
黄土高原刺槐人工林林下植物多样性对模拟降水变化的响应
2021-01-12 03:28:20时新荣袁志友
水土保持研究 2021年1期
邓 强, 刘 迪, 时新荣, 袁志友
(1.中国科学院 水利部 水土保持研究所 黄土高原土壤侵蚀与旱地农业国家重点实验室,陕西 杨凌 712100; 2.中国科学院大学, 北京 100049; 3.西北农林科技大学 林学院,陕西 杨凌 712100; 4.西北农林科技大学 黄土高原土壤侵蚀与旱地农业国家重点实验室, 陕西 杨凌 712100)
以气候系统变暖和降水格局(降水量、降水间隔、降水季节分布和降水年际变异)变化为主要特征的全球气候变化已经成为国际公认的事实。IPCC第五次评估报告指出,1880—2012年全球海陆表面平均温度已上升0.85℃,北半球中纬度陆地降水呈增加趋势,极端天气和气候事件(如干旱、热浪、强降水)发生的频率和强度在增加[1]。众所周知,水分是植物体的重要组成部分,也是植物养分循环和流动的载体,在土壤—植被—大气系统物质与能量转化中起着核心和纽带的重要作用。未来气候变化导致的降水量变化将会对生态系统的结构、功能与过程产生深刻影响[2]。物种多样性是表征和评价生态系统结构和功能的重要指标,研究物种多样性对降水量变化的响应规律对未来气候变化下的植被建设和生态恢复具有重要意义。
由于降水格局、生态系统类型、土壤类型和物种组成等因素的差异,降水量变化对物种多样性的影响各不相同。如研究表明物种多样性与多年平均降水量在全球尺度上呈显著正相关关系,降水量增加允许更多物种在生理上耐受环境并达到最小存活种群来提高物种丰富度[3]。对于水分为主要限制因子的生态系统,适度增加降水,更适宜的生境条件可以供更多的生命体生存,缓解种内和种间竞争,提高群落的物种丰富度、均匀度和多样性[4]。降水主要是通过改变土壤中的水分含量来间接影响植物群落,它具有一定的滞后效应和累积的效应,因此降水量短暂增加或单个湿润年对群落物种多样性的影响有限[5]。Adler等[3]对美国堪萨斯州高草草原连续36 a的物种丰富度和降水量之间的关系进行了研究,结果表明单一年份的降水量与物种丰富度的相关性不强,物种丰富度在干旱年之后的湿润年里增加最多。降水变化也会导致物种种间关系的变化,导致群落优势种的更替。Gherardi等[6]研究发现,长期干旱会导致美国西部荒漠草原灌木入侵,草本植物多度下降。在某些地区,降水量增加可能会通过某些形式的竞争性排斥导致物种多样性下降:最适宜高水分条件的物种垄断了有限的资源,从而遏制了其他物种的侵入和正常生长。前人研究表明,极端降水和极端干旱均会对植物多样性产生严重的影响,导致群落物种多样性显著下降[7]。另有一些研究表明,降水量变化对物种多样性影响不显著。研究发现,降水处理对青藏高原高寒草原群落物种多样性无显著影响,证明水分并不是该植物群落的主要限制因子[8-9]。上述结果说明,由于降水变化程度和生态系统的巨大差异,物种多样性对降水变化的响应并不存在一致的规律。
降水变化对物种多样性的影响作为目前全球变化生态学热点问题,全球范围内已有大量研究,但主要集中于草地生态系统,关于森林生态系统物种多样性对降水变化尤其是极端降水量变化的响应研究仍然较少。
刺槐(RobiniapseudoacaciaLinn.)具有耐旱、耐贫瘠、生长迅速、根蘖性好、易繁殖等特点,是黄土高原广泛种植的造林树种之一。但是在造林过程中存在树种选择单一,轻视林下草本和灌木的问题,一些林分的林冠郁闭后,林下层植被盖度低,其水土保持和水源涵养能力大打折扣[10]。研究表明,林下层植被(主要是草本和灌木)虽然只占森林生态系统生物量的很少的一部分,但是生物量和养分的周转速率比乔木树种快得多,同时具有涵养水源、保持水土的重要作用[11]。同时,由于黄土高原独特的植被和土壤条件,林下层植被对气候变化和人为干扰非常敏感[12]。目前,关于全球变化背景下黄土高原生态系统水文过程、生产力、碳循环、演替等方面已有大量研究,但是对于人工林林下植被的多样性、适应性及演替对降水量变化的响应研究较少。因此,开展黄土高原刺槐人工林林下植被物种多样性对降水变化的响应研究,对于提高黄土高原植被恢复效益、维护生物多样性及提高生态系统稳定性等方面具有重要意义。本研究以黄土高原35 a生刺槐人工林林下植被为研究对象,进行野外降水控制试验,旨在研究:(1) 土壤含水量对9个降水量变化强度的响应规律;(2) 群落物种组成和结构对降水量变化的响应规律;(3) 群落物种多样性对降水量变化的响应规律;(4) 探索土壤含水量和群落物种多样性的相关性。
1 材料与方法
1.1 研究区概况
研究区域位于陕西省咸阳市永寿县槐平林场,地处渭北黄土高原南缘与关中平原的交汇地带,属六盘山支脉千山余脉,为黄土高原丘陵沟壑地貌,地理位置为107°56′—108°20′E,34°29′—34°59′N,海拔1 116~1 505 m。该区属于暖温带大陆性季风气候,年平均气温10.8℃,年平均降水量570.1 mm,无霜期210 d,试验期间月平均降雨量和月平均气温见图1。土壤类型主要是灰褐土和黑垆土,呈中性、微碱性,pH值为6~8.5。研究区处于暖温性森林地带向暖温性森林草原地带的过渡地带,原始森林植被破坏殆尽,残存有少量辽东栋、白桦、侧柏等天然次生林,现多为人工种植而形成的次生植被[13]。槐平林场现有林地面积1.33万hm2,其中人工造林1.07万hm2,造林树种以刺槐(Robiniapseudoacacia)和油松(Pinustabulaeformis)为主。刺槐人工林林下主要植物有蛇莓(Duchesneaindica)、山莓(Rubuscorchorifolius)、达乌里胡枝子(Lespedezadavurica)、黄花苜蓿(Mcdicagofalcata)、艾蒿(Artemisiaargyi)、黄花蒿(Artemisiaannua)、蒲公英(Taraxacummongolicum)、早熟禾(Poaannua)、鹅观草(Roegneriakamoji)、赖草(Leymussecalinus)、大披针薹草(Carexlanceolata)、宽叶薹草(Carexsiderosticta)、茜草(Rubiacordifolia)、老鹳草(Geraniumwilfordii)等。
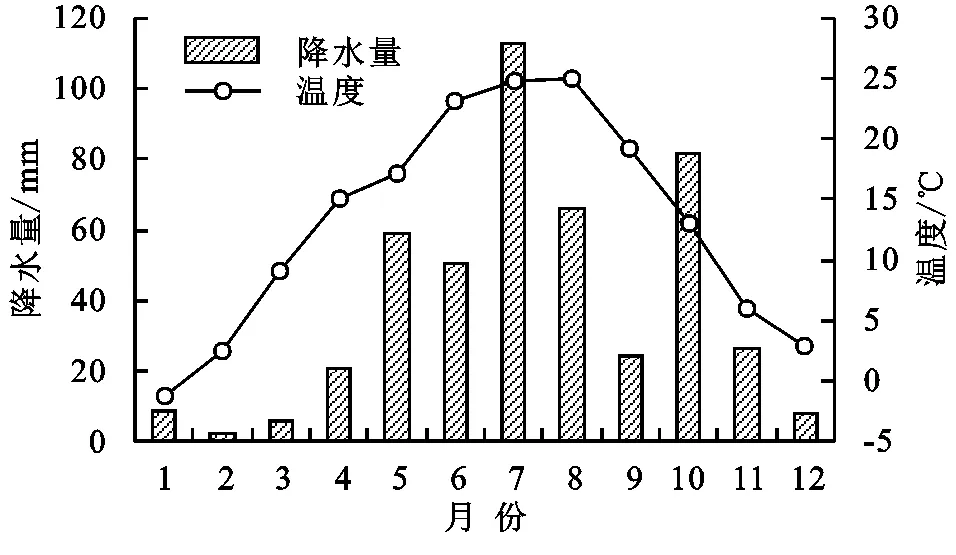
图1 试验期间研究区月降水量和气温变化
1.2 试验设计
本试验于林龄35 a生的刺槐人工林中进行,选取立地条件相似、同时远离道路、人为干扰较少的3块130 m ×60 m样地,样地间隔大于1 km,样地概况见表1,样地布局见图2。每块样地设置9块10 m ×10 m样方,采用随机区组设计,共设置27块样方,样方间隔大于10 m,分别设置9种处理:P80-(拦截穿透雨面积为林地投影面积的80%)、P60-(拦截穿透雨面积为林地投影面积的60%)、P40-(拦截穿透雨面积为林地投影面积的40%)、P20-(拦截穿透雨面积为林地投影面积的20%)、CK(对照)、P20+(施加P20-样地拦截的穿透雨)、P40+(施加P40-样地拦截的穿透雨)、P60+(施加P60-样地拦截的穿透雨)、P80+(施加P80-样地拦截的穿透雨)。通过安装遮雨棚来控制降水,遮雨材料采用透光率90%的聚乙烯薄膜(图2)。在每块减雨样方中,依据减雨比例铺设若干条0.4 m×10 m的聚乙烯薄膜于木架上用于收集穿透雨,减雨薄膜一端离地2 m,另一端离地1 m。在遮雨棚低端铺设聚氯乙烯水管,用于将收集到的穿透雨转移出样方。根据气象资料计算,次降雨P80-拦截的穿透雨量最大不超过1 m3,因此减雨样方外挖出开口1 m×1 m深1 m的集水坑,并设计有盖子,用于存储转移出样方的降水。每次降水结束后,人工将集水坑里的雨水均匀地转移到相同比例的增雨样方中。所有减雨薄膜于生长季开始前(2016年4月)首次布设,生长季结束后(2016年10月)移除,试验进行期间,每周将落于减雨薄膜的凋落物收集并均匀地撒回样地。为阻断样方内外的地表径流和壤中流,于2016年3月沿每块样方的四周嵌入深度0.5 m露出地面0.2 m的聚氯乙烯隔水板。同期,在试验区内架设FY-QBX型自动气象站(武汉富源飞科电子科技有限责任公司)对大气降水量等气象参数进行观测。

表1 刺槐人工林样地概况
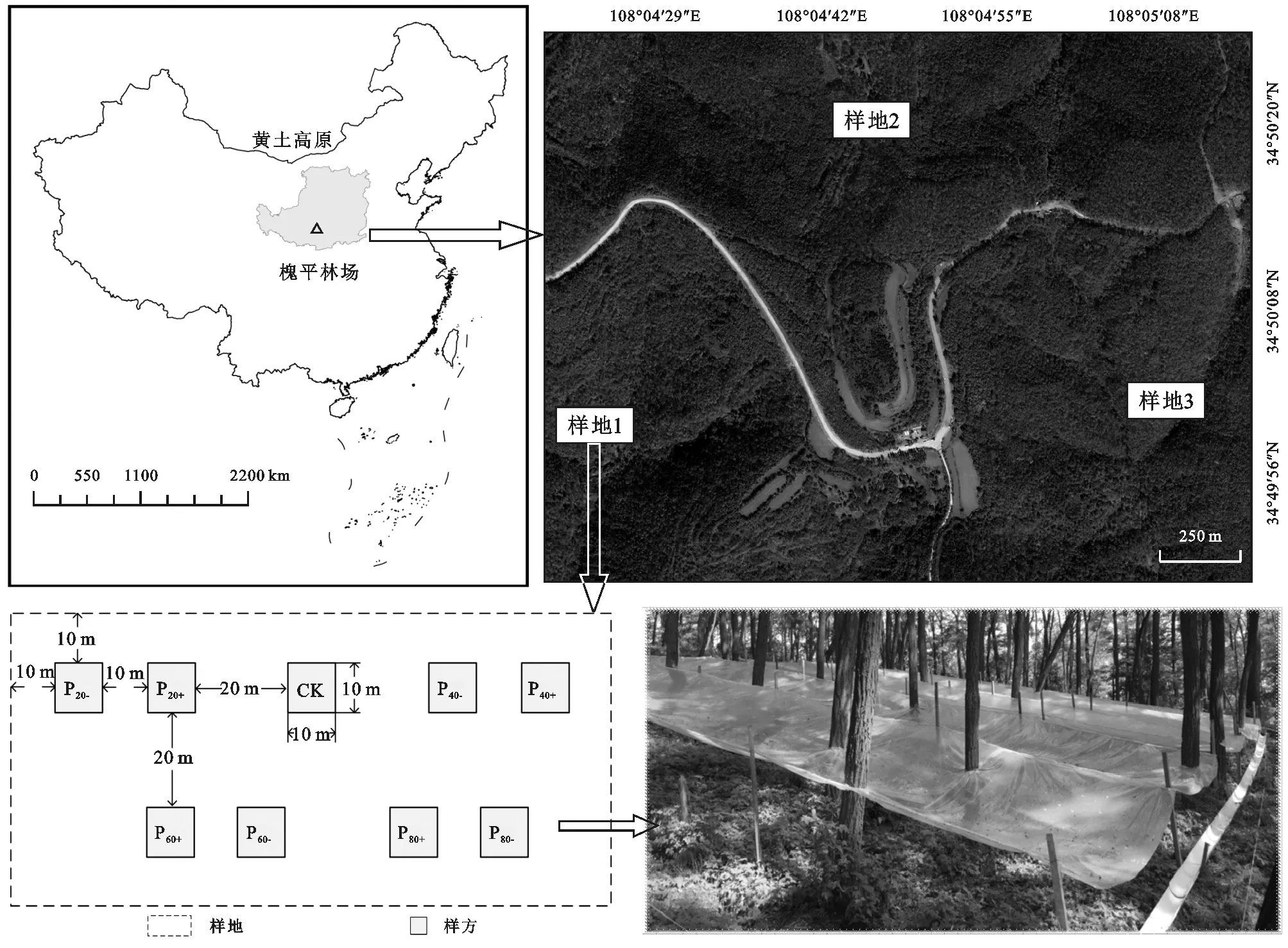
注:P80-,P60-,P40-,P20-分别为拦截林地投影面积80%,60%,40%,20%的穿透雨处理,P80+,P60+,P40+,P20+分别为施加P80-,P60-,P40-,P20-样地拦截的穿透雨处理,CK为对照,下同。图2 研究区域与试验布设示意图
1.3 数据收集
于2016年8月进行群落调查,在每个处理样地内随机选择5个50 cm×50 cm的样方,记录每个样方中出现的物种并测量盖度和多度,记录优势种。然后沿每块样地坡面的上、中和下部随机选择5点用土钻取样,分层采集0—20 cm,20—40 cm的土样并混合,每个样地重复3次,用烘干法测定土壤质量含水量。
1.4 数据计算与统计分析
(1) 物种重要值
重要值是研究某个物种在群落中的重要性的综合数量指标[14],采用如下公式:
IV=(RA+RF+RC)/3
式中:Ⅳ为重要值;RA为相对多度,即某个种的个体数占全体物种个体数之和的百分比;RF为相对频度,即某个种出现的样方数占所有种出现的样方数之和的百分比;RC为相对盖度,即某个种的分盖度占所有种的分盖度之和的百分比。
(2) 物种丰富度指数
Patrick指数(R):R=S
Margalef指数(Ma):Ma=(S-1)/lnN
式中:S是样方内物种数;N为样方中物种个体总数。
(3) 物种多样性指数
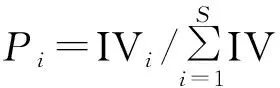
式中:Pi为第i个物种的相对重要值;Ⅳi为第i个物种的重要值。
(4) 物种均匀度指数
Pielou均匀度指数(E):E=H′/lnS
运用R(4.0.2)进行数据的整理与分析,采用单因素方差分析(One-way ANOVA)和Duncan多重比较法检验9种降水处理之间土壤含水量、物种丰富度、多样性和均匀度指数差异显著性(ɑ=0.05),采用线性回归分析分析土壤含水量和物种丰富度、多样性、均匀度指数的相关性。
2 结果与分析
2.1 降水处理对土壤含水量的影响
随着降水减少—对照—降水增加处理梯度,土壤含水量呈增加趋势,降水处理对不同土层土壤含水量的影响存在差异(表2)。0—20 cm土层,P80-土壤含水量最小,为16.98%,显著低于对照(p<0.05),P60+,P80+土壤含水量分别为25.59%,28.05%,显著高于对照,其他降水处理与对照差异不显著;20—40 cm土层,P80-,P60-土壤含水量分别为14.69%,15.45%,显著低于对照,P60+,P80+土壤含水量分别为18.65%,19.51%,显著高于对照,其他降水处理之间差异不显著。同一降水处理下,随着土层的增加,土壤含水量呈下降趋势。除P80-外,其他降水处理0—20 cm土层土壤含水量显著高于20—40 cm土层。

表2 不同降水处理下不同土层土壤含水量
2.2 降水处理对群落物种组成和结构的影响
由表3所示,刺槐人工林样地林下灌草层的分层现象不明显,灌木层物种数和个体数较少,主要有蔷薇科的蛇莓和山莓、豆科的达乌里胡枝子等。林下层共出现31种植物,分属15个科,优势种主要出现在蔷薇科、豆科、菊科和禾本科,这4科物种占总物种数的58.06%。其中,对照样地林下层出现植物23种,群落优势种为早熟禾、鹅观草和艾蒿;P20-林下层出现植物27种,数量最多,群落优势种为山莓和早熟禾;P80-林下层共有20种植物,数量最少,群落优势种为山莓和艾蒿。
不同降水处理下刺槐人工林林下植物群落组成和结构发生了明显变化(表3,图3)。随着降水减少—对照—降水增加处理梯度,蔷薇科植物重要值呈先减少后增加趋势,最大值和最小值分别为29.87(P60-)和7.29(P40+),并且优势种由山莓逐渐变为山莓和蛇莓,与对照相比,降水减少和P60+,P80+显著提高了蔷薇科植物重要值;豆科植物优势种为达乌里胡枝子,与对照相比,P80-,P60-,P40-,P60+,P80+显著降低了豆科植物重要值,其他处理间差异不显著,最大值和最小值分别为11.32(CK)和0.05(P80-);随着降水减少—对照—降水增加处理梯度,菊科优势种由艾蒿逐渐变为艾蒿和黄花蒿,与对照相比,P80-,P60-,P40-显著降低了菊科植物重要值,P60+显著提高了菊科植物重要值,其他处理间差异不显著,最大值和最小值分别为40.76(P60+)和11.06(P60-);禾本科优势种为早熟禾和鹅观草,与对照相比,P80-,P60+,P80+显著降低了禾本科植物重要值,P20+显著提高了禾本科植物重要值,其他处理间差异不显著,最大值和最小值分别为36.17(P20+)和11.87(P80+);降水处理对杂类草物种组成和重要值影响较大,随着降水减少—对照—降水增加处理梯度,杂类草植物重要值呈先减少后增加趋势,最大值和最小值分别为33.99(P80+)和16.57(P20-),P80-和P60-大披针薹草和宽叶薹草消亡,紫花地丁、灰绿藜、鹅绒藤、马蔺重要值增加,杂类草植物重要值显著高于对照,降水增加,大披针薹草逐渐为杂类草优势种,P40+,P60+,P80+杂类草植物重要值显著高于对照。
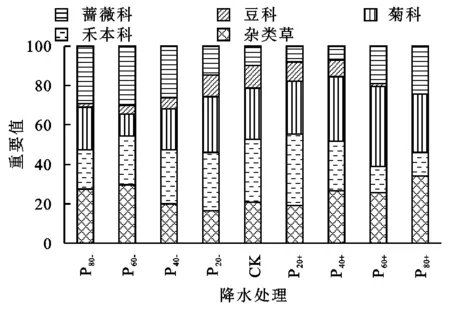
图3 不同降水处理样地林下植物群落物种重要值
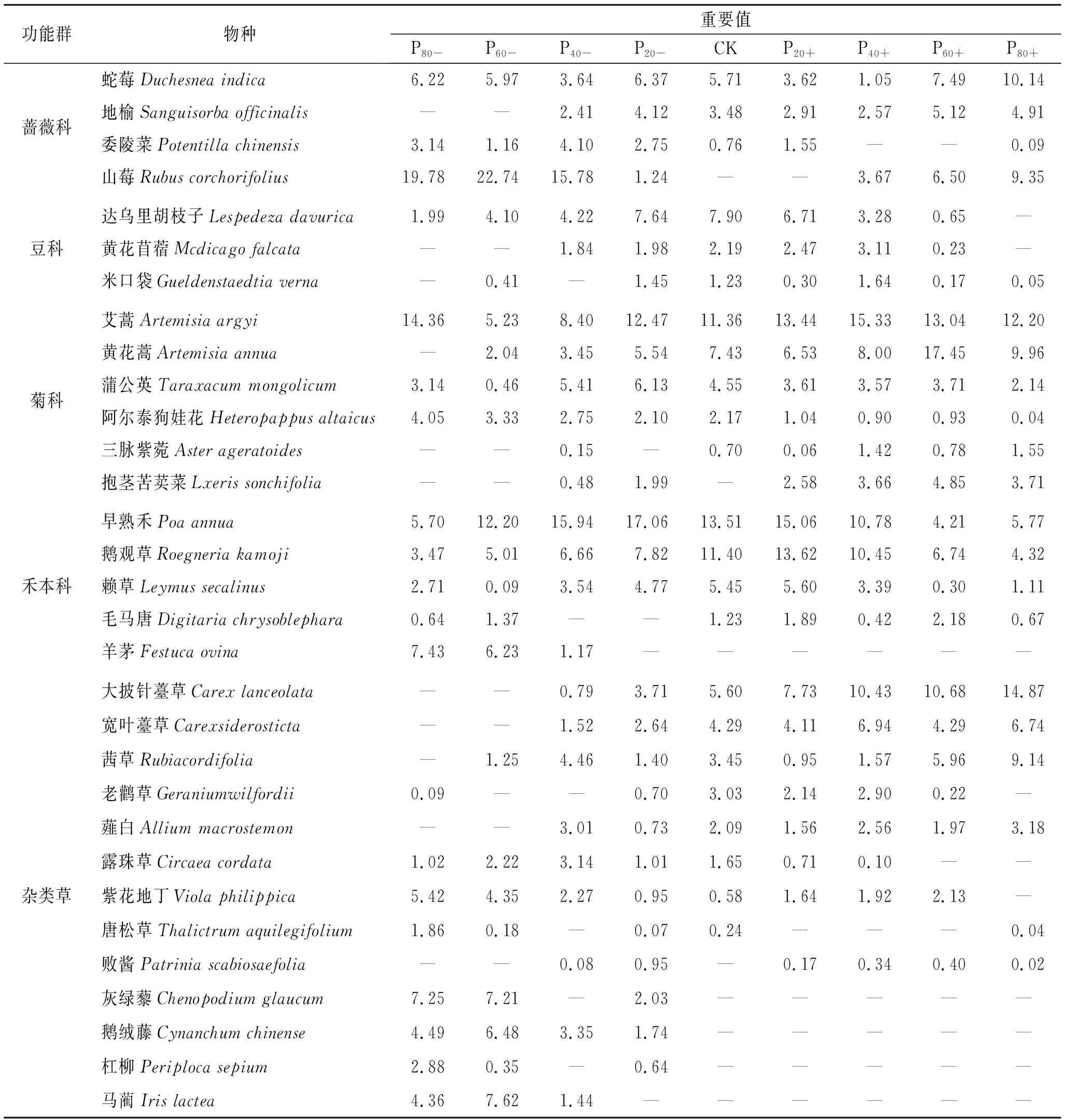
表3 不同降水处理下林下植物群落物种组成和重要值
2.3 降水处理对群落物种多样性的影响
群落物种丰富度在不同降水处理间存在差异(图4)。其中,对照样地Patrick指数为23,P80-,P60-,P80+Patrick指数低于对照,P40-,P20-,P20+,P40+Patrick指数高于对照,P60+Patrick指数和对照相同。随着降水减少—对照—降水增加处理梯度,Margalef指数呈先上升后下降趋势,最大值出现在P20-。与对照相比,P80-和P80+显著降低了Margalef指数,其他降水处理对Margalef指数影响不显著。
2种群落物种多样性指数随降水处理的变化规律基本一致(图4)。随着降水减少程度的增加,Simpson指数和Shannon-Weiner指数逐渐下降,P80-和P60-物种多样性指数显著低于对照;随着降水减少程度的增加,Simpson指数和Shannon-Weiner指数先上升后下降,在P20+处达到峰值;与对照相比,P20+和P40+显著增加了Simpson指数,P20+显著增加了Shannon-Weiner指数,P80+显著降低了Simpson指数,P60+和P80+显著降低了Shannon-Weiner指数。
随着降水减少程度的增加,Pielou均匀度指数逐渐下降,P80-,P60-,P40-显著低于对照(图4);随着降水减少程度的增加,Pielou均匀度指数先上升后下降,在P20+处达到峰值,P40+与对照差异不显著,P60+和P80+显著降低了Pielou均匀度指数。

注:图表中数据均为平均值±标准误。不同小写字母表示不同降水处理之间差异显著(p<0.05)。图4 不同降水处理样地林下植物群落物种多样性
2.4 群落物种多样性与土壤含水量的相关性
群落物种丰富度、多样性和均匀度指数随土壤含水量增加呈先上升后下降趋势(图5)。群落物种丰富度、多样性和均匀度指数与0—20 cm土层土壤含水量呈显著正相关关系;Simpson指数与20—40 cm土层土壤含水量呈显著正相关关系(R2=0.630,p<0.05);Margalef指数、Shannon-Weiner指数和Pielou均匀度指数与20—40 cm土层土壤含水量无显著相关关系(p>0.05)。
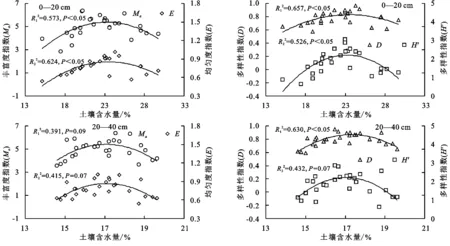
图5 群落物种多样性与土壤含水量的关系
3 讨 论
3.1 降水处理对群落物种组成和结构的影响
我们的研究表明,生长季林内穿透雨增加、减少处理显著改变了黄土高原刺槐人工林土壤水分含量,0—20 cm土层土壤含水量的变化相对于20—40 cm土层更加明显,与前人的研究结果一致[12,15]。研究区刺槐人工林林下灌草层的分层现象不明显,主要原因是造林时经过全面整地,天然灌草被破坏,林冠郁闭后,林下层的光照量不足,使得灌木层和草本层的生长受到了抑制[10,13]。
林下层植被主要由蔷薇科、豆科、菊科和禾本科植物构成,植物水分生态类型以中生、中旱生植物占主要优势,符合黄土高原半干旱区的植被组成特征,与郭琳等的研究结果一致[13]。降水减少和增加处理都会提高蔷薇科物种重要值,表现出较强的适应性。优势种蛇莓和山莓都属于耐旱、耐贫瘠、适应性强的物种,并且具有蔓生性,形态为矮小的灌木覆被在地面上,并且具有强大的根系,使其能很好地适应极端水分条件[16]。豆科植物优势种为达乌里胡枝子,极端降水处理显著降低了豆科物种重要值,这与段东平在黄土丘陵区天然草地群落进行的人工补水试验结果一致[17]。可能的原因是,相对于其他草本植物,达乌里胡枝子根系较深,主要利用较深层土壤水分,土壤表层增加的降水对其影响有限,并且达乌里胡枝子为中旱生植物,极端干旱环境不利于其生长[17-18]。菊科优势种主要为艾蒿和黄花蒿,都属于中生性蒿属植物,相对于分布在黄土高原森林草原和典型草原地带旱生性的铁杆蒿(Artemisiagmelinii)和茭蒿(Artemisiagiraldii),更容易受到干旱胁迫的影响。因此,降水减少处理显著降低了菊科物种重要值[19]。早熟禾和鹅观草属于中生植物,生态位较宽,但是过分的干旱胁迫和水涝均不利于其生长,因此极端降水处理显著降低了禾本科物种重要值[20]。与对照相比,降水增加处理提高了杂类草物种重要值,这与Miao等[21]对青藏高原东北缘草地植物群落的研究结果一致,说明群落中地位较低的伴生种能够在外界环境改善的条件下充分利用生长繁殖所需的资源。大披针薹草和宽叶薹草是簇丛型中生植物,增加降水显著提高了其重要值。寇萌等[22]对黄土高原延河流域植物群落特征的研究发现,大披针薹草是降水量较高的中南部区域草本群落的优势种。降水减少,杂类草物种重要值增加,这与张志南等[23]对黄土高原半干旱区天然草地群落的研究结果一致。灰绿藜、鹅绒藤、杠柳、马蔺这4种植物为研究区林下层植被偶见种,仅出现在降水减少处理样地,并且随着降水量下降逐渐成为群落优势种。灰绿藜、杠柳、马蔺都属于耐旱、耐盐碱植物,具有较宽的生态位,在极端干旱胁迫下仍能生存[24-25]。
3.2 降水处理对群落物种多样性的影响
物种多样性是表征和评价植物群落组成和功能的重要指标。本研究中,除P80-,P60-,P60+和P80+外,其他降水处理物种数都高于对照,符合Connell[26]的“中度干扰”假说,即中等程度的干扰使竞争替代减弱,从而维持较高的物种多样性。Grime[27]指出,环境可以通过两种不同的方式影响物种丰富度,一是影响物种优势度;二是影响潜在丰富度(物种库)。本研究中,降水变化促进了多年生草本植物和灌木的萌发。前人研究表明,降水量增加或减少都可能打破土壤潜在种子库中部分种子休眠,促使其萌发,进而增加物种数[28-29]。不同降水处理下Margalef指数变化规律与Patrick指数略有不同,表明降水处理后物种数增加,但是样方内物种个体总数下降。刺槐人工林林下物种多样性指数和均匀度指数随降水处理表现出基本一致的变化规律。物种多样性随降水量的减少呈下降趋势,一定程度的降水增加有利于物种多样性提高,过量降水显著降低物种多样性。我们的结果与黄土高原典型天然草地群落进行的相似降水变化处理试验的结果基本一致[30]。相对于20—40 cm土层,0—20 cm土层土壤含水量对群落物种多样性的影响更显著,这是由于群落主要由草本植物构成,主要利用浅层土壤的水分,深层土壤含水量变化对其影响较小。
不同生态系统物种多样性对降水变化的响应存在差异,主要是由于气候、土壤、生态系统类型、降水变化的程度和频度等多种复杂因素共同导致的。大量研究表明,降水量和物种多样性之间存在正相关关系[15,29,31-32]。降水量增加,植物能获取更多的水分用于生长和发育,缓解水分胁迫,同时能够降低群落内种群之间的生态位重叠程度,缓解种间竞争,使得同一生境能容纳更多的物种生存[4,31]。对于水分为主要限制因子的干旱、半干旱生态系统,降水量增加对物种多样性的促进作用更加明显[15,29,32-33]。但是,研究证实,过量降水会降低群落物种多样性[15,30-31,33]。这是由于过量降水会导致涝害发生,导致土壤缺氧和土壤退化,超越了植物生理耐受范围,影响植物体吸收水分和养分,进而引起植物生长发育受阻,甚至死亡[2,34]。还有一些研究发现,降水变化对群落物种多样性没有显著影响[8-9,35]。造成这种现象可能的原因,一是生态系统通过改变群落物种组成,可以抵抗一定程度的短期降水变化对物种多样性的影响[35];二是降水变异程度小,不足以对物种多样性产生显著影响[8]。前人研究证实,由于生态系统具有抵抗力稳定性,极端气候变化并不一定引起极端的生态系统反应[7,36]。Alon等[37]的研究表明,极端干旱(5—8月截雨66%,6—7月截雨100%)处理显著降低了荒漠草原沙生针茅群落优势种的丰富度和生物量,缓解了群落内的种间竞争,其他物种相对丰富度提高,从而短期内提高了物种多样性和均匀度。张蕊等[28]在内蒙古荒漠草原进行的相同极端干旱处理试验也得到了相似的结果。
姜基春等[38]研究了黄土高原4种植被带草本群落物种多样性对降水变化的响应规律,结果表明,典型草地和荒漠草地物种多样性与降水量呈正相关,森林带和森林—草原带草本群落物种多样性随降水量的增加呈下降趋势。这说明黄土高原森林林下草本植物群落受乔木与灌木的影响,群落物种多样性对降水变化的响应规律要比草地群落复杂的多。本研究发现,一年生草本植物受降水变化的影响最大,与王军锋等[32]在黄土高原荒漠草原进行的降水控制试验的结果一致。一年生草本植物根系分布较浅,相对于多年生草本植物和灌木,更容易受到浅层土壤水分波动的影响,干旱胁迫时不能利用土壤深层的水分,过量降水导致的土壤涝害对其影响也较大。前人研究发现,黄土高原刺槐人工林在成熟之后,林下层物种多样性随着林龄的增加逐渐下降,并且灌木层基本消失[13]。根据生态系统稳定性理论,生物多样性可以提高系统对干扰的抵抗力,生物多样性越高系统稳定性较高,可以一定程度的减小降水量变化对生态系统的影响[39]。因此,未来降水变化将不可避免的影响黄土高原刺槐人工林林下植物多样性,进而影响到生态系统结构和功能的稳定性。
4 结 论
随着降水量的增加,土壤含水量呈增加趋势,降水处理对0—20 cm土层土壤含水量的影响大于20—40 cm土层。不论降水减少或增加,都提高了蔷薇科和杂类草物种的重要值,但降低了豆科和禾本科物种的重要值,降水量的增加也提高了菊科物种的重要值。降水减少处理降低了物种多样性指数和均匀度指数,并且随着降水减少程度的增加呈下降趋势。增雨20%和40%增加了物种多样性指数和均匀度指数,但增雨60%和80%降低了物种多样性指数和均匀度指数。群落物种丰富度、多样性和均匀度指数与0—20 cm土层土壤含水量呈显著正相关关系。研究降水变化对物种多样性的影响对气候变化背景下的黄土高原植被建设和生态恢复具有重要意义。
猜你喜欢
作物研究(2023年2期)2023-05-28 13:44:14
现代园艺(2021年23期)2021-12-01 07:47:44
林业勘查设计(2020年1期)2021-01-18 02:40:48
新农业(2020年18期)2021-01-07 02:17:08
河南畜牧兽医(2020年21期)2020-01-10 00:20:08
公民与法治(2016年3期)2016-05-17 04:09:00
公民与法治(2016年1期)2016-05-17 04:07:56
纺织科技进展(2015年1期)2015-11-28 05:56:28
中国火炬(2015年7期)2015-07-31 17:39:57
中成药(2014年10期)2014-02-28 22:29:24
