化的影响,对发生宿根黄化后甘蔗体内激素水平高低、叶绿素指数变化,以及分蘖、株高等主要农艺性状进行分析。结果表明,黄化株经过了近40 d的生长,叶绿素指数达到40,与正常株刚出苗时的叶绿素指数相当,经过62 d基本完成复绿。正常株的株高、叶片数、节数和分蘖数的增量分别是黄化株的1285%、371%、860%和1650%。在5个观察期,正常株和黄化株的株高、叶片数、节数和分蘖数都有极显著差异;黄化株赤霉素含量比正常株低21.2%,生长素和细胞分裂素含量分别比正常株高17.3%和12.9%,与正常株差异显著;黄化株的玉米素、脱落酸和乙烯含量分别比正常株提高54.5%、195.5%和47.4%,与正常株差异极显著。本研究初步探索了宿根黄化对甘蔗主要农艺性状及内源激素的影响,为甘蔗宿根黄化病的有效防控提供依据。
关键词:甘蔗宿根黄化;农艺性状;内源激素;叶绿素指数
中图分类号:S566.1 文献标识码:A
Abstract: The hormone level, chlorophyll index, tiller number and plant height were analyzed to investigate the effects of chlorosis on the main agronomic traits and endogenous hormone changes of ratoon sugarcane. After 40 days of growth, the chlorophyll index of the chlorosis plant reached 40, equal to the chlorophyll index of the normal plant at the time of emergence, and it took 62 days for the chlorosis plant to turn green and get normal. The increment of plant height, leaf, internode and tiller number of normal plant was 1285%, 371%, 860% and 1650% of that of chlorosis plants, respectively. And there were significant differences between normal and chlorosis plants at 5 determination periods. The contents of gibberellin, auxin and cytokinin were 17.3% and 12.9% higher than those of normal strains, respectively, which were significantly different from normal strains. The contents of zeatin, abscisic acid and ethylene in chlorosis plants increased by 54.5%, 195.5% and 47.4%, respectively, which were significantly different contrast to normal plants. This study initially revealed the effects of chlorosis on the main agronomic traits and endogenous hormones of sugarcane, and would provide some knowledge for the prevention and control of chlorosis in ratoon sugarcane.
Keywords: chlorosis in ratoon sugarcane; agronomic traits; endogenous hormone; chlorophyll index
DOI: 10.3969/j.issn.1000-2561.2021.12.024
甘蔗是最重要的糖料作物,我國90%的食糖来源于甘蔗。广西是我国最大的产蔗区,历年甘蔗种植面积占全国的60%~75%。广西的甘蔗种植以“一年新植两年宿根”为主,宿根蔗面积常年占甘蔗总种植面积的65%左右,持续稳定的宿根蔗产量对广西蔗糖产业发展有举足轻重的作用。
甘蔗的宿根黄化是一种生理性病害,目前仅见宿根蔗发生,表现为发株后失绿,长出黄色甚至接近白色的叶片,在无处理的情况下,通常50~70 d能自动复绿,但在黄化期间,植株生长“僵滞”,几乎不能分蘖和拔节,新叶抽出速度异常缓慢,造成减产20%~40%的严重后果[1]。
甘蔗宿根黄化病呈现逐年加重的趋势,从2012年前后报道的仅在桂南蔗区酸性粘重的土壤上零星发生,到2020年3—5月本研究小组不完全调查发现,甘蔗宿根黄化病已覆盖桂南上思、崇左、百色、田东、田阳、桂中来宾、柳州、桂南沿海北海、钦州等县(区),共27个蔗区[2]。近十余年,研究者相继开展宿根黄化病研究。目前认为,土壤中微量元素不平衡是主要病因,土壤中过量酚酸类物质累积,加速锰、铝等有毒物质积累后,造成植株体内铁锰比例失衡而黄化[3-6];在此过程中,高锰抑制了铁的活性,铁向叶绿体运输受阻,使得甘蔗幼苗生理性缺铁黄化[5]。但也有研究认为是宿根蔗蔸留土时间长,老根在高盐分、高酚酸类化感物质、高矿质元素累积的土壤中逐渐缺乏活力,导致甘蔗根系对养分和水分的吸收能力不足造成黄化[6-8]。宿根蔗黄化复绿研究也取得一定进展,研究发现,施用铵态氮肥有利于减轻幼苗叶片黄化,加速黄化叶片复绿[9];叶面喷施铁、镁、硼、钙、钾、锌、氮等营养元素均能提高黄化苗的转绿率,以喷施铁的效果最好,其次是镁、硼[1];土施硅肥、石灰等也能在一定程度上降低当年的宿根黄化率[10-11];叶面喷施硫酸亚铁、硫酸锰、硫酸镁和钼酸铵能改善黄化病情,提高作物产量[12]。此外,还发现甘蔗的宿根黄化存在明显的基因型差异,此差异与甘蔗品种锰的含量、分布及铁素营养有关[3]。综上所述,研究者开展了甘蔗宿根黄化病因及复绿措施研究,取得一定进展,但对致病原因的探索主要集中在外部因素,特别是土壤矿质元素丰缺对黄化发生的影响上,目前尚无探索宿根蔗自身的生理或生长发育特点与黄化发生相关性的相关研究。对复绿措施的研究尚少,找到一些能降低发病率的措施,但还没有能在生产上根本解决甘蔗宿根黄化病的方法。鉴于此,本研究小组开展了甘蔗宿根黄化病对甘蔗主要农艺性状及内源激素变化的影响研究,对甘蔗发生宿根黄化病后的激素水平高低、叶绿素指数变化,以及分蘖、株高等主要农艺性状进行探讨,以期为甘蔗宿根黄化病的有效防控提供依据。
1 材料与方法
1.1 材料
1.1.1 试验点概况 在广西崇左市扶绥县岜盆乡那何村选择一块宿根黄化病蔗地为试验点(图1),试验点东经107.90°,北纬22.52°。甘蔗品种为‘桂糖42号’。
1.1.2 仪器设备与试剂 超高效液相色谱与质谱联用仪为美国AB SCIEX公司生产的4000 QTRAP型。生长素(IAA)、油菜素内酯(BR)、细胞分裂素(CTK)、赤霉素(GA)、玉米素(ZT)、脱落酸(ABA)和乙烯(ETH)标样均购于美国Sigma-Aldrich有限公司;C18萃取小柱购买于沃特世公司;0.22 μm微孔滤头、甲醇、乙醇和乙腈(色谱纯)购于生工生物工程(上海)股份有限公司。
1.2 方法
1.2.1 重要农艺性状的测量与统计 2020年3月1日起,将试验地块按五点法划分成东、南、西、北、中5个小区,每个小区中随机取病株和正常株各18株,利用SPAD手持叶绿素仪,隔天测定植株的叶绿素指数直至复绿;每10 d测定株高、节数、分蘖数,记录叶片数至复绿。株高从地面测量到第1片见肥厚带的叶鞘处,节数以节和节间都完全拔出记1节,分蘖数以田间实际计数为准。
1.2.2 植物激素的测定 2020年3月1日,在试验点随机选取病株和正常株各18株,参考并改良朱莉莉等[13]和Dobrev等[14]的方法测定IAA、BR、CTK、GA、ZT、ABA和ETH的含量。
取+1叶,用清水漂洗数次后用液氮迅速冻干,称取0.1 g冻干样品于EP管中,用组织破碎研磨仪充分破碎,加入5 mL 80%甲醇提取液于装有样品粉末的EP管中,于4 ℃冰箱内浸提过夜。浸提液于4 ℃下,12 000 r/min高速离心15 min,将上清液转入干净离心管,加入200 μL提取液,4 ℃,12 000 r/min高速离心15 min,此过程重复1次。依次用10 mL纯乙醇,20 mL超纯水,10 mL 80%甲醇通过装有提取液的C18萃取小柱(200 mg,3 mL),使过滤柱活化。最后将纯化后的提取液用0.22 μm的有機相滤膜过滤备用。
1.2.3 定量工作曲线的建立 分别准确称取10 mg的IAA、BR、CTK、GA、ZT、ABA和ETH标准品,用100%甲醇定容至1 mL,配制成10 mg/mL的标准贮备液。使用时分别吸取上述标
准贮备液配制成10 μg/mL的混合标准溶液,并进一步稀释成一系列的标准溶液。最终标准曲线相关系数R2>0.99方可使用。
1.2.4 色谱-质谱条件 色谱条件:Waters C18(100 mm×2.1 mm, 3 μm)色谱柱,柱温25 ℃,样品盘温度4 ℃,进样量5 μL,分析时间10 min。质谱条件:多反应检测模式(MRM);气帘压力25 psi,离子化电压–4500 V,离子源温度600 ℃。
1.3 数据处理
采用Microsoft Excel软件处理数据和作图,利用DPS 7.05软件对数据进行方差分析。
2 结果与分析
2.1 叶绿素指数变化
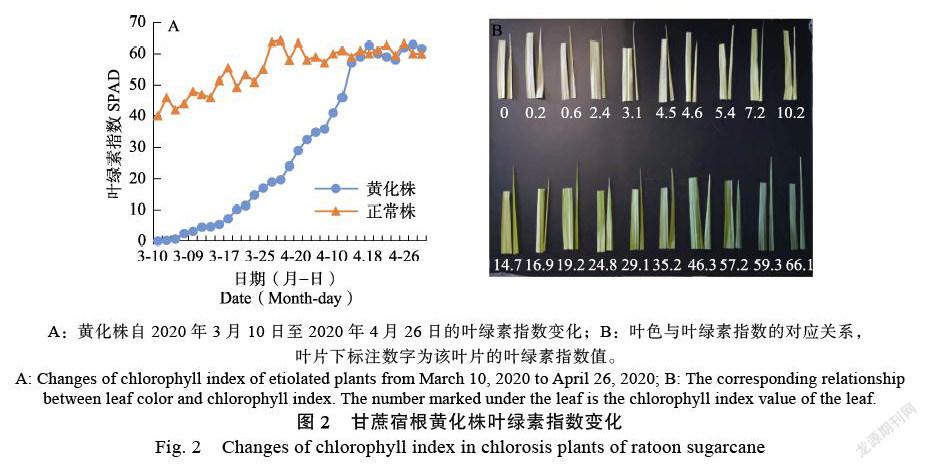
叶绿素指数Spad值是衡量叶绿素相对含量的参数,通过测量叶片在650 nm和940 nm 2种波长范围内的透光系数来确定当前叶片的叶绿素相对含量。所选试验地在2020年1月20日砍收后,2月10日开始陆续出苗,2020年3月1日基本完成出苗。由图2可见,正常株出苗后叶绿素指数保持在Spad值在40以上并随着植株的生长发育快速提高,出苗15 d时Spad值达到50,其后稳定在50~65之间。黄化株在刚出苗时叶绿素指数极低,一些植株的叶绿素指数甚至为0,经过近40 d的生长,Spad值达到正常株刚出苗时40的水平。说明宿根黄化对甘蔗体内叶绿素含量有显著影响,明显降低了植株体内的叶绿素含量。4月14日后,正常株和黄化株的叶绿素指数持平,此时距2月10日始苗日已有62 d,黄化株体内的叶绿素水平恢复正常,植株完成复绿。
2.2 宿根黄化对甘蔗重要农艺性状的影响
甘蔗宿根黄化病影响植株叶绿素形成,使植物不能进行正常的光合作用,造成植株营养供应受阻,细胞结构破坏,甘蔗生长发育受到严重影响。如图3所示,宿根蔗在黄化期间,特别在3月21日以前,即叶绿素指数Spad值在20以下时,生长几乎停滞。黄化株的株高、叶片数、节数和分蘖数均处于极低水平,且在3月1日至3月21日期间,黄化植株只长高0.7 cm,叶片数增加0.35片,节数增加0.25节,分蘖数增加0.1个,而正常植株长高9.0 cm,叶片数增加1.3片,节数增加2.15节,分蘖数增加1.65个。正常株的株高、叶片数、节数和分蘖数的增量分别是黄化株的1285%、371%、860%和1650%。除了在宿根刚发蔸的3月1日,黄化株和正常株都没有分蘖,二者间没有明显差异外,此后的5个观察期,正常株和黄化株的株高、叶片数、节数和分蘖数都存在极显著差异。尽管4月14日后黄化株基本复绿,叶绿素水平恢复正常,但复绿前的生长“缓慢”状态延续了60多天,植株的株高、叶片数、节数和分蘖数都远低于正常水平,说明宿根黄化严重影响了甘蔗的生长发育。
2.3 宿根黄化对甘蔗内源激素的影响
植物内源激素的生理效应复杂多样,从影响细胞的分裂、伸长、分化到影响植物发芽、生根、伸长、分蘖、开花、休眠和脱落等,对植物的生长发育有重要的调控作用。甘蔗发生宿根黄化后,其体内的激素含量发生明显变化,总体表现为赤霉素含量降低,生长素、脱落酸、玉米素、乙烯、细胞分裂素升高。由图4可见,黄化株赤霉素含量为74.5 pmol/L,正常株赤霉素含量为90.3 pmol/L,黄化株赤霉素含量比正常株低21.2%,二者间存在显著差异;黄化株生长素含量为21.0 μmol/L,正常株生长素含量为17.9 μmol/L,黄化株生长素含量比正常株高17.3%,二者差异显著;黄化株和正常株的细胞分裂素含量分别为53.9 pg/mL和47.7 pg/mL,黄化株比正常株高12.9%,二者差异显著。玉米素、乙烯和脱落酸受黄化的影响更大, 黄化株的玉米素、脱落酸和乙烯含量分别为3.4 ng/mL、45.5 ng/mL和1107.2 pmol/L,比正常株的2.2 ng/mL、15.4 ng/mL和751.3 pmol/L分别提高了54.5%、195.5%和47.4%。黄化株的玉米素、乙烯和脱落酸含量和正常株相比有极显著差异。
3 讨论
3.1 叶绿素含量与农艺性状的相关性
叶绿素是植物进行光合作用的主要色素,它在光合作用的光吸收中起核心作用。龙春芬[15]研究认为,烟草叶面积和总叶绿素含量存在极显著相关关系,在烟草生长发育前期,总叶绿素的含量随着叶面积的增大而增加;刘明等[16]研究认为,高大韧水稻剑叶的叶绿素含量高,则水稻根系活力较强,长势旺;姚晓云等[17]研究发现,黄叶稻在抽穗期和灌浆期叶绿素各项指标最低,其穗长、结实率和产量也低于其他对照品种;彭晓邦等[18]研究证实,叶绿素含量较低的白花丹参的花序长度、复叶数、叶面积最低,生长势也最差。李传宗[19]和谢宁昆[20]在鉴定水稻黄叶性状时发现,黄化突变体的穂长、株高、每穗总粒数、结实率及千粒重与野生型相比有着明显的降低;此外,叶绿素指数Spad值还被证实与玉米产量、木瓜水分利用效率和叶冠大小呈显著的正相关,与玉米穗轴成熟度、木瓜的水分利用效率呈负相关[21-22]。在本研究中,前期叶绿素指数Spad值低于20期间,黄化植株的生长基本停滞,Spad值缓慢上升至与正常株相当时,植株逐渐恢复生长,长高、拔节和叶片抽出速度也逐渐恢复正常。可见,叶绿素含量对甘蔗的多个农艺性状有较大影响。
在本研究中,宿根甘蔗发生黄化症状后,虽然经历了62 d的缓慢生长最终复绿,但株高、叶片数、分蘖数、节数都比正常株极显著减少。这与Li等[23]研究遮阴62 d的玉米Spad值下降后,虽解除遮阴部分生长恢复,但产量仍下降的研究结果相符。Fan[1]认为,这种前期黄化的生长缓慢状态,对后期产量有严重影响。Zhao等[24]认为,甘蔗Spad值下降造成减产,是因为光合作用和光合产物积累受阻;阿布都克尤木·阿不都热孜克等[25]进一步研究了Spad值降低造成减产的生理机制,认为Spad值降低导致橡胶草叶片和根系中的总酚、总黄酮、有机酸组分、绿原酸和咖啡酸含量下降,光合性能降低,光合产物累积能力及输出能力受阻。Spad值與作物产量和农艺性状的相关性虽然多次被证实,也证实叶绿素下降引发了多个代谢和生理过程改变,但叶绿素降低引发甘蔗宿根黄化现象的生理机制仍未知,值得深入探索。
3.2 植物内源激素与黄化的相关性
植物激素作为植物内源发育程序的中介,可以整合外部信号从而控制植物生长,调节植物从胚胎发生、器官生成、生物防御、胁迫耐受到生殖发育等各个方面的活动。植物产生的主要激素是生长素(IAA)、赤霉素(GA)、细胞分裂素(CK)、脱落酸(ABA)、乙烯(ET)、水杨酸(SA)、茉莉酸(JA)、油菜素类甾体(BR)和独脚金内脂等。其中,生长素、赤霉素、细胞分裂素能促进植物生长和发育过程,而脱落酸和乙烯的作用则是抑制植物生长,促进成熟和衰老。这几种激素在植物生长发育的不同时期除各有其独特作用外,还能互相促进或抑制,充分发挥调节植物生长发育的作用[3]。前人研究发现,植物内源激素能对叶色造成影响,如油菜素内酯(BRs)能通过抑制黑暗中原叶绿素酸酯的过量积累而促进植物黄化幼苗见光转绿[26];IAA能抑制叶绿素合成和叶绿体发育,诱导叶色黄化[27];茉莉酸也可以在光照条件下诱导烟草叶片的黄化[28]。在本研究中,甘蔗宿根黄化与激素水平变化表现出明显的相关性,黄化株的赤霉素含量显著降低,生长素、脱落酸、玉米素、乙烯、细胞分裂素显著或极显著升高。李根[29]发现,IAA与GA3的互作以及IAA调控在枳砧‘红绵蜜柚’叶片黄化过程中扮演重要的角色,在黄化表型中,GA3含量下降,这与本研究中黄化株的赤霉素含量显著降低相符。贾兵等[30]认为,外源GA3能诱导黄化叶片内FER1、FER2、FER3、FRO2、IRT1和FD1的表达,揭示了GA3在诱导‘黄金梨’黄化叶复绿中起作用;Ranwala等[31]也发现喷施100 mg/L的GA4能完全防止采后百合叶片黄化。可见,赤霉素的调控和表达与黄化高度相关。
在本研究中,黄化株体内IAA含量显著增加。Han等[32]研究也发现,缺铁胁迫下,黄化小金栎的根系IAA含量是正常铁水平下的4~5倍,认为IAA可能是诱发苹果属植物缺铁反应的信号因子。此外,Chen等[33]证实,H2S通过调节缺铁溶液中大豆植物激素相关基因表达丰度来改变植物激素浓度,完全抑制了叶脉间黄化,提高了大豆幼苗生物量。由此推断IAA很可能是引发植物缺铁黄化的重要因素。除了IAA外,黄化株的脱落酸、玉米素、乙烯、细胞分裂素都显著或极显著升高,Takeshi[34]认为,这有可能是激素对生长所需养分的消耗有加速作用,使叶片叶绿体缺少养分而收缩变小,而且激素浓度越高,叶片叶绿体收缩程度越大,植株越容易黄化。
前人研究发现与叶片黄化相关的差异基因富集在激素信号转导途径中,植物激素信号转导的差异基因,如IAA、POD、HCT和CAD,以及调控生长素、茉莉酸和乙烯信号转导途径中SAUR、JAZ和ERF1等基因显著上调或下调表达,都对植物黄化症状有重要的调控作用[28, 35]。本研究证实了甘蔗宿根黄化能引起内源激素的显著变化,但对与此相关的差异基因或信号转导途径未知,这值得进一步研究。
参考文献
[1] Fan Y G, Chen R F, Qiu L H, et al. Quantitative proteomics analysis of sugarcane ratoon crop chlorosis[J/OL]. Sugar Tech, 2021: 1-9. DOI: 10.1007/s12355-021- 00952-0.
[2] 罗 霆, 吴建明, 邓宇驰, 等. 广西宿根甘蔗黄化病现状及病因分析[J]. 甘蔗糖业, 2021, 50(1): 68-73.
[3] 常敏敏, 梁莉燕, 杨 曙, 等. 高锰诱导甘蔗幼苗黄化的基因型差异研究[J]. 西南农业学报, 2019, 32(9): 1980- 1985.
[4] 黎晓峰. 锰诱导甘蔗幼苗缺铁黄化及抗性机制研究[Z]. 广西大学, 2016-03-26.
[5] 龙光霞. 铁、锰对广西宿根甘蔗幼苗黄化的影响[J]. 南方农业学报, 2020, 51(8): 1939-1945.
[6] 李贤宇, 周洁琼, 兰 靖, 等. 土壤中有毒物质对宿根甘蔗苗期黄化的效应研究[J]. 湖北农业科学, 2016, 55(14): 3641-3647.
[7] 罗维钢, 刘要鑫, 仇惠君, 等. 宿根甘蔗黄化苗根系活力及其营养特性研究[J]. 南方农业学报, 2013, 44(6): 930- 933.
[8] 李贤宇. 化感作用与宿根甘蔗苗期黄化的相关性初探[D]. 南宁: 广西大学, 2015.
[9] 于 华, 陈杨明珠, 黎晓峰. 氮素形态对甘蔗锰毒黄化的影响[J]. 西南农业学报, 2018, 31(9): 1821-1824.
[10] 凌桂芝, 蒋雄英, 杨 曙, 等. 硅对甘蔗锰毒的缓解作用研究[J]. 西南农业学报, 2016, 29(7): 1639-1643.
[11] 陈杨明珠, 杨 曙, 梁丽萍, 等. 石灰及其与有机肥配施对甘蔗幼苗黄化的影响[J]. 热带作物学报, 2018, 39(6): 1050-1054.
[12] 刘要鑫, 罗维纲, 仇惠君, 等. 宿根甘蔗黄化苗调控研究[J]. 广东农业科学, 2013, 40(21): 32-35.
[13] 朱莉莉, 陈雅雯, 王棚涛, 等. HPLC-MS/MS同时测定植物6种內源激素含量方法的优化[J]. 河南大学学报(自然科学版), 2020, 50(3): 298-304.
[14] Dobrev P I, Havlicek L, Vagne M, et al. Purification and determination of plant hormones auxin and abscisic acid using solid phase extraction and two-dimensional high performance liquid chromatography[J]. Journal of Chromatography A, 2005, 1075(1): 159-166.
[15] 龙春芬. 湘南浓香型烟叶产区质体色素含量的动态变化规律研究[D]. 长沙: 湖南农业大学, 2013.
[16] 刘 明, 何龙飞, 李创珍, 等. 稍为高大韧水稻结实率较低的生理原因的研究[J]. 广西农业生物科学, 2008(3): 241-245.
[17] 姚晓云, 蓝海军, 邓 伟, 等. 水稻淡白叶突变体的叶绿素含量测定及农艺性状比较分析[J]. 江西农业学报, 2020, 32(12): 12-15.
[18] 彭晓邦, 秦绍龙. 商洛不同花色丹参光合生理特性及农艺性状比较研究[J]. 陕西农业科学, 2020, 66(10): 22-24, 28.
[19] 李传宗. 谷子苗期黄叶性状的生理基础及候选基因鉴定[D]. 北京: 中国农业科学院, 2020.
[20] 谢宁昆. 水稻叶色突变体生物学特性分析与基因定位研究[D]. 长沙: 湖南师范大学, 2020.
[21] Tang Y L, Wu X L, Li C S, et al. Yield, growth, canopy traits and photosynthesis in high-yielding, synthetic hexap-loid-derived wheats cultivars compared with non-synthetic wheats[J]. Crop and Pasture Science, 2017, 68(2): 115-125.
[22] Yang Y Z, Zhang Z H, Wang G X, et al. Photosynthesis, growth, and yield of Paeonia ostii in tree-based agroforestry systems[J]. Photosynthetica, 2019, 58(1): 29-36.
[23] Li Y j, Ma L S, Wu P, et al. Yield, yield attributes and pho-tosynthetic physiological characteristics of dryland wheat (Triticum aestivum L.) / maize (Zea mays L.) strip intercrop-ping[J/OL]. Field Crops Research, 2020, 248(1): https:// doi.org/ 10.1016/j.fcr.2019.107656.
[24] Zhao D L, Barry G, Jack C C. Sugarcane leaf photo-synthesis and growth characters during development of water-deficit stress[J]. Crop Science, 2013, 53(3): 1066-1075.
[25] 阿布都克尤木·阿不都热孜克, 高 强, 徐 麟, 等. 遮蔭对橡胶草生理特性的影响[J]. 新疆农业科学, 2020, 57(11): 2126-2134.
[26] 王灵燕. 油菜素内酯调控拟南芥黄化幼苗转绿的分子机制研究[D]. 济南: 山东大学, 2020.
[27] 田月月. 黄金芽茶树叶色响应光质的生理特性及机制研究[D]. 泰安: 山东农业大学, 2020.
[28] 丁永强. 茉莉酸和光照调控烟草叶片黄化的作用机制分析[D]. 重庆: 西南大学, 2017.
[29] 李 根. ‘红绵蜜柚’生长素诱导相关基因CgPCNT115的克隆及功能分析[D]. 雅安: 四川农业大学, 2019.
[30] 贾 兵, 郭国凌, 王友煜, 等. ‘黄金梨’缺铁黄化叶片受GA3诱导复绿的机理研究[J]. 园艺学报, 2021, 48(2): 254-264.
[31] Ranwala A P, Miller W B, Kirk T I, et al. Ancymidol drenches, reversed greenhouse temperatures, postgreenhouse cold storage, and hormone sprays affect postharvest leaf chlorosis in easter lily[J]. Journal of the American Society for Horticultural Science American Society for Horticultural Science, 2000, 125(2): 248-253.
[32] Han Z H, Han C Q, Xu X F, et al. Relationship between iron deficiency stress and endogenous hormones in iron-efficient versus inefficient apple genotypes[J]. Journal of Plant Nutrition, 2005, 28(11): 1887-1895.
[33] Chen J, Zhang N N, Pan Q, et al. Hydrogen sulphide alle-viates iron deficiency by promoting iron availability and plant hormone levels in Glycine max seedlings[J]. BMC Plant Biology, 2020, 20(1): 417-525.
[34] Takeshi T. Different chlorosis types in the leaf and stem of Hydrilla; hormone effect in culture solution[J]. The Botani-cal Magazine Tokyo, 1953, 66(775/776): 13-17.
[35] 禤 哲. 基于转录组测序的槟榔叶片黄化分子机理研究[D]. 海口: 海南大學, 2019.
责任编辑:谢龙莲
猜你喜欢
江苏农业科学(2016年8期)2017-02-15 19:34:32
江苏农业科学(2016年8期)2017-02-15 19:34:32
山东农业科学(2016年12期)2017-01-21 15:13:33
山东农业科学(2016年12期)2017-01-21 15:09:01
山东农业科学(2016年12期)2017-01-21 14:59:11
长江蔬菜·学术版(2016年12期)2017-01-12 20:57:59
现代农业科技(2016年19期)2016-12-12 15:25:19
现代农业科技(2016年19期)2016-12-12 15:16:23
农业与技术(2016年19期)2016-12-12 03:13:59
中国中药杂志(2016年20期)2016-11-19 12:30:29
