
罗氏沼虾第二步足的性二型研究
2020-12-25 02:31杨明丁福江
中国渔业质量与标准 2020年6期
杨明,丁福江
(1. 上海市水产研究所; 2. 上海市水产技术推广站:上海 200433; 3. 上海申漕特种水产开发公司,上海,201516)
性二型(sexual dimorphism),即两性异形,是指种内雌、雄个体之间在诸如形态大小、身体结构、体色或身体局部形态等特征上存在的显著差异,在生物界中普遍存在[1-2]。甲壳动物的性二型现象也较为常见,如近方蟹(Hemigrapsustakanoi),雄性头胸甲宽显著大于雌性[3]。此外,对于沼虾属(Macrobrachium)种类也有不少形态方面的性二型报道,如红足沼虾(Macrobrachiumdayanum)雄性的头部、头胸甲、额角、第一和第二步足均大于雌性[4],海南沼虾(Macrobrachiumhainanense)性成熟雄虾个体大于雌虾[5]。对于两性身体局部特征的研究也较为常见,其中对甲壳动物的战斗武器——螯足的性二型研究引起了进化生物学家的极大兴趣[6-10]。许多甲壳纲中的十足目动物拥有一对(等螯型,isochelous)或一只(异螯型,heterochelous)强大的螯足,这些强大的螯足作为甲壳动物的战斗武器,在社群等级序列的确定、夺取雌性的交配权、繁殖行为和领域占有等方面发挥着关键的作用[11-14]。甲壳动物在生长发育过程中,身体某个器官相对于整体生长常表现出不同的生长速率,因此在甲壳动物的性二型研究中,异速生长常被用于比较雌、雄两性间形态的差异程度[15-17]。此外,对于甲壳动物螯足的异速生长,生物学家也开展了相关研究。如Gu等[18]研究了红螯螯虾(Cheraxquadricarinatus)步足的异速生长;Kobayashi[19]对日本绒螯蟹(Eriocheirjaponica)步足的异速生长式型也进行了相关报道;Mossolin等[20]对欧式沼虾(Macrobrachiumolfersi)第二步足的异速生长进行了相关探讨。
罗氏沼虾(Macrobrachiumrosenbergii)原产于印度洋-西太平洋的热带、亚热带水域,隶属于甲壳纲(Crustacea)、十足目(Decapoda)、腹胚亚目(Pleocyemata)、长臂虾科(Palaemonidae)、沼虾属(Macrobrachium),目前是中国重要的淡水虾类养殖品种。性成熟的罗氏沼虾种群结构较为复杂,其中性成熟的雄性罗氏沼虾个体显著大于雌虾。雄性依照第二步足的特征依次分为蓝螯型(blue-clawed males, BC)、橙螯型(orange-clawed males, OC)和小型雄虾(small males, SM)3种形态型[21-22]。其中BC雄虾第二步足各部分均呈现深蓝色,其长度大于身体,且其上有棘,棘长且坚硬,呈现深蓝色;OC雄虾第二步足长度与身体长度相当,有时也小于身体长度,掌节和指节常呈现橙色,其上同样有棘,但棘与BC相比小且易碎,呈现橙色,但棘的尖端末处呈现黑色;SM雄虾第二步足长度小于身体长度,步足颜色变化较大,有时很难仅仅根据颜色将SM与OC进行有效区分,其指节上的固定指有时为蓝色,腕节末端有红带,掌节大多时候呈现白色或淡蓝色,其棘与OC类似。尽管已有学者注意到罗氏沼虾的第二步足在繁殖行为、隐蔽物的占取、交配权的竞争等方面发挥着重要作用[23-25],但对其第二步足的性二型基础研究尚欠深入。鉴于此,本研究基于性成熟罗氏沼虾第二步足相关性状参数的测量资料,对罗氏沼虾第二步足的性二型现象进行了研究,以期为深入探讨罗氏沼虾性二型的进化提供基础资料。
1 材料与方法
1.1实验材料和性状测量方法
取池塘养殖185日龄达性成熟的罗氏沼虾70尾:其中,雌虾30尾,体长(90.84±8.47) mm、体重(20.35±5.89)g;雄虾40尾,体长(100.44±9.11)mm、体重(29.88±8.59)g。分别对雌、雄罗氏沼虾第二步足重(second cheliped weight,SCW)、第二步足长(second cheliped length,SCL)、头胸甲长(carapace length,CL)以及第二步足之座节长(ischium length,IL)、长节长(merus length,ML)、腕节长(carpus length,CP)、掌节长(propodus length,PL)、指节长(dactylus length,DL)、掌节宽(propodus width,PW)、掌节高(propodus height,PH)等10个性状参数进行测定,用以比较分析罗氏沼虾雌、雄虾第二步足的性二型。鉴于在采样过程中SM数量过少,且其体型有时小于雌虾,因此雄虾仅选择BC和OC。长度测量采用数显游标卡尺(精度0.01 mm),质量测定采用灵敏度为0.01 g的电子天平,称重前用纱布吸干样本表面水分。鉴于罗氏沼虾为等螯型沼虾,因此仅选取第二步足右足进行测量。各性状参数测量标准如下:
头胸甲长:眼柄基部到头胸甲中线的距离;第二步足长:平直状态下座节基部至指节前端的距离;第二步足重:虾体的第二步足右足质量;掌节宽:掌节的最大宽度;掌节高:掌节的最大高度;第二步足之座节、长节、腕节、掌节和指节长测量标准为自关节基部中线到另一端(或尖端)中线的距离(图1)。

图1 罗氏沼虾第二步足测量示意图Fig.1 Schematic illustration of the second cheliped of Macrobrachium rosenbergii
1.2 数据分析与处理
试验数据在做进一步统计分析前,分别检验其正态性(Kolmogorov-Smirnov test)和方差齐性(Levene test)。对于符合正态分布,但方差不一致的数据,则对数据进行适当转换。经检测,雌、雄虾所有性状参数均符合正态分布(P>0.05);对数据进行ln转换后符合方差齐性。采用独立样本t-检验比较雌、雄两性的第二步足长和第二步足重。对于座节长、长节长、腕节长、掌节长及指节长等特征变量,则以第二步足长为协变量,性别为因子,做单因素协方差分析(One-way ANCOVA),检测两性第二步足各变量特征的性别效应。在进行单因素协方差分析前,检验各组总体斜率的一致性,经检测,座节长、长节长、腕节长、掌节长和指节长适合协方差分析,而掌节宽(P=0.030)、掌节高(P=0.002)主体间效应(性别×第二步足长)显著,不适合使用协方差分析。利用线性回归进一步分析第二步足相关性状参数与头胸甲长的关系,检验有无明显的线性关系来分析雌、雄个体间第二步足相关变量的异速生长;采用Gibbons等[26]所报道的性二型指数(sexual dimorphism index,SDI)表示雌、雄罗氏沼虾个体间第二步足形态特征差异的不同程度,其计算公式如下:
采用幂函数方程W=aLb拟合第二步足长与第二步足重之间的关系,其中a代表第二步足生长的条件因子,b值可用来判定第二步足是否处于匀速生长。将W=aLb经过自然对数转换后化为线性方程lnW=lna+blnL,式中W代表第二步足重,L代表第二步足长。当b=3时,为等速生长;当b<3时,表示负异速生长模式,即第二步足长生长快于第二步足重生长;当b>3时,表示正异速生长模式,即第二步足重生长快于第二步足长生长。
异速生长模型以幂函数方程y=axb构建,其中x为头胸甲长,y为第二步足各性状之长度、宽度或高度,将y=axb进行以e为底的自然对数转换,化为线性方程lny=lna+blnx,a为y轴截距,b为异速生长指数。当b=1时,为等速生长,此时罗氏沼虾第二步足各部分性状参数的生长与头胸甲等比例增长;b>1时,为快速生长,此时第二步足各部分性状参数的生长比头胸甲生长快;b<1时为慢速生长,此时第二步足各部分性状参数的生长比头胸甲生长慢。
本研究中所有试验数据通过IBM SPSS Statistics 23统计软件进行统计分析,P<0.05为差异显著,P<0.01为差异极显著。描述统计值用平均值±标准差(Mean±SD)表示。采用Origin 2018进行绘图。
2 结果与分析
2.1 第二步足的性二型
表1为雌、雄罗氏沼虾第二步足相关变量形态特征值和性二型的比较结果。经Kolmogorov-Smirnov检验,罗氏沼虾两性第二步足重、第二步足长以及第二步足之座节长、长节长、腕节长、掌节长、指节长及掌节宽、掌节高等9项性状参数均符合正态分布(P>0.05)。其中第二步足长、第二步足重的正态分布见图2。所有性状参数经loge(自然对数)转化后方差齐性,符合参数统计要求。从表1可以看出,在所测定的70尾样品中,雌虾第二步足长均值为(76.12±10.11)mm,第二步足长度范围56.15~94.00 mm,变异系数为13.28%;雄虾第二步足长均值为(105.74±16.54) mm,第二步足长度范围71.90~138.11 mm,变异系数为15.64%;雌虾第二步足重均值为(0.86±0.37)g,第二步足重范围0.27~1.59 g,变异系数为43.02%;雄虾第二步足重均值为(2.49±1.15)g,第二步足重范围0.61~5.85 g,变异系数为46.18%;对两性第二步足长、第二步足重进行的独立t-检验分析结果显示:雄虾第二步足长和第二步足重均极显著大于雌虾(P<0.001)。协方差分析结果表明:座节长、长节长、腕节长、掌节长和指节长在两性间的差异均达到了极显著水平(♂>♀,P<0.01)。

图2 雌、雄罗氏沼虾的第二步足长、第二步足重正态分布图(A、B)Fig.2 Normal distribution of the second cheliped length and second cheliped weight of female and male Macrobrachium rosenbergii,respectively (A, B)
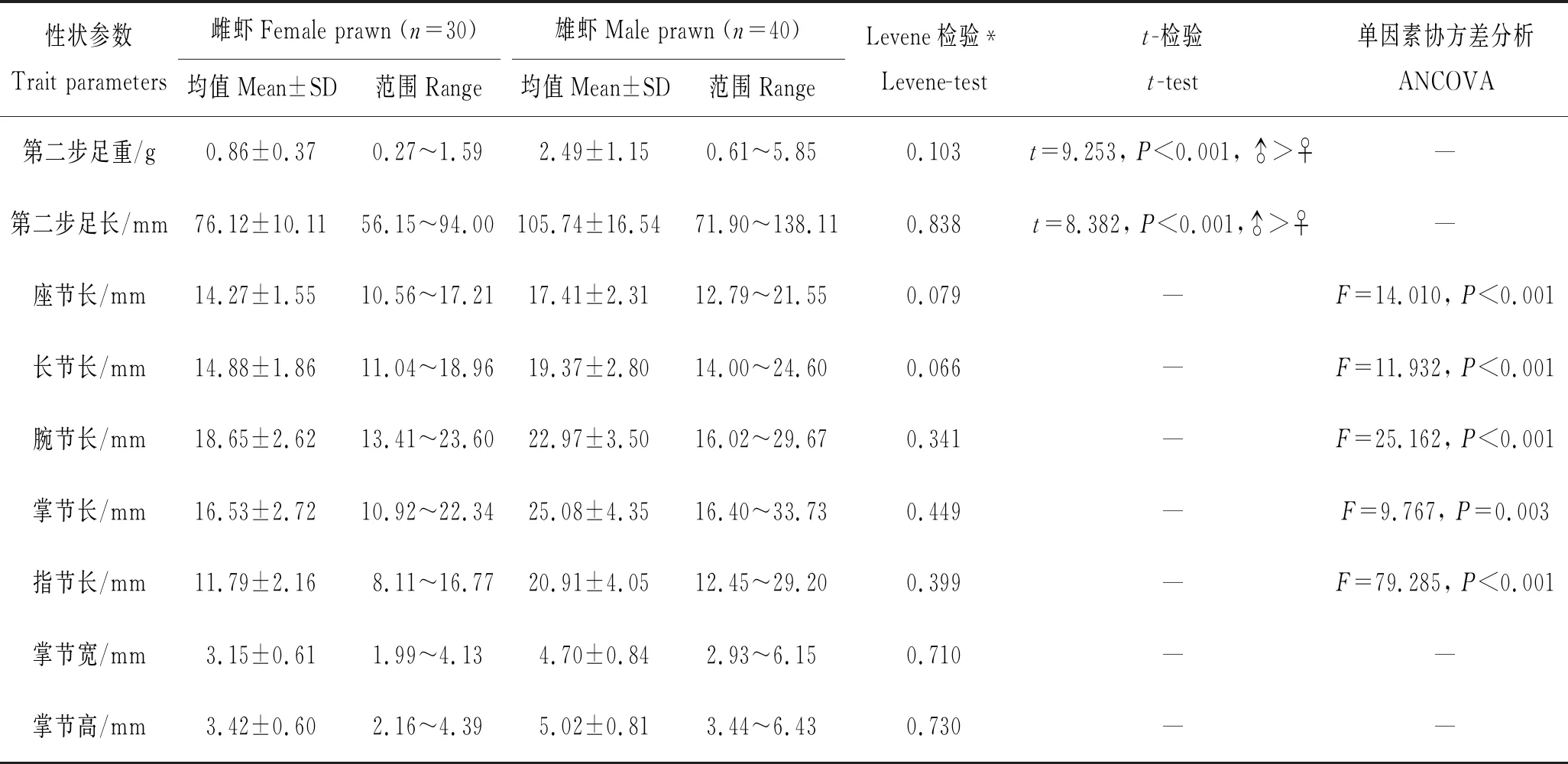
表1 罗氏沼虾两性第二步足变量统计值和性二型的比较结果
2.2 异速生长
对与性别有高度统计学意义的第二步足重、第二步足长以及第二步足之座节长、长节长、腕节长、掌节长和指节长等7项性状参数分別进行第二步足长-第二步足重、头胸甲长-座节长、头胸甲长-长节长、头胸甲长-腕节长、头胸甲长-掌节长及头胸甲长-指节长的一元线性回归分析(图3、图4)。图3为雌、雄虾第二步足长-第二步足重的自然对数回归关系。依图3可知,雌、雄虾第二步足长-第二步足重的自然对数回归呈显著的线性关系,决定系数均在0.91以上,说明第二步足长生长与第二步足重生长的相关程度较高。雌、雄虾b值均大于3,且雄虾的b值小于雌虾b值,表明雌、雄虾第二步足重的生长为正异速生长型,即第二步足质量生长快于第二步足长度生长,雄虾的第二步足-体重异速生长水平大于雌虾。
图4(A~H)分别为头胸甲长与第二步足长、座节长、长节长、腕节长、掌节长、指节长、掌节宽及掌节高的线性回归方程。从图4可以看出,罗氏沼虾两性成体的第二步足长、座节长、长节长、腕节长、掌节长、指节长与头胸甲长有明显的线性关系。雄性的第二步足长、座节长、长节长、腕节长、掌节长和指节长随头胸甲长的生长速率大于雌性,且雄性均呈现正异速生长(b>1);雌性第二步足长、座节长、长节长、腕节长随头胸甲长的生长速率小于雄性,呈现负异速生长(b<1);掌节长和指节长随头胸甲呈现正异速生长(b>1),且异速生长水平小于雄性。尽管掌节宽和掌节高在两性间均表现为正异速生长,但两者异速生长水平相当,即雌、雄掌节宽和掌节高生长速率一致。

图3 罗氏沼虾雌、雄虾第二步足长与第二步足重的回归关系(数据经过自然对数转换)Fig.3 Regression relationship between the second cheliped length and second cheliped weight in female and male Macrobrachium rosenbergii(data was processed with ln-transformation)

图4 罗氏沼虾第二步足长、座节长、长节长、腕节长、掌节长、指节长、掌节宽、掌节高与头胸甲长的线性回归(A-H)Fig.4 Linear regression of SCL, IL, ML, CP, PL, DL, PW, PH via CL of Macrobrachium rosenbergii with ln-transformed data from Figure A to Figure H(SCL: second cheliped length; IL: ischium length; ML: merus length; CP: carpus length; PL: propodus length; DL: dactylus length; PW: propodus width; PH: propodus height; CL: carapace length)
2.3 性二型指数
根据罗氏沼虾第二步足两性变量特征值,计算出罗氏沼虾第二步足性二型指数(表2)。从表2可以看出,两性间第二步足长和第二步足重的SDI分别为1.39和2.90,其余变量间的SDI均大于1,表明罗氏沼虾第二步足相关变量均是雄性大于雌性(P<0.05)。

表2 罗氏沼虾第二步足的性二型指数Tab.2 Sexual dimorphism index of the second cheliped characters of Macrobrachium rosenbergii
3 讨论
3.1 罗氏沼虾第二步足的性二型
甲壳动物的性二型可以表现为体形[4-5]或者身体局部特征—如斑点[3]、螯足[6-9,27]等两性间的差异。性选择假说认为一种生物如果某个特征在与同性的配偶竞争中具有优势,并有利于提高交配成功率,这个优势特征将会被选择,从而产生性二型。性选择一般作用于雄性,使具备优势特征的雄性适合度更高,进而使这种身体特征在不断的同性竞争中得到强化。对于雄性罗氏沼虾而言,雄虾拥有较大的第二步足有助于维持其在社群中的优势地位,从而在领域维持、同性间的生殖竞争、繁殖期间实现同更多雌虾的交配等行为中发挥重要作用[13-14]。许多甲壳纲中的十足目动物螯足发育强大,且与身体明显不成比例,而与体形大小相比,武器,即螯足的大小更能体现雄性优先权的获取能力。如BC雄虾体重比OC雄虾少45%或25%,但第二步足长度比OC雄虾多10%、20%,则BC雄虾总能在与OC雄虾的战斗中取得胜利;既使BC雄虾的体重比OC少93%,但只要BC与OC的第二步足长度接近,那么BC在与OC的战斗中也有50%的胜率[28]。身体规格对雄性岸蟹(Carcinusmaenas)之间的争斗结果影响较小,与体形相比,武器大小是搏斗结果更可靠的预测指标,拥有更重、更长螯足的雄性更有可能赢得战斗[29]。对闪光活额寄居蟹(Diogenesnitidimanus)的研究表明,雄性之间在对雌性配偶的争夺斗争中,相对于体形来说,武器大小是决定战斗胜负的更重要因素[30]。
雄性通常以战斗的方式来确定个体在种群中的等级序列,以便有效夺取雌性交配权或维持领域地位。个体战胜种内竞争者的可能性同其相对于对手的绝对战斗能力或资源占有潜力(resource holding potential, RHP)密切相关,而RHP常常决定于个体的武器大小[30]。因此,在长期的自然进化过程中为竞争配偶而击败种内竞争者的特征会显著增强并得以更好进化。由于性选择是依据个体获得配偶的能力进行的,对于雄性个体来讲,作为武器部分的性征会进化的更加显著,因此性选择被认为是促使雄性动物比同性其他个体更具竞争力的某些身体器官(如作为武器的螯足)进化发展的主要动力。
3.2 罗氏沼虾两性第二步足的异速生长
在个体生长发育过程中,雌、雄个体的各形态特征会随着个体大小的增大而呈现出不同的生长速率。关于十足目甲壳动物螯足结构演变的研究常集中于种内的异速生长领域,因为甲壳动物的步足在种间和种内对资源的竞争以及争夺领域方面起着重要的作用,常被用作研究异速生长的最好实例[31]。Mossolin和Bueno[32]曾对欧式沼虾的第二步足异速生长进行了报道,研究结果表明雌、雄群体的掌节长-头胸甲长及掌节高-头胸甲长均表现为显著的正异速生长。对巴西沼虾(Macrobrachiumbrasiliense)构建的掌节长-头胸甲长及掌节高-头胸甲长的异速生长方程研究结果表明[33],掌节长无论是在雌性还是雄性群体中均表现为正异速生长,而掌节高在雄性群体表现为正异速生长,在雌性群体中则表现为等速生长。对日本沼虾(Macrobrachiumnipponense)第二步足长-体长的异速生长研究表明,无论是体长小于40 mm的性未成熟雄虾,还是体长大于40 mm的性成熟雄虾,第二步足相对于体长的异速生长速度都比雌虾快[34]。Gu等[35]曾对红螯螯虾螯足的异速生长进行了研究,建立的螯足宽-头胸甲长和螯足长-头胸甲长的两性异速生长方程揭示出,雄性的螯足宽和螯足长相对于体长的生长速率要快于雌性。对南太平洋寄居蟹的性二型研究表明[36],雄性的左掌节长-头胸甲长呈现高度正异速生长,而雌性的左掌节长-头胸甲长呈现等速或负异速生长。
本研究建立了性成熟罗氏沼虾第二步足长-第二步足重的两性生长方程,结果表明雌、雄虾第二步足重呈现正异速生长,即第二步足质量生长快于第二步足长度生长,且雄虾的第二步足质量的异速生长水平大于雌虾。此外还构建了第二步足长-头胸甲长、座节长-头胸甲长、长节长-头胸甲长、腕节长-头胸甲长、掌节长-头胸甲长、指节长-头胸甲长、掌节宽-头胸甲长及掌节高-头胸甲长的异速生长方程,结果显示雄性的第二步足长、座节长、长节长、腕节长、掌节长和指节长随头胸甲长的生长速率快于雌性,且雄性均呈现正异速生长,而雌性第二步足长、座节长、长节长和腕节长随头胸甲长的生长速率小于雄性,呈现负异速生长;掌节长和指节长随头胸甲呈现正异速生长,且异速生长水平小于雄性。此外,掌节宽和掌节高在两性间均表现为正异速生长,且两者异速生长水平相当,即雌、雄掌节宽和掌节高生长速率一致。与雌性相比,雄性第二步足更快的生长可保证身体的战斗器官优先发育,更好地在社群中建立优势地位,进而提高自身的繁殖适合度。
猜你喜欢
当代水产(2022年7期)2022-09-20
当代水产(2022年6期)2022-06-29
当代水产(2021年8期)2021-11-04
小读者(2021年2期)2021-03-29
小学阅读指南·低年级版(2020年9期)2020-10-12
阅读与作文(小学低年级版)(2019年11期)2019-12-26
华人时刊(2019年13期)2019-11-17
文苑(2018年22期)2018-11-19
红领巾·萌芽(2016年1期)2016-09-10
飞碟探索(2016年5期)2016-05-10
