
不同郁闭度华山松人工林林下灌木和草本多样性
2021-01-05 02:54:02欧江地地木古张时林简毅王益志
四川林业科技 2020年6期
欧江, 地地木古, 张时林, 简毅, 王益志
1. 资阳市自然资源和规划局,四川 资阳 641300;
2. 石棉县自然资源和规划局,四川 雅安 625400;
3. 金阳县脱贫攻坚综合帮扶工作队,四川 凉山 616250;
4. 四川省林业科学研究院,四川 成都 610081;
5. 四川省长江国有林保护局凉山分局,四川 凉山 615608
华山松(Pinus armandii)因其重要的生态和经济价值,是我国西南地区主要造林树种之一。但因大面积的纯林经营,华山松林出现了长势衰弱、多样性减少等诸多问题[1,2]。为提高华山松人工林生态系统的稳定性,已有学者在抚育间伐、林窗式疏伐等方面做了少量研究,但结果不尽相同[3-5]。舒圣评等[6]研究指出,通过林下补植等低效林改造措施,群落结构有所改善。林下植被具有维护森林生态功能稳定性的重要作用;郁闭度反映森林环境和结构,是森林经营的重要指标[7,8]。李双喜等[9]研究指出,林分郁闭度是影响林地光照状况的决定性因素。但当前对郁闭度的研究多在苗木更新等方面和红松(Pinus koraiensis)、马尾松(Pinus massoniana)等树种,不同郁闭度对华山松人工林林下植物的影响还不甚清晰[8,10,11]。本文通过郁闭度调控,研究华山松人工林林下灌木和草本植物的组成和多样性等变化特征,以期为华山松人工林的近自然经营提供一定理论依据。
1 材料与方法
1.1 研究区域概况
研究区域位于四川省凉山州金阳县境内(27°22′02″ — 27°56′58″N,102°56′53″ — 103°30′23″ E),邻近金沙江,属中亚热带季风气候区,气候呈立体垂直带状分布,海拔920 ~2 500 m,年均气温15.7 ℃、降水量800 mm、蒸发量146 mm、日照1 574 h,太阳辐射能441 188.78J·cm-2,无霜期约300 d;土壤以黄棕壤、紫色土为主;常见灌木有杜鹃(Rhododendron simsii)、马桑(Coriaria nepalensis)、火棘(Pyracantha fortuneana)、羊奶子(Elaeagnus pungens)、黄荆(Vitex negundo)等,草本有扭黄茅(Heteropogon contortus)、荩 草(Arthraxon hispidus)、萎 菱 菜(Potentilla chinensis)、羊茅草(Festuca)等。
1.2 样地设置及调查方法
区域华山松人工林造林方式为飞播造林,并进行一定的补植,造林密度较大,行间距约为1.5 m×1.5 m。经长期的自然或人为因素,样地现有林分郁闭度约为0.5~0.95。选择立地条件和生长状况等基本相同,且连片分布的22 年生华山松人工林为研究样地。参考前人华山松以及西南地区主要造林树种相关研究的梯度区间和郁闭程度划分[8,12],以现有林分郁闭度为基础,通过抚育修枝、均匀间伐等方式进行适当调控,形成郁闭度约为0.5、0.7、0.9 的3 个梯度,各梯度形成时间大于1 年,并分别设置3 个20 m×20 m 的重复样地,样地间隔大于20 m 且远离林缘。2020 年6 月,在各样地内根据对角线方法,选取5 个5 m×5 m 的样方调查灌木层,在其中心各选取1 个1 m×1 m 的样方调查草本层,记录每种植物的种名、高度、盖度、冠幅、株数/丛数等,记录样地海拔、坡度、坡向等基本情况(见表1)[8,13]。按照Raunkiaer 生活型分类系统进行生活型分类并计算生活型谱。
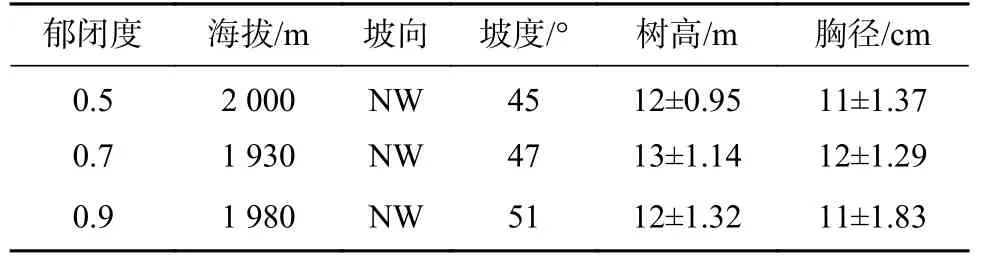
表 1 样地设置及林分状况Tab. 1 Basic information of the experimental plots and forest stands
1.3 数据处理及分析
根据调查结果,按照以下方法计算重要值、多样性指数等指标。
重要值 =(相对盖度+相对高度)/2。
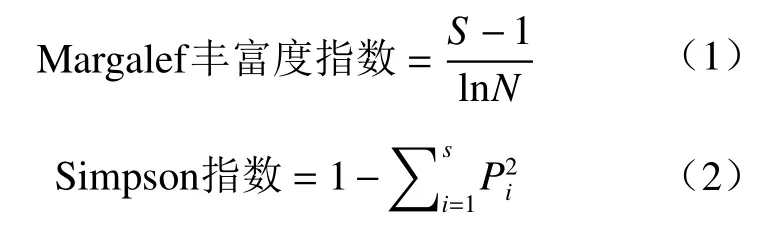
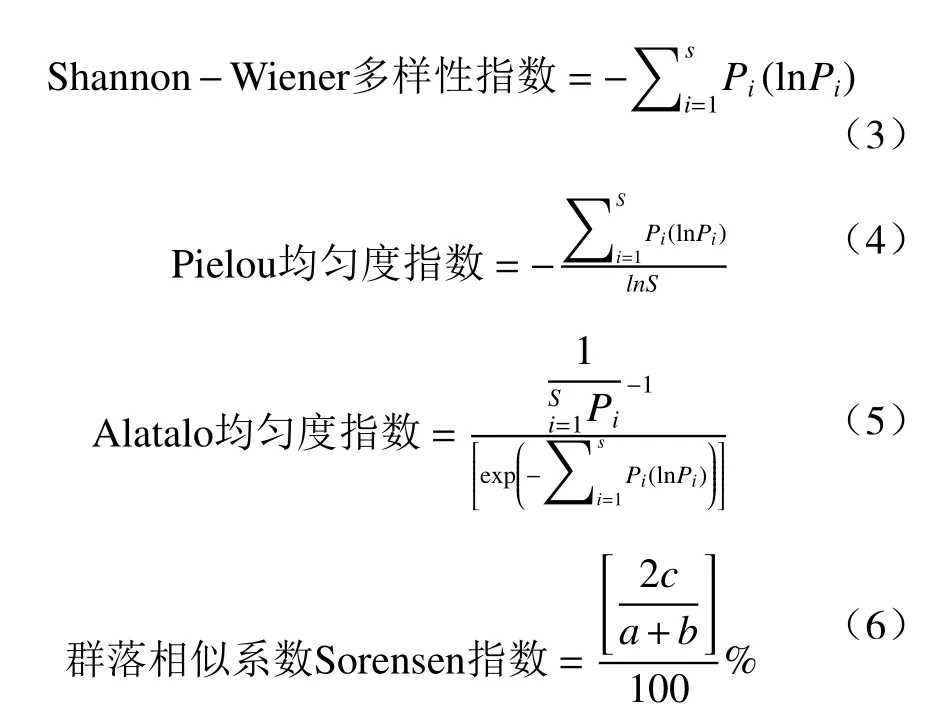
以上式中,N为调查样方中物种个体数之和,Pi为相对重要值,S为调查样方中物种数,a为某个样地的物种数,b为另一个样地的物种数,c为这两个样地的共有物种数。
采用Excel2016 和SPSS25.0 进行数据统计,Origin 2019 作图。采用单因素方差分析(One-way ANOVA)检验林下灌木和草本植物多样性在不同郁闭度间的差异。设定显著性水平α=0.05。
2 结果与分析
2.1 植物组成、生活型及重要值
植物种类共37 科54 属57 种(见表2 和表3),其中灌木15 科21 属23 种,草本22 科33 属34 种,蔷薇科(Rosaceae)植物最多,菊科(Asteraceae)、唇形科(Lamiaceae)次之。随郁闭度增大,植物种数减少,分别为46、34、30 种;0.9 郁闭度灌木植物最少,仅11 种,0.5 郁闭度草本植物最多,达到29 种。灌木层野拔子(Elsholtzia rugulosa)、小叶杜鹃(Rhododendron parvifolium)、凉山悬钩子(Rubus fockeanus)为较优势植物,草本层中蹄盖蕨(Athyrium filix-femina)、凤尾蕨(Pteris cretica)为较优势植物,但重要值均不超过0.21。由表4 可见,高位芽植物比例最高,其次是地面芽植物、地下芽植物,而高位芽植物中以矮、小高位芽居多;高位芽、地面芽、地下芽植物分别在0.7、0.9、05 郁闭度林分中比例较高。
2.2 植物丰富度指数
不同郁闭度灌木、草本和灌草物种Margalef 丰富度指数均呈显著差异(见表5,P< 0.05)。随郁闭度增大,丰富度指数明显降低(见图1),灌木、灌草在0.5、0.7 郁闭度显著高于0.9 郁闭度,草本在0.5、0.9 郁闭度间差异显著。3 个郁闭度草本植物丰富度指数均高于灌木植物。

表 2 灌木层主要物种组成及重要值Tab. 2 Species composition and important values of the main species in the shrub layer
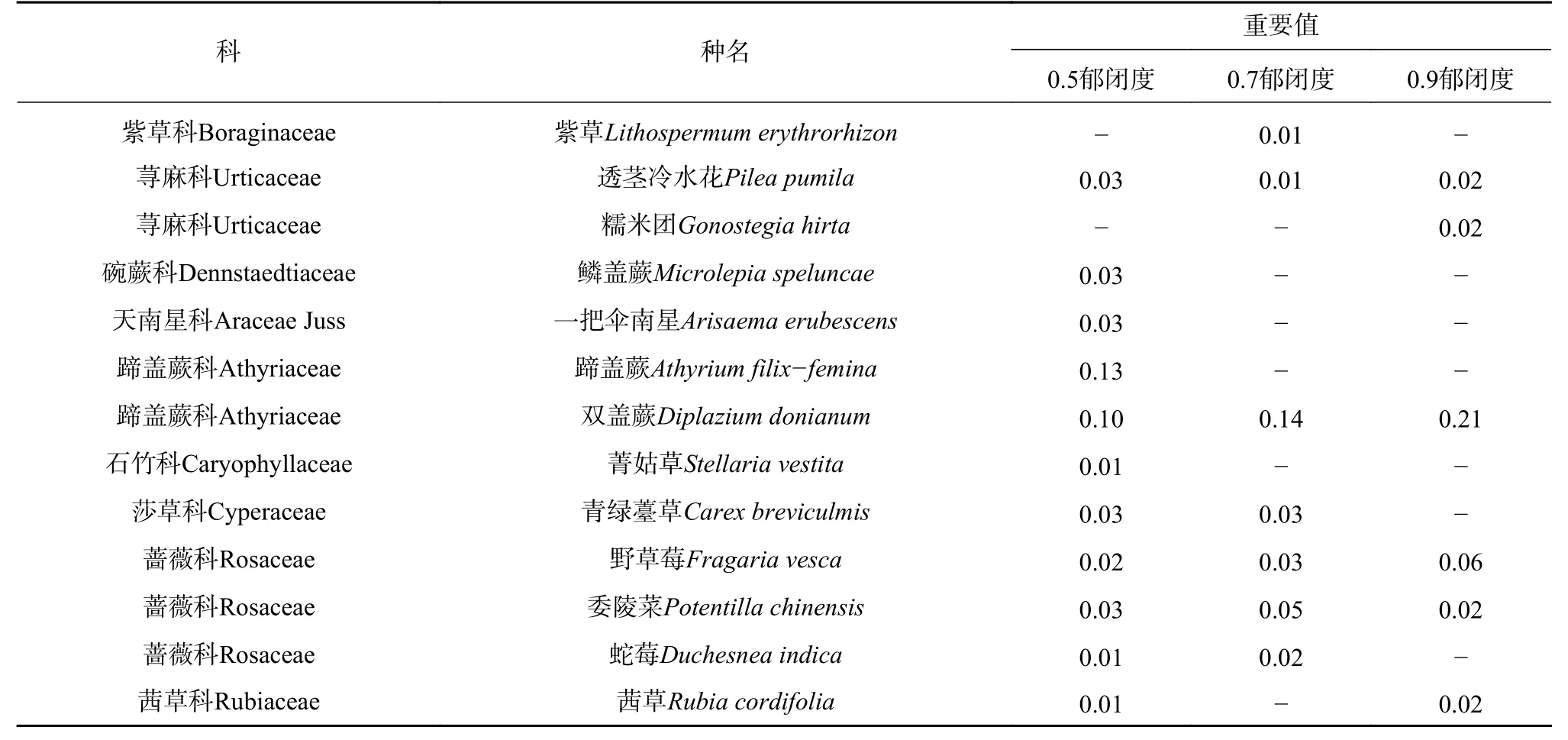
表 3 草本层主要物种组成及重要值Tab. 3 Main species composition and important values of the main species in the herbaceous layer
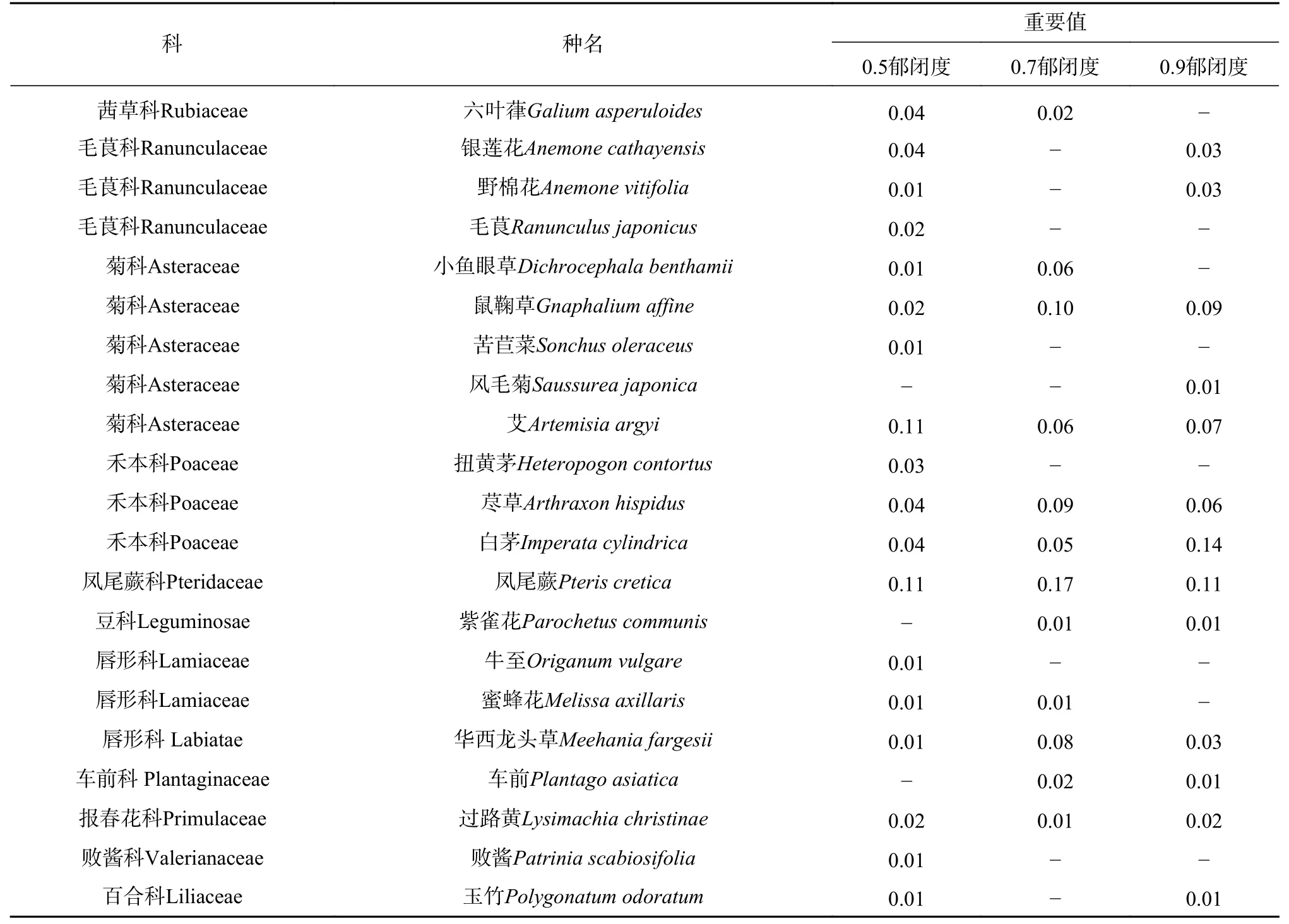
(续表 3)
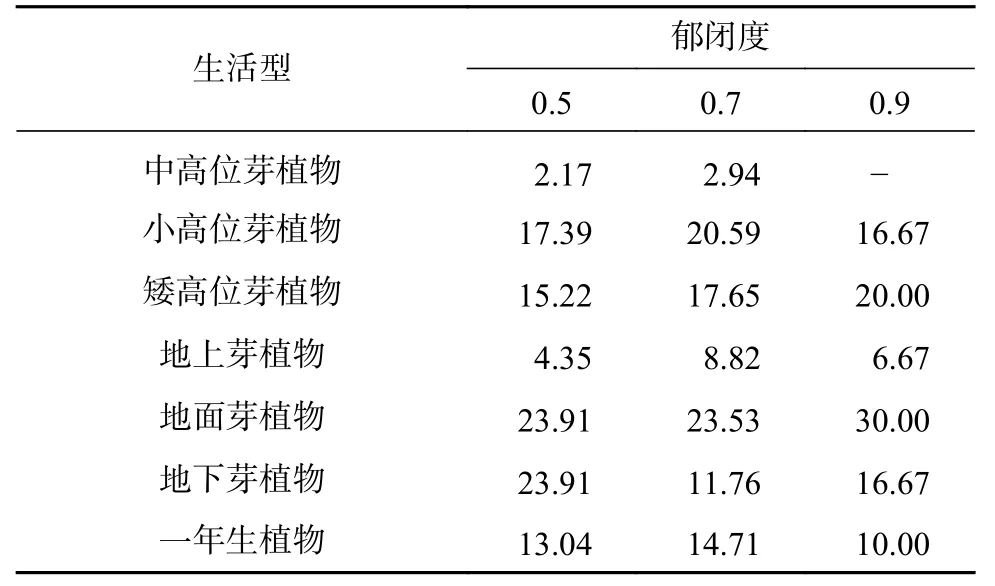
表 4 林下植物生活型的百分数Tab. 4 Proportion of life forms in understory vegetation单位:%
2.3 植物多样性指数
不同郁闭度灌木和草本植物Simpson 指数、Shannon-Wiener 指数均呈显著差异(见表5,P<0.05)。灌木和草本植物的Simpson 指数、Shannon-Wiener 指数趋势基本一致(见图2),均随郁闭度增大而降低,0.5、0.7 郁闭度的灌木和草本多样性指数均显著高于0.9 郁闭度,0.5、0.7 郁闭度间无明显差异。同时,草本植物的多样性指数高于灌木植物。
2.4 植物均匀度指数
不同郁闭度灌木和草本植物Pielou 指数、Alatato指数均不显著(见表5,P>0.05)。由图3 可知,随郁闭度增大,灌木植物Pielou 指数呈现先升后降的趋势,草本植物0.5、0.7 郁闭度基本一致,0.9 郁闭度降低;灌木植物Alatato 指数先降后升,草本与之相反。总体上,灌木植物的均匀度指数高于草本植物。
2.5 植物群落相似性
表6 显示,0.5、07 郁闭度林分植物共有种最多,为27 种,0.9 郁闭度与其他郁闭度共有种差异不大。0.7 郁闭度与其他郁闭度群落相似系数均高于65%,植被群落的生境趋于稳定。
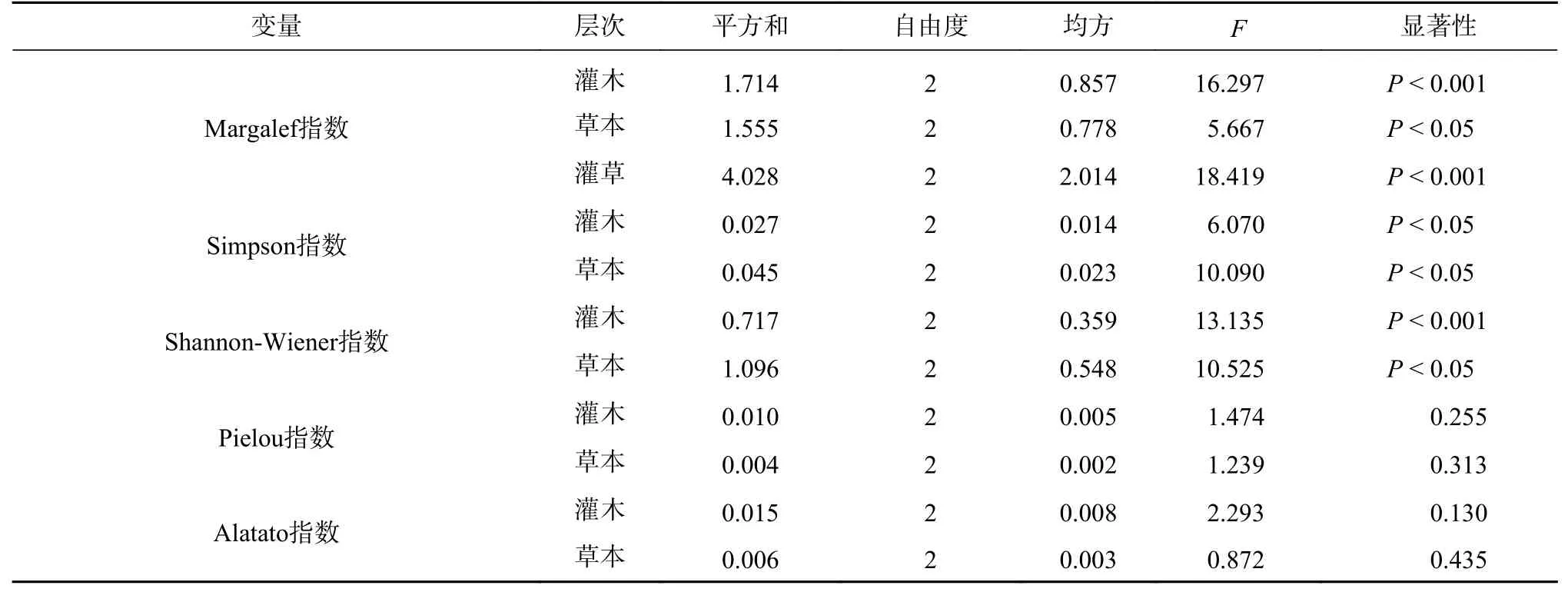
表 5 郁闭度对林下灌木和草本层多样性指数的影响Tab. 5 Variance analysis of canopy density on diversity index of understory shrubs and herbs
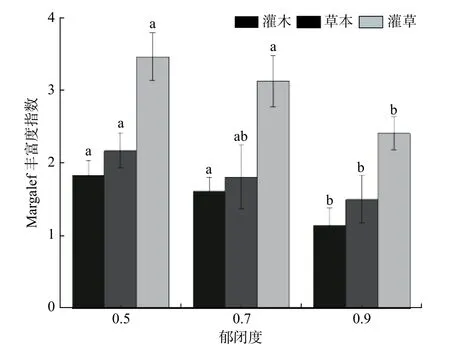
图 1 林下灌木和草本Margalef 丰富度指数。不同字母表示差异显著(P < 0.05)Fig. 1 Margalef richness index of the understory shrubs and herbs.Different letter indicates significant difference at P < 0.05
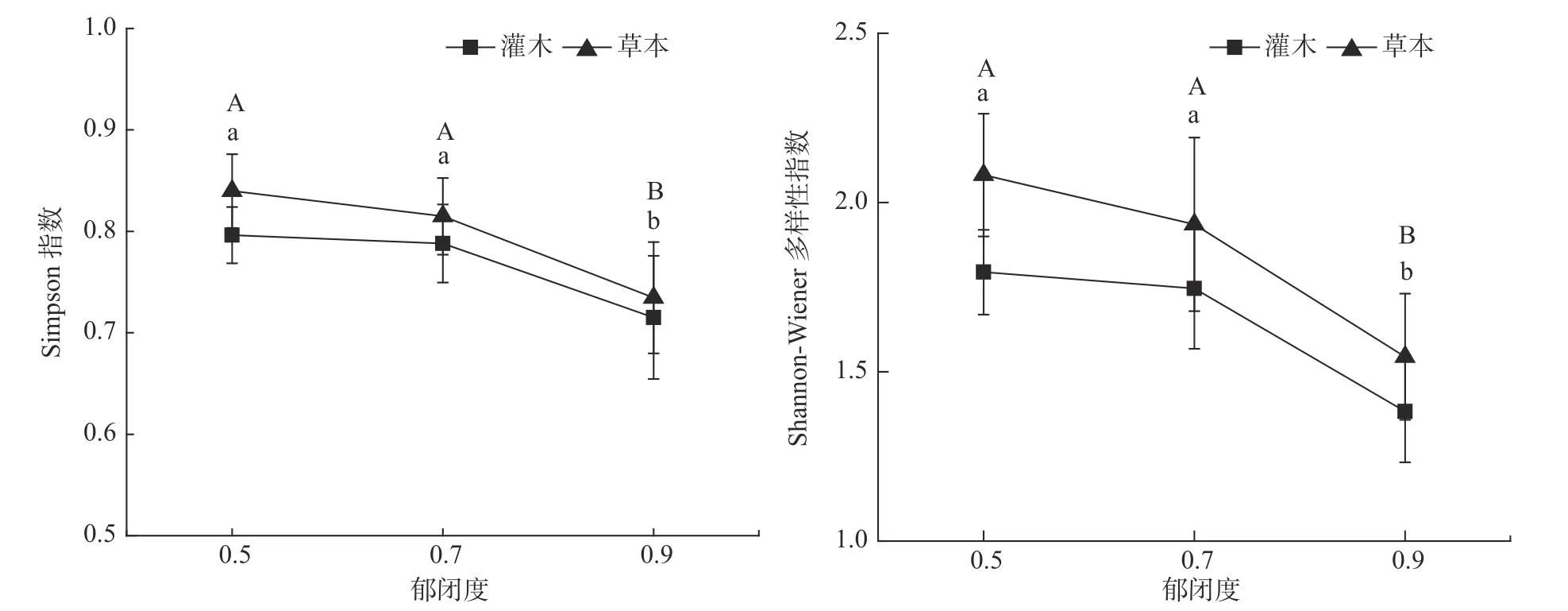
图 2 林下灌木和草本Simpson 优势度指数和Shannon-Wiener 物种多样性指数。不同字母表示差异显著(P < 0.05)。大写字母表示灌木,小写字母表示草本Fig. 2 Simpson dominance index and Shannon-wiener species diversity index of understory shrubs and herbs. Different letter indicates significant difference at P < 0.05; capital letter represents shrub; lowercase letter represents herb.
3 讨论与结论
物种组成及优势种的分布特征,决定植物群落结构类型的基本特征[7]。林下植物种类共37 科54 属57 种,蔷薇科、菊科、唇形科植物适应性较强,在各郁闭度均有分布。0.5 郁闭度林分林下植物种类远大于其他郁闭度,这与赵燕波等[8]、刘红炎等[14]研究结果相似。林冠空隙的增大,林下光照、温度、水分等条件改善,有利于植物的萌发和生长,植物种类逐渐增多[3]。各郁闭度林下植物重要值差异不大,分配较平均,最大值为0.21。已有研究指出,高郁闭度林分生境条件相对较差,种间竞争激烈,适宜生存的物种更有优势[8,13]。但本研究发现,优势种小叶杜鹃、凤尾蕨在0.7 郁闭度的重要值较高,凉山悬钩子却最低;野拔子随郁闭度增大,重要值降低。物种的适应性随林下生境的改变而不同,导致优势物种重要值存在差异[7]。本研究各郁闭度林分林下植物生活型均以高位芽植物为主,所占比例高于36%,其次是地面芽植物、地下芽植物,符合亚热带季风气候带植物的生长特征[13,15]。高位芽、地面芽、地下芽植物分别在0.7、0.9、05 郁闭度林分中比例较高。随郁闭度的增大,草本植物种类相对降低,灌木植物所占比重增大;大郁闭度林分相对更厚的凋落物层有利于地面芽植物的生存;小郁闭度林分中出现蕨类等地下芽植物更新种,其对环境因子的竞争较其他草本植物更有优势。
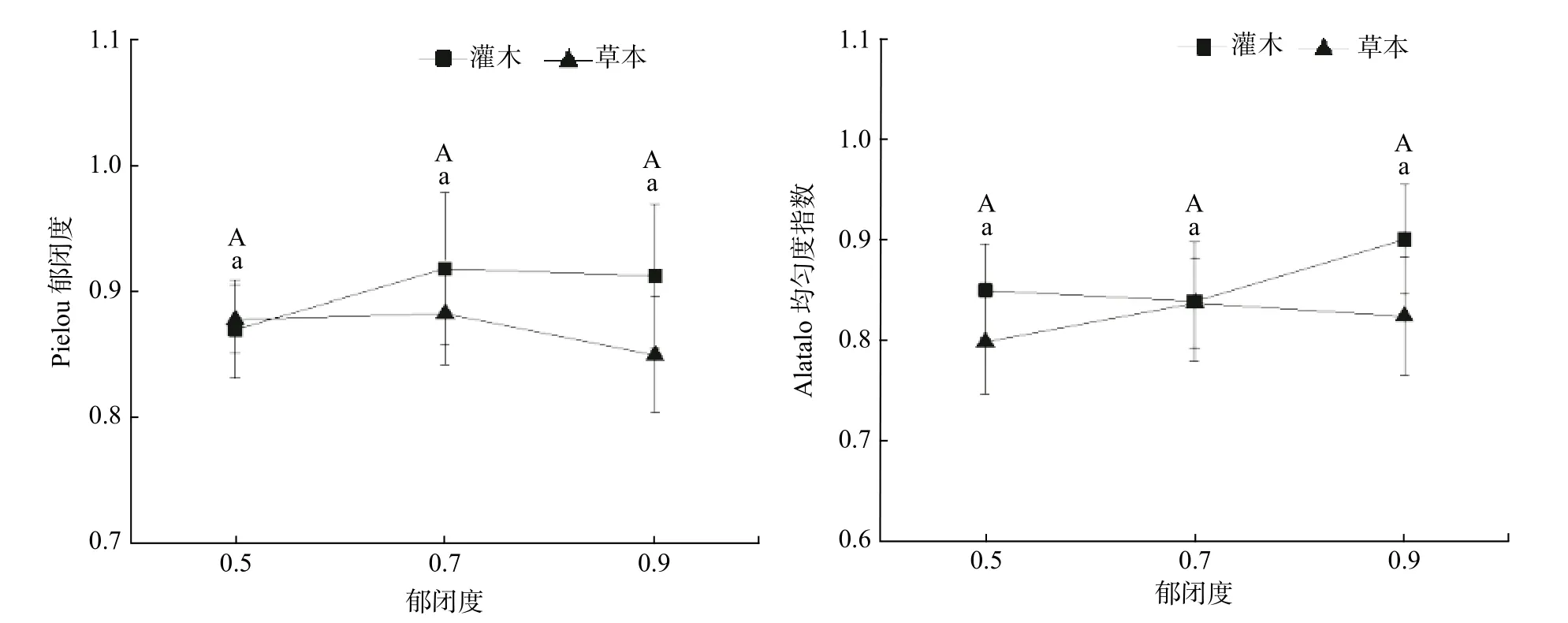
图 3 林下灌木和草本Pielou 均匀度指数,Alatato 均匀度指数。不同字母表示差异显著(P<0.05)。大写字母表示灌木,小写字母表示草本Fig. 3 Pielou evenness index and Alatato evenness index of understory shrubs and herbs. Different letter indicates significant difference at P < 0.05;capital letter represents shrub; lowercase letter represents herb.
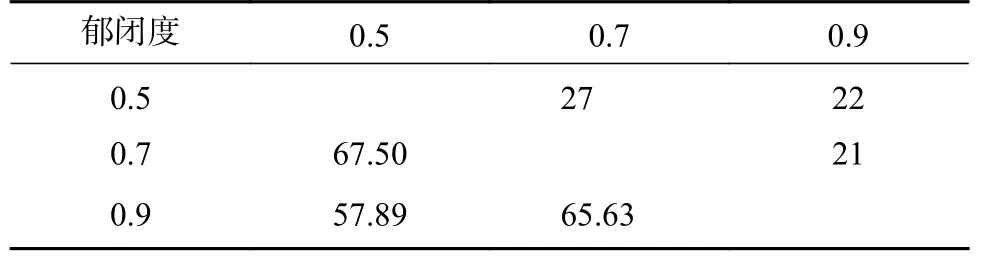
表 6 林下植物共有物种数和群落相似系数(SO/%)Tab. 6 Common species number and community similarity coefficient (SO/%) of understory vegetation
植物多样性指数反映群落结构、群落稳定程度和生境差异[16]。本研究灌木和草本植物的丰富度指数、Simpson 指数、Shannon-Wiener 指数差异显著。小郁闭度异质性更大,环境条件的改善和土壤养分的加快分解,有利于土壤种子库种子的萌发,增加植物种类及数量,提高植物多样性。因此,0.5 郁闭度林分林下植物多样性最高,并且显著高于0.9 郁闭度。此研究结果与张柳桦等[17]、赵燕波等[8]的研究一致,但曹子林等[12]对昆明地区华山松的研究指出,中郁闭度(0.7~0.8)林分灌木、草本植物的多样性指数高于弱郁闭度林分(0.5~0.6),其认为弱郁闭度林分植物均匀度小而导致植物多样性降低。本研究草本植物的丰富度指数和多样性指数均高于灌木植物。草本植物生长周期短,其更强的环境适应能力,更大地影响物种多样性[18]。灌木层和草本层的物种均匀度指数在各个郁闭度间均无明显差异,说明郁闭度的变化未造成华山松林下生态因子在空间分配上的明显差异[17]。也有研究指出,均匀度指数的变化可能受到种间种内竞争、环境因子、种群密度等因素的综合影响[12,19,20]。
林分共有种和相似系数是物种多样性差异的重要表现特征[21]。本研究0.5、0.7 郁闭度共有种最多,但0.9 郁闭度与其他郁闭度共有种仅差1 种,说明郁闭度降低增加物种数的同时,灌木植物的逐渐密闭为草本植物提供与高郁闭度相似的生长环境[8,22]。0.7 郁闭度与其他郁闭度林下物种Sorensen 指数相近且较高,林下植被群落结构较为稳定。
研究表明,通过适当的郁闭度调控,林下灌木和草本植物的更新加快,华山松人工林生态系统的稳定性有所提高。随郁闭度的降低,灌木和草本植物的种类、数量和生物多样性明显增加,0.5 郁闭度林分最大,各郁闭度植物在空间分布上无明显差异,0.7 郁闭度林分林下植物群落较为稳定。不同郁闭度植物多样性的变化还与土壤养分、物种竞争等因素密切相关,还有待进一步研究。
猜你喜欢
中华建设(2024年3期)2024-03-16 08:04:52
中华建设(2023年10期)2023-10-07 05:23:04
科海故事博览(2022年28期)2022-11-25 03:09:48
少儿美术(2020年8期)2020-11-06 05:21:38
福建林业(2020年5期)2020-03-18 08:23:02
中国森林病虫(2020年1期)2020-01-18 07:21:52
热带林业(2019年4期)2019-03-05 09:53:58
新闻传播(2018年15期)2018-09-18 03:20:12
森林工程(2018年3期)2018-06-26 03:40:46
农民致富之友(2014年7期)2014-10-21 20:06:25
