CpG ODN协同小鼠肝癌细胞裂解物对小鼠原位移植性肝癌的抑制作用研究①
2020-12-26 06:16李欢欢王卫芳周晓晶
中国免疫学杂志 2020年23期
孙 鹏 李欢欢 王卫芳 窦 瑶 周晓晶
(长春中医药大学临床医学院,长春 130117)
肝癌是最常见的肝脏原发恶性肿瘤,其致死率极高[1]。寻找抗肝癌免疫中有效靶抗原是抗肝癌免疫的难题[2]。在肿瘤细胞裂解物(tumor cell lysate,TCL)中含有丰富的肿瘤相关抗原(tumor associated antibody,TAA),因此肿瘤细胞内所有抗原都可能是免疫攻击靶点,从而引起作用更强的抗肝癌免疫反应[3,4]。但肿瘤抗原的免疫原性弱,研究者们尝试了多种方法来提高TCL的免疫原性。常用的方法包括:用DC装载 TCL、以HSP为佐剂等[5,6]。Dong等[7]研究发现,结核分枝杆菌的HSP65能协助lewis肺癌的细胞裂解物诱导抗肺癌免疫反应。此外,人们也研究了含有胞苷酸-鸟苷酸的寡聚脱氧核苷酸(CpG ODN)与肝癌细胞裂解物联合应用的效果,CpG ODN是一种Th1型免疫增强剂,刺激机体产生Th1型细胞因子IFN-γ、IL-12等,并且能够辅助TAA产生CTL反应从而清除肿瘤细胞[8]。Dong等[9]研究发现,CpG1826能增强TCL的免疫原性,从而应用于人类HPV病毒永生化的肿瘤模型中。因此CpG ODN或许会辅助TCL诱导抗肝癌免疫。根据以上推测,我们将小鼠肝癌细胞H22的TCL与具有免疫刺激增强作用的C型CpG ODN联合应用于小鼠原位移植性肝癌模型,研究其抑瘤效果及机制。
1 材料与方法
1.1材料 H22细胞株由本实验室液氮冷冻储存,为悬浮生长,使用含10%FBS IMDM培养基,培养于5%CO2、37℃培养箱中。纯化的单链寡聚脱氧核苷酸购自日本TaKaRa大连分公司(OPC级,纯度90%,HPLC级,纯度99%)。IFN-γ、IL-17检测试剂盒购自R&D System公司。本研究使用的CpG ODN序列如下:C274(5′-TCGTCGAACGTTCGAGATGAT-3′),被溶于无菌PBS中,紫外分光光度计定量,检测内毒素<0.03 EU/mg,符合动物实验要求,于-20℃ 冷藏备用。雌性BALB/c 小鼠,6~8 周龄,18~22 g,购自吉林大学白求恩医学院动物实验中心。
1.2方法
1.2.1TCL的制备和鉴定 将生长状态良好的H22细胞用PBS洗涤、计数后取适量细胞制成细胞悬浮液。将细胞悬液于37℃水浴和-70℃冰箱中反复冻融5次,期间震荡混合20 s。用台盼蓝染细胞并于显微镜下观察细胞状态,保证没有活细胞存在。将制备合格的 TCL 放于-70℃冰箱中冻存备用。
1.2.2小鼠原位移植性肝癌模型的建立 将 1×107个H22细胞注射于BALB/c小鼠腹腔,1周后形成腹水。取腹水分离H22细胞并计数,以2×106个细胞注射于小鼠右后腿外侧皮下。待肿瘤直径 1 cm 左右分离肿瘤组织于冰冷无菌PBS中,显微镜下将瘤块剪碎至直径(1.2±0.2)cm,将瘤块种植于小鼠肝脏。
1.2.3原位移植性肝癌接种及免疫 BALB/c小鼠随机分为4组,每组10只。在-1 d每只小鼠肝内接种肝癌组织,然后于0、7、14、21 d经双侧腹股沟淋巴结引流区皮下分别注射PBS、TCL(2×106个/只)、CpG ODN(12.5 μg/只)和CpG ODN(12.5 μg/只)+TCL (2×106个/只),监测小鼠生存期。
1.2.4CTL杀伤实验 小鼠分组及免疫程序同1.2.3,于末次免疫后7 d,分离小鼠脾细胞并用TCL刺激培养。培养24 h时,加入小鼠IL-2(20 U/ml)继续培养24 h。然后离心并计数。刺激后的脾细胞作为效应细胞, H22细胞作为靶细胞进行杀伤实验。5×105个脾细胞稀释后与靶细胞共同培养,效靶比分别为12.5∶1、25∶1及50∶1。4 h后收集细胞并用台盼蓝染色,于显微镜下观察存活的细胞,体积较大的细胞为H22细胞并计数,每毫升中的细胞数=四象限中的细胞总数/4×稀释倍数×104,4个象限的细胞总数必须介于200~500个之间,而且细胞浓度要超过104个/ml。按以下公式确定CTL杀伤情况,CTL细胞毒性%=(靶细胞对照孔细胞数-实验孔细胞数)/靶细胞对照孔细胞数×100%。
1.2.5Th1型细胞因子的检测 小鼠分组及免疫程序同1.2.3,于末次免疫后7 d,分离小鼠脾细胞并与TCL共同孵育24 h,收集刺激上清并用IFN-γ和IL-12检测试剂盒检测IFN-γ和IL-12的水平。
1.3统计学分析 所有结果应用 SPSS17.0 软件进行分析,生存曲线用Kaplan-Meier方法绘制;肿瘤发生率用Fish检验进行比较。组间比较采用独立样本t检验,以P<0.05 为差异有统计学意义。
2 结果
2.1CpG ODN与TCL联合应用对小鼠原位移植性肝癌的抑制作用 BALB/c小鼠肝内接种瘤块后,给其注射PBS、TCL、CpG ODN、CpG ODN+TCL,观察小鼠生存期。结果如图1B所示,与其他组相比CpG ODN+TCL组的小鼠生存期明显被延长了(P<0.05),到77 d各组小鼠的存活率分别是PBS组20%、TCL组20%、CpG ODN组10%、CpG ODN+TCL组80%。每只小鼠死亡后分离其肿瘤组织如图1A所示,CpG ODN+TCL组小鼠10只有2只成瘤并且分别于接瘤后48 d和53 d死亡,其他8只小鼠均未成瘤,这8只小鼠直到77 d全部成活且状态良好。而其他各组成瘤率均在80%以上,且相继死亡。

图1 CpG ODN+TCL对小鼠原位移植性肝癌的抑制作用Fig.1 Inhibitory effect of CpG ODN+ TCL against orthotopic-transplantated liver cancer in miceNote:A.Tumors in liver from dead mice.PBS.dead/total=8/10,TCL.dead/total=8/10,CpG ODN.dead/total=9/10,CpG ODN+TCL.dead/total=2/10;B.Survival of mice with orthotopic-transplantated liver cancer.*.vs PBS;#.vs CpG ODN;&.vs TCL.
2.2CpG ODN协同TCL诱导小鼠脾细胞增殖反应 BALB/c小鼠被随机分为4组,末次免疫后7 d,取小鼠脾,CpG ODN组及CpG ODN+TCL组小鼠的脾较其他两组明显增大,见图2A。分离脾细胞并计数,计数结果如图2B所示,CpG ODN组及CpG ODN+TCL组小鼠脾细胞数分别是1.8×108个和1.78×108个,远远高于PBS组、TCL组的脾细胞数(P<0.05),但二者之间差异无统计学意义(P>0.05)。
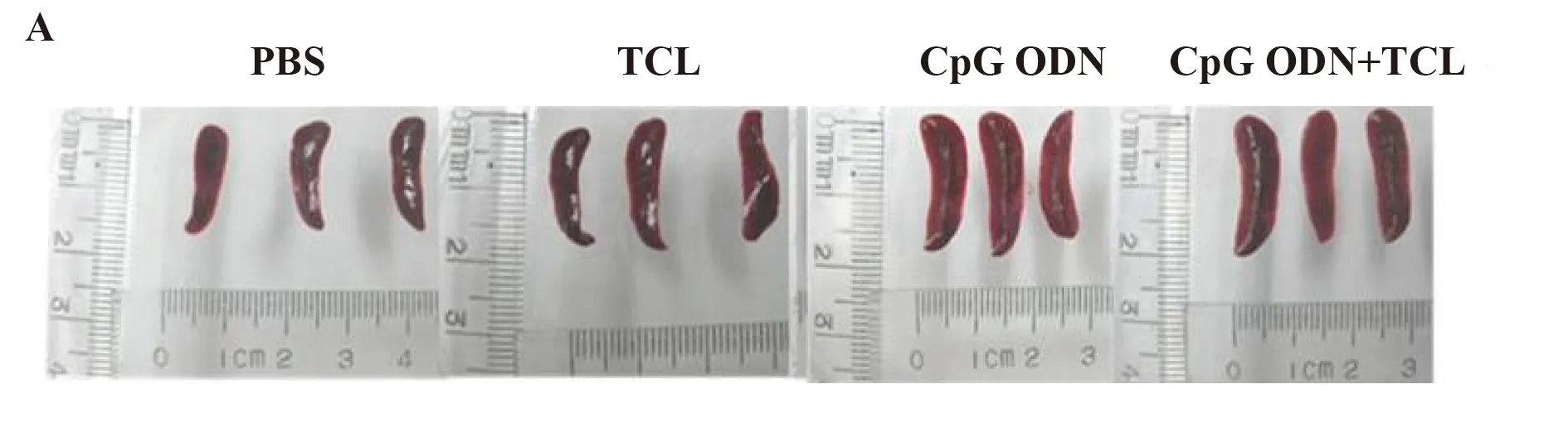
图2 CpG ODN协同TCL对小鼠脾的影响Fig.2 TCL adjuvanted CpG ODN induced response of spleen in miceNote:A.Morphology of spleens from three mice after 7 d of the last injection.Sizes of the spleens were compared based on the scale;B.Lymphocyte numbers in spleens from three mice.Each bar represents of lymphocyte number from spleens(down).*.P<0.05 vs PBS group;#.P<0.05 vs TCL group.
2.3CpG ODN协同TCL诱导特异性CTL反应 杀伤结果如图3所示,当效靶比为分别为25∶1和50∶1 时,来源于CpG ODN+TCL免疫鼠的脾细胞能裂解H22细胞,裂解率分别为48%和51%,而其他各组均无裂解效应(P<0.05)。

图3 CpG ODN协同TCL诱导了CTL反应Fig.3 CTL respense induced by CPG PDN+TCLNote:*.vs PBS;#.vs CpG ODN;&.vs TCL.
2.4CpG ODN协助TCL诱导小鼠分泌Th1型细胞因子 结果如图4所示,CpG ODN+TCL组与CpGODN组产生的IFN-γ 和 IL-12水平均较高,均为PBS组及TCL组的2倍左右(P<0.005),且两组间无差异(P>0.05)。
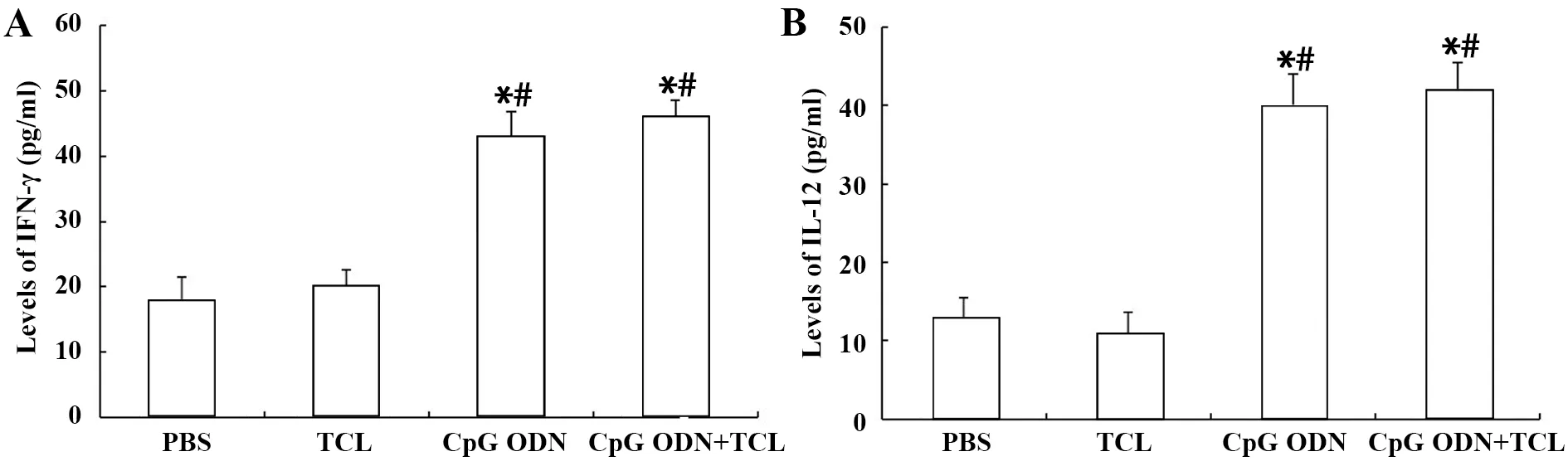
图4 CpG ODN协助TCL诱导小鼠分泌Th1 型细胞因子Fig.4 Th1 biased cytokines induced by CPG ODN+TCL in miceNote:A.Levels of IFN-γ;B.Levels of IL-12.*.P<0.05 vs PBS group;#.P<0.05,vs TCL group.
3 讨论
在TCL制备过程中本研究组发现其含有分子量不同的多种蛋白质,这些蛋白质中一定有丰富的肿瘤抗原存在肿瘤抗原的免疫原性弱,不足以诱导产生特异性的肿瘤杀伤反应。而C型CpG ODN具有强大的免疫刺激作用,作为Th1型免疫佐剂可增强抗原的免疫原性,研究组也证实了其能够协同TCL有效的抑制原位移植性肝癌的发生和延长小鼠的生存期。
本研究组推测CpG ODN与TCL联用对小鼠原位移植性肝癌的抑制作用与CpG ODN能协助TCL诱发特异性细胞免疫反应进而杀伤肿瘤细胞有关。CpG ODN能通过以下途径协同TCL产生特异性细胞免疫反应清除肝癌细胞。第一,本研究组另一个研究发现CpG ODN能上调MHC Ⅰ分子的表达,从而促进TCL递呈及交叉递呈[10]。第二,CpG ODN通过刺激B细胞和pDC产生IL-12 和 IFN-γ,为TCL提供Th1免疫环境,促进CD4+T细胞分化为Th1 CD4+T 细胞,使CTL浸润到肿瘤位点特异性清除肿瘤细胞。而且IL-12还能诱导T细胞和NK细胞分泌IFN-γ,它能提高肝癌细胞的MHC分子和共刺激分子的表达,从而使肝癌细胞更容易被CTL识别和清除。本研究中,发现单独使用CpG ODN或将其与TCL联用明显促进了小鼠脾细胞增殖、促进了IFN-γ 和 IL-12的分泌,且两者之间无差别,说明了CpG ODN有强大的免疫增强作用和促进Th1型细胞免疫反应作用。在进一步的CTL杀伤实验中,CpG ODN能协助TCL杀伤肝癌细胞,说明CpG ODN促进的Th1型细胞免疫反应增强了TCL特异性肿瘤细胞杀伤反应。而单独应用CpG ODN却没有这样的作用,说明CpG ODN虽能具有免疫刺激作用,但在没有肿瘤抗原存在的情况下CpG ODN的免疫刺激引起的非特异性抗肿瘤效应不足以抑制肿瘤细胞的生长,这也与在抑制肿瘤实验中CpG ODN与TCL联合有抑瘤效果而单独使用没有效果相统一。说明CpG ODN与TCL联用的抑瘤效果是通过诱导了特异性细胞杀伤反应清除肿瘤细胞实现的,具体机制还需进一步研究。
猜你喜欢
现代仪器与医疗(2022年4期)2022-10-08
实用手外科杂志(2022年2期)2022-08-31
健康体检与管理(2022年4期)2022-05-13
陶瓷学报(2021年5期)2021-11-22
现代临床医学(2021年5期)2021-11-02
昆明医科大学学报(2021年1期)2021-02-07
中国生殖健康(2020年2期)2021-01-18
中国兽医杂志(2019年5期)2019-09-18
生物学教学(2018年10期)2018-11-29
太空探索(2016年6期)2016-07-10