
宝天曼国家级自然保护区锐齿槲栎林短期动态变化
2020-12-29 08:11:22孙怡洁李镇江路阳任世坦肖曼于晨一潘名好王婷
河南农业大学学报 2020年6期
孙怡洁, 李镇江, 路阳, 任世坦, 肖曼, 于晨一, 潘名好, 王婷
(河南农业大学林学院,河南 郑州 450002)
森林动态变化是指森林生长和衰亡的变化,由于森林动态周期较长,更多的学者开始强调对森林群落进行长期重复研究的重要性。建立长期监测型固定样地是研究物种分布格局,群落动态和生物多样性维持机制等森林生态系统特征,过程与机制的重要平台[1]。自1980年在巴拿马中部Barro Colorado Island(BCI)建立第一个永久样方,全球形成了以热带雨林为中心的长期生态研究网络,在自然生态系统中建立了数十个大型动态监测点,对群落动态等理论研究也取得了长足进展。如日本科学家UMEKI[2]分析了日本北海道天然林固定样地的群落动态,得出了影响该样地物种死亡的非生物因素的结论,GASTAUER等[3]对巴西Cerrado地区2个廊道森林的物种组成、群落和种群动态进行研究,相关研究成果主要集中在森林种群生态学和空间分布格局等方面。中国也有大量学者对森林固定样地进行研究,葛结林等[4]和金毅等[5]分别对湖北神农架9.6 hm2样地、古田山24 hm2样地进行动态监测,通过对物种组成、径级分布、群落特征等方面进行研究,发现这两个样地群落动态特征变化不大;李效雄等[6]和许涵等[7]分别对祁连山自然保护区青海云杉林内10.2 hm2和海南尖峰岭60 hm2的动态监测样地进行物种调查,分析了样地内物种组成、优势植物和群落结构。宝天曼自然保护区是中国北亚热带和暖温带的气候划分线[8]。多年来,宝天曼作为“世界生物圈保护区”,为生态研究提供了大量的参考数据。藏亚静[9]、王亚平等[10]对宝天曼森林群落空间格局进行了分析,结果表明,在完全空间随机零模型下,物种的空间分布为聚集分布,闫满玉[11]对木本植物单物种-面积关系进行研究,说明随机过程决定宝天曼落叶阔叶林群落空间多样性格局的形成。尤业明[12]研究阐明了地上植物和地下微生物在土壤C转化中的作用机制及其调控过程,白晓旭等[13]和赵爱花等[14]研究发现,宝天曼不同林龄和林分类型土壤分别具有特有的种属组成及高丰度和低丰度种属。林龄与林分类型都对土壤微生物群落结构影响显著,且林分类型的影响大于林龄。许传阳等[15]研究发现,物种多样性变化的主成分不明确,土壤有机质与物种多样性相关性最显著。齐光等[16]对宝天曼的森林植被碳库进行分析,明确森林类型和群落结构对研究区森林植被碳库及其分配格局的影响。牛晓栋等[17]对宝天曼锐齿栎林净生态系统碳交换特征作进一步研究。樊鹏振等[18]对宝天曼保护区树附生苔藓植物物种分布进行研究,发现树附生苔藓植物多附生在胸径大于 15 cm的树干上且与附生树种相关性总体不明显。现有的研究多数集中于群落格局分析、土壤与微生物的关系、物种多样性、物种与面积和森林碳库及碳循环的研究,对宝天曼自然保护区优势种群的动态分析不足,韦博良等[19]对宝天曼锐齿槲栎林群落进行分析,认为锐齿槲栎群落是宝天曼地区的重要植被类型之一,研究其动态变化对分析宝天曼自然保护区群落特征具有重要意义。在此基础上,本研究以宝天曼1 hm2锐齿槲栎林为研究对象,通过分析2009年和2014年2次调查各物种的数量和胸径变化,比较不同物种胸高断面积变化,探讨优势物种死亡率、补员率和胸径的关系,预测优势种群的变化趋势,为宝天曼自然保护区森林动态观测提供基础资料,以期更好地把握群落动态规律,为宝天曼自然保护区的可持续发展提供一定的科学依据。
1 研究方法
1.1 研究地概况
宝天曼国家级自然保护区是河南省建立的第一个自然保护区,被誉为“世界生物圈保护区”和“世界地质公园”,是中原地带唯一保存最为完整的过渡综合性森林生态系统,位于河南省西南部、秦岭东段伏牛山南坡的内乡县北部山区(33°35′43″~33°20′12″N, 111°46′55″~112°03′32″E),第二级地貌阶梯向第三级地貌阶梯过渡的边缘,处于中国暖温带与北亚热带的气候过渡带,具有明显东部季风区的过渡性特点,四季分明,年平均气温为15.1 ℃,年平均降水量为885.6 mm[20]。地带性植被为落叶阔叶林、温性针叶林针阔混交林。保护区内土壤可划分为山地棕壤、山地黄棕壤和山地褐土3种,主要基岩是花岗岩、石灰岩和砂岩。
1.2 研究方法
1.2.1 样地设置 在宝天曼选取1 hm2(100 m×100 m)的锐齿槲栎林样地,2009年首次群落调查时对样地内所有胸径(DBH)≥1 cm的乔木和灌木进行编号并标记挂牌,记录其物种名称、胸径、坐标位置、树高和生活力(枯死或良好),并进行长期监测。2014年对样地进行复查,包括所有2009年所调查过的植物以及新增的植物,并对新增植物进行挂牌编号,记录物种名称、坐标位置、胸径、树高和生活力。
1.2.2 数据处理 为统计和比较方便,本次研究仅分析2次调查中DBH≥1.0 cm的个体,计算2次调查时物种的数量、死亡率、补员率、以及重要值,并通过计算各个物种的死亡率(M)和补员率(R)参数,比较各物种的生长情况。
M=(lnN0-lnSt)/T
(1)
R=(lnNt-lnSt)/T
(2)
重要值=(相对密度+相对频度+相对胸高断面积)/3
(3)
式中:T为2次调查时间间隔5 a;N0和Nt分别为某一种群在第1次和第2次调查时的存活个体数;St是该物种在2次调查时都存活的个体数。(3)式中,
相对密度=单个物种个体数/全部物种个体数×100%;
相对频度=单个物种出现的样方数/全部样方数×100%;
相对胸高断面积=单个物种平均胸高断面积/全部物种平均胸高断面积之和×100%。
参考文献[21],各物种的径级划分标准为,Ⅰ级(DBH<5 cm),Ⅱ级(5 cm≤DBH<10 cm),Ⅲ级(10 cm≤DBH<15 cm),Ⅳ级(15 cm≤DBH<20 cm),Ⅴ级(20 cm≤DBH<25 cm),Ⅵ级(DBH≥25 cm)。
采用Microsoft Excel 2016 和 R3.5.2(http://www.r-project.org)进行数据分析和绘图。
2 结果与分析
2.1 宝天曼锐齿槲栎林主要物种种群数量变化
该样地中,5 a间木本植物个体数由2 677株减少到2 364株,减幅为11.69%,其物种数由68种减至66种,死亡4种,新增2种。从表1 可以看出,2009年重要值前10名的物种是锐齿槲栎(Quercusalienavar.acuteserrata)、华山松(Pinusarmandii)、四照花(Dendrobenthamiajaponicavar.chinensis)、崖樱桃(Cerasusscopulorum)、三桠乌药(Linderaobtusiloba)、秦岭木姜子(Litseatsinlingensis)、山核桃(Caryacathayensis)、青榨槭(Acerdavidii)、朴树(Celtissinensis)和灯台树(Bothrocaryumcontroversum)。2014年重要值前10名的物种分别是锐齿槲栎、华山松、四照花、崖樱桃、山核桃、三桠乌药、秦岭木姜子、青榨槭、灯台树和朴树。在这5 a中,山核桃的重要值增加最大,为3.5%,华山松次之,为2.8%,重要值减少最多的是海州常山(Clerodendrumtrichotomum),减少了0.8%,朴树的重要值没有发生变化。
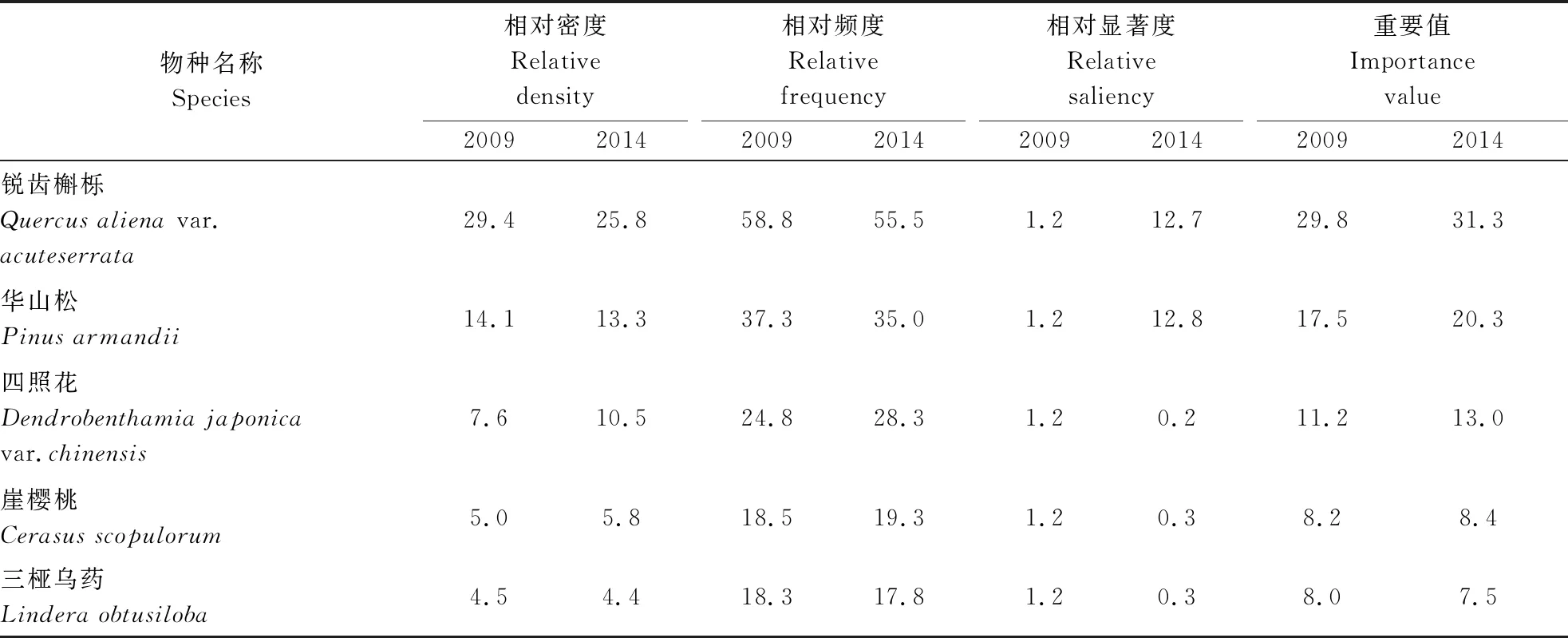
表1 宝天曼锐齿槲栎林主要物种重要值

续表1
样地内主要物种的死亡率和补员率见图1。样地内物种的年均补员率为2.64%,年均死亡率为4.56%。样地主要物种中仅有34.78%物种的补员率大于死亡率,其中水榆花楸的补员率最大,为8.93%,灯台树和海州常山次之,分别为5.88、5.75%,锐齿槲栎的补员率最低,仅有0.07%,华山松、山核桃、漆树没有补员。死亡率最大的是白桦,高达35.84%,除了水榆花楸没有死亡个体,四照花的死亡率最低,仅为0.60%。
2.2 宝天曼锐齿槲栎林主要物种种群平均胸径的变化
由于个体死亡、补员以及树木正常生长发育,使物种胸径的增长发生变化。在2009—2014年间,样地内主要物种的平均胸径变化见图2。由图2可知,样地主要物种2009年平均胸径为6.75 cm,2014年为7.54 cm,81.25%的物种平均胸径呈增加趋势。其中锐齿槲栎平均胸径增加量最大,为3.70 cm,漆树次之,增加了3.63 cm。锐齿槲栎、华山松、四照花、朴树等几个物种的平均胸径5 a间增加超过2 cm,另有16个物种的平均胸径增加不超过1 cm,卫矛和山梅花的平均胸径变化幅度不大。5 a间椴树和白桦的平均胸径减小,分别减少了0.46、2.72 cm。
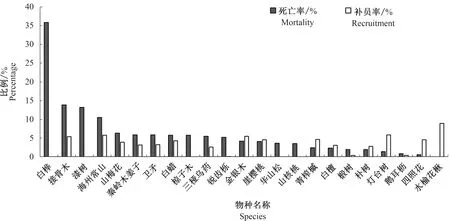
注:白桦Betula platyphylla, 接骨木Sambucus williamsii, 漆树Toxicodendron vernicifluu,海州常山Clerodendrum trichotomum, 山梅花Philadelphus incanus, 秦岭木姜子Litsea tsinlingensis, 卫矛Euonymus alatus, 白蜡Fraxinus chinensis, 椋子木Cornus macrophylla, 三桠乌药Lindera obtusiloba, 锐齿栎Quercus aliena var. acuteserrata, 金银木Lonicera maackii, 崖樱桃Cerasus scopulorum, 华山松Pinus armandii, 山核桃Carya cathayensis, 青榨槭Acer davidii, 白檀Symplocos paniculata, 椴树Tilia tuan, 朴树Celtis sinensis, 灯台树Bothrocaryum controversum, 鹅耳枥Carpinus turczaninowii, 四照花Dendrobenthamia japonica var. chinensis, 水榆花楸Sorbus alnifolia。下同。The same as below.
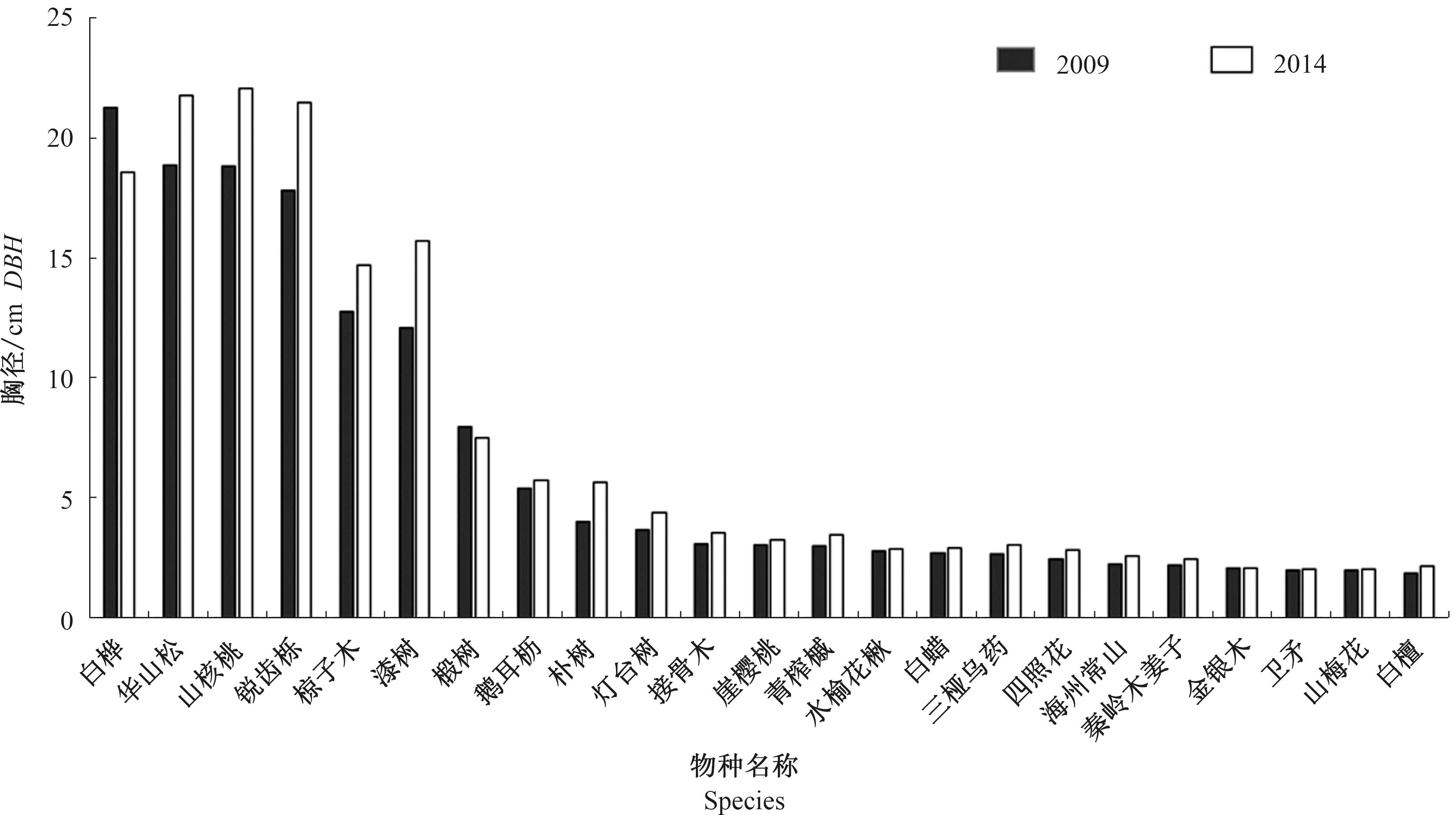
图2 宝天曼锐齿槲栎林主要物种(重要值前20)平均胸径变化
2.3 宝天曼锐齿槲栎林主要物种种群的个体径级分布
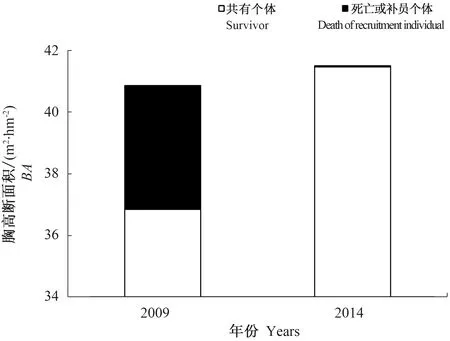
不同径级的死亡率和补员率见图3,由图3-A和3-B可知,第Ⅱ径级(5 cm 5 a间补员个体较少,都集中在Ⅰ级(DBH≤5 cm) ,此径级补员率达到100%。2次调查存活个体胸径在Ⅰ级(DBH≤5 cm)都是最多,分别占样地总个体数的47.29%和55.09%,2009年胸径在25 cm以上个体最少,占全部个体的8.04%,2014年第Ⅲ径级(10 cm 注:A为不同径级个体死亡率,B为不同径级个体补员率,C为2009年存活个体径级分布,D为2014年存活个体径级分布。 5 a间样地胸高断面积(Basal area,BA)变化如图4所示,由图4可以看出,2009年BA为40.87 m2·hm-2,2014年BA为41.51 m2·hm-2,样地BA净增0.64 m2·hm-2。分析数据发现,在这5 a期间,由于树木死亡而减少的BA为4.03 m2·hm-2,新增4.67 m2·hm-2,其中因为原有树木生长而增加的BA为4.62 m2·hm-2,占新增的大部分;因补员个体而增加的BA为0.05 m2·hm-2,占少部分。 5 a 间有48个物种的BA为净增加,其中锐齿槲栎的增量最大,为0.85 m2·hm-2,山核桃、四照花、金银木的BA增加量分别为0.07、0.07和0.04 m2·hm-2。有23个树种的BA为净减少,占总数的58.62%。其中,白桦BA减少最多,达0.21 m2·hm-2,其次为山杨,减少0.13 m2·hm-2,个体死亡数量较多可能是BA减少的主要原因。总体来看,5 a 间BA减少较多的树种其死亡数量较大,而新增BA较多的物种个体生长发育良好。 图4 宝天曼锐齿槲栎林补员个体、死亡个体和共有个体对2009和2014年胸高断面积的贡献 样地内物种的年均补员率为2.64%,年均死亡率为4.56%。重要物种补员数目差距较大,补员和死亡数量较多的物种,其个体数量和重要值在物种中排名前10位,说明物种的补员与死亡数与树种数量呈正相关关系。在这5 a发育过程中,水榆花楸补员率最大无死亡个体,华山松、山核桃和漆树没有补员个体,说明群落中水榆花楸物种正在逐渐取代其他3个物种的位置。不同物种的死亡率和补员率相差较大,锐齿槲栎、华山松等物种死亡率大于补员率,种群呈负增长;四照花、灯台树等物种补员率大于死亡率,种群呈正增长。 宝天曼自然保护区锐齿槲栎林1 hm2样地中植物个体平均死亡112.8株·hm-2·a-1,高于古田山5 hm2样地植物个体平均死亡株数[22](55.4株·hm-2·a-1),低于鼎湖山1 hm2季风常绿阔叶林木本植物在1999—2004年期间的平均死亡株数[23](261.4株·hm-2·a-1)。原因在于古田山的物种大部分为自然死亡,鼎湖山常绿阔叶林曾受过一定程度的自然灾害和人为因素的影响,而宝天曼锐齿槲栎林处在暖温带-北亚热带气候过渡带,很大部分原因可能是受气候变化的影响,自然死亡的个体较多。 群落的死亡率和补员率是反映群落更新动态的直接指标[24]。不同地点样地的补员率和死亡率不同,马来西亚Pasoh样地补员率和死亡率分别为1.65%和1.46%[25],样地内物种生长情况稳定;中国长白山阔叶红松林5 a间补员率为 5.5%,死亡率为10.9%[26],群落为负增长,有衰退趋势;古田山5 hm2样地补员率和死亡率则分别为6.65%和2.02%[5],群落为正增长,样地物种生长旺盛。由此可见,宝天曼锐齿槲栎林样地补员个体少,死亡个体多,总补员率处在较低水平,死亡率相对较高。 宝天曼锐齿槲栎林样地内主要物种2009年平均胸径为6.75 cm,2014年为7.54 cm,高于湖南八大公山样地(平均胸径5.41 cm)[27]。5 a间,大部分物种的胸径都有所增加,增幅较大的主要集中在乔木树种,其中锐齿槲栎、华山松、山核桃和漆树的平均胸径增加值超过2 cm,其死亡个体多集中在小径级的物种;卫矛和山梅花平均胸径变化幅度不大,是因为死亡个体数虽多于补员个体数,但死亡和补员的都是小胸径个体,所以对平均胸径变化没有什么影响。椴树和山杨的平均胸径减小,是因为胸径较大个体有死亡,而补员个体胸径较小,因此平均胸径减小。 不同径级的树木,其个体死亡情况有较大差异。在宝天曼锐齿槲栎林样地中,大多数的死亡树木其径级都在5~15 cm小径级之间,与干旱区雪岭云杉林群落物种死亡个体主要发生在小径级个体上[27]相似。树木死亡随径级增加而减小,大致呈倒“J”型分布,这与长白山典型真阔混交林中主要物种的倒“J”型分布[28]相似。这正好体现了森林更新的一种机制, 即如果树木死亡主要是由竞争所驱动, 则小径级树木相对于大径级树木死亡概率大[29],大量小径级个体的死亡表明,与其他径级相比小径级个体存在更为激烈的竞争[30]。因为小径级多为幼树或林下层的灌木树种,由于资源有限,小径级个体在发生光照、营养、空间等资源竞争时,明显处于弱势地位,同时,当受到病虫害侵袭时,小径级树木抵抗力稳定性更差,因而死亡率也更高[31]。虽然补员的树木也均为小径级,但是由于密度制约的原因,使得小径级个体在森林中的竞争更加激烈,死亡率也就相对较高。 5 a间样地内并没有发生大的外界因素干扰,大径级个体的死亡多数是由于风倒和被压所致, 因为大径级个体生长情况稳定, 在种内种间竞争中均处于优势地位,受自然因素影响很小,并且个体数量所占比例很小,因此死亡率相对较低[32]。 胸高断面积的变化除了树木自身增长引起,还与物种个体的死亡和补员有关。树木死亡对胸高断面积的变化影响较大,会减小胸高断面积的变化,而补员会使胸高断面积略有增加。5 a间样地种群的胸高断面积涨幅缓慢,这是由于虽然物种个体死亡较多,但是大径级个体在BA上占有绝对优势,相对于5 a间树木生长增加的BA(4.62 m2·hm-2)而言, 由死亡造成的BA减少量相4.03 m2·hm-2, 总的来看,BA减少较多的树种其死亡数量较大,而新增BA较多的物种个体生长发育良好,所以BA是增加的。 物种补员与死亡数目的差值, 反映出该种群大小的变化,而补员率与死亡率的差值, 则表征种群的变化率[23]。5 a间该样地主要物种四照花、崖樱桃、灯台树、朴树等的补员数量大于死亡数量,种群增长,其余种群有所减小,其中,漆树的降幅最大,为-13.18%。对比干扰较小的Pasoh样地只有2%的物种变化率超过5%。古田山样地内43个重要种群中有10个种群变化率超过5%,比例达23.26%。在宝天曼中有11.11%的重要种群的变化率超过5%,说明宝天曼样地种群的更新速度明显较快,这可能与宝天曼样地复杂的地形环境和较高的生物异质性有关。此外,种群变化率较高的几个物种(漆树、水榆花楸、接骨木、白桦树等)属于快速变动的种群,这些物种在未来十几年内数量会加倍或减半,那么未来十几年内宝天曼样地的群落组成与结构会发生很大变化[4]。 综上所述,宝天曼自然保护区锐齿槲栎林样地5 a间补员和死亡在物种间分布不均匀,随物种不同而变化;不同物种的死亡率和补员率相差较大;不同径级个体死亡分布有明显差异,小径级的物种死亡率较高,但是由于其大量补员个体,其平均胸径增加变化不大;大径级个体胸径增加量明显大于小径级个体,虽然有部分的物种死亡,但从整体来看,群落总体平均胸径处于增长的状态。宝天曼自然保护区锐齿槲栎林的群落结构在这5 a间发生了巨大变化,可能与气候变化、种间竞争、演替进程等因素有关,但对锐齿槲栎林如何响应环境变化,演替过程与气候变化间的联系,以及群落内部竞争机制缺乏研究,未来需要对这些方面进行深入研究,加强暖温带-亚热带过渡地区森林的监测和管理。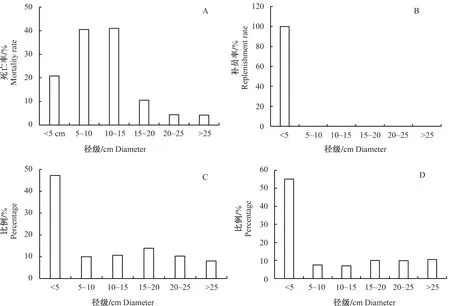
2.4 宝天曼锐齿槲栎林主要物种种群的胸高断面积变化
3 结论与讨论
3.1 宝天曼锐齿槲栎林种群数量变化
3.2 宝天曼锐齿槲栎林种群平均胸径变化
3.3 宝天曼锐齿槲栎林种群个体及补员死亡个体径级分布情况
3.4 宝天曼锐齿槲栎林种群胸高断面积与种群变化
猜你喜欢
吉林林业科技(2023年6期)2023-11-20 02:13:16广西林业科学(2022年4期)2022-11-07 01:50:34林业调查规划(2022年3期)2022-06-13 09:31:04广西林业科学(2021年6期)2022-01-21 07:39:10内蒙古林业调查设计(2021年5期)2022-01-05 02:50:40现代园艺(2021年23期)2021-12-01 07:47:44林业勘查设计(2020年1期)2021-01-18 02:40:48新农业(2020年18期)2021-01-07 02:17:08林业勘察设计(2017年4期)2017-07-06 08:46:12西北植物学报(2015年9期)2015-07-05 07:03:42
