
干旱胁迫对葛藤种苗生理特性的影响
2020-12-21 03:56谭景晨赵丽丽张淑炜席溢陈超
农业与技术 2020年22期
谭景晨 赵丽丽 张淑炜 席溢 陈超


摘 要:以葛藤为材料,研究不同渗透势(CK、-0.2MPa、-0.4MPa PEG-6000)下葛藤种苗的保护性酶活性(CAT、POD、SOD)及MDA、可溶性糖、可溶性蛋白含量的变化,拟揭示葛藤适应干旱的生理机制。结果表明:随着干旱胁迫的增强,CAT、POD活性呈现上升趋势;SOD活性呈先上升后下降趋势;MDA和可溶性蛋白含量都有所升高,而可溶性糖含量先降低后升高。说明干旱胁迫均有利于提高葛藤种苗活性氧清除系统中的CAT、POD和SOD的酶活性,提高可溶性蛋白和可溶性糖的含量,来缓解干旱胁迫对葛藤种苗造成的膜脂过氧化伤害。
关键词:葛藤;种子;干旱胁迫;保护酶活性
中图分类号:S-3 文献标识码:A DOI:10.19754/j.nyyjs.20201130006
葛藤(Pueraria lobata (Wild) Ohwi)是豆科蝶形花亚科多年生藤本植物,其根系发达,在表土层形成稠密的根网,在防止冲刷、崩塌、护坡固沟、保护堤岸、路基等方面有显著作用,为优良水土保持树种和贫瘠地区的绿化先锋植物[1]。葛藤还具有改良土壤作用,Gama等在油棕榈林地行间种植三裂叶野葛,不仅起到固氮的作用,而且可提高土壤中的钾、磷利用效率[2];Jayasinghe将葛藤作为橡胶园的覆盖物,用以减少水流冲蚀,增加了土壤中的生物活动,提高了固氮能力,对橡胶园中的土壤改良起到了良好的促进作用[3]。
中国是世界上岩溶区分布面积最大的国家之一,约占国土面积的1/3[4]。中国西南岩溶区表层岩溶带发育有大量溶槽、溶沟等孔隙结构,直接导致地高水低、雨多地漏、石多土少、土薄易旱的二元水文格局[5]。因此,干旱成为该区域严重影响植物正常生长发育,制约当地经济发展的主要因素之一[6]。鉴于此,本试验旨在利用PEG-6000模拟干旱胁迫,研究葛藤种苗的生理指标,探讨葛藤早期生长阶段应对干旱胁迫的生理变化,以期为今后系统、深入地研究葛藤抗旱调节机制及西南岩溶山区葛藤种植推广提供理论参考。
1 材料与方法
1.1 试验材料
由江苏园林绿化种子公司提供产地为澳大利亚的葛藤种子。
1.2 试验设计与方法
1.2.1 试验设计
利用聚乙二醇6000(PEG-6000)溶液模拟干旱胁迫,在室温下配制0.0MPa、-0.2MPa、-0.4MPa的渗透压梯度。选择大小均匀、籽粒饱满的葛藤种子用次氯酸钠溶液浸泡20min后用蒸馏水冲洗4~5次待用。将15cm玻璃培养皿提前灭菌并垫上2层滤纸,将次氯酸钠溶液浸泡后的种子随机摆在培养皿中,每皿50粒。并向各培养皿中加入30mL PEG-6000溶液,4个重复,然后置于25℃光照培养箱中,每日补足散失的水分。
1.2.2 测定内容与方法
待葛藤种子培养到第10天,每皿随机选取10株种苗,一同测定不同干旱胁迫下葛藤种苗的生理指标。过氧化氢酶(Catalase,CAT)活性以催化1μmol·min-1过氧化氢为1个CAT活性单位[7];超氧化物歧化酶(Superoxide dismutase,SOD)活性采用氮蓝四唑光还原法;过氧化物酶(Peroxidase,POD)活性采用愈创木酚法;丙二醛(Malondialdehyde,MDA)含量采用硫代巴比妥酸(TBA)法;可溶性糖含量采用蒽酮比色法;可溶性蛋白含量采用考马斯亮蓝法,以上各指标测定方法均按照相关试剂盒说明书进行,结果按说明书中公式计算,试剂盒由索莱宝生物科技有限公司提供。
1.3 数据的统计与分析
利用Excel 2016软件进行数据处理,采用GraphPad Prism 5软件作图,差异性分析采用SPSS 21.0软件。
2 结果与分析
2.1 干旱胁迫对葛藤种苗过氧化氢酶活性的影响
由图1可知,干旱胁迫显著影响了种苗的过氧化氢酶活性。当PEG为-0.2MPa时,与对照相比对葛藤种苗CAT活性影响不显著(P>0.05);当PEG为-0.4MPa时,与对照相比葛藤种苗的CAT活性显著提高(P<0.05)。即随着胁迫强度增加,CAT活性呈上升趋势。
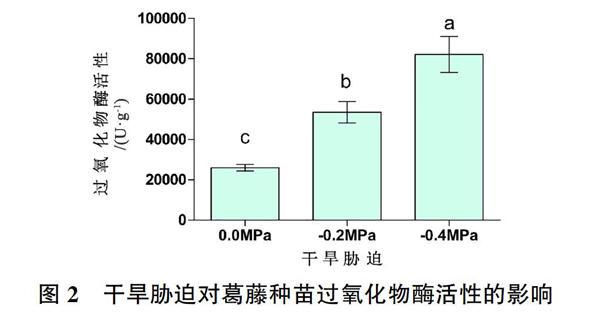
2.2 干旱胁迫对葛藤种苗过氧化物酶活性的影响
由图2可知,干旱胁迫显著影响了种苗的过氧化物酶活性。与对照组相比,PEG为-0.2MPa和-0.4MPa时,葛藤种苗的POD活性均显著提高(P<0.05);与-0.2MPa相比,-0.4MPa处理时葛藤种苗的POD活性也显著提高(P<0.05)。即随着干旱胁迫强度增加,POD活性呈上升趋势。
2.3 干旱胁迫对葛藤种苗超氧化物歧化酶活性的影响
由图3可知,干旱胁迫显著影响了葛藤种苗的超氧化物歧化酶活性。PEG为-0.2MPa和-0.4MPa时,与对照组相比葛藤种苗的SOD活性变化不显著(P>0.05);但与-0.2MPa相比,-0.4MPa时葛藤种苗的SOD活性显著降低(P<0.05)。即随着干旱胁迫强度增加,SOD活性呈先上升后降低的趋势。
2.4 干旱胁迫对葛藤种苗丙二醛含量的影响
由图4可知,干旱胁迫显著影响了种苗的丙二醛含量。PEG为-0.2MPa和-0.4MPa时,与对照组相比葛藤种苗的MDA含量影响均无显著变化(P>0.05);但与-0.2MPa相比,-0.4MPa时葛藤种苗的MDA含量显著升高(P<0.05)。即随着干旱胁迫强度增加,MDA含量呈先降低后上升的趋势。
2.5 干旱胁迫对葛藤种苗可溶性糖含量的影响
由圖5可知,干旱胁迫显著影响了种苗的可溶性糖含量。与对照组相比,PEG为-0.2MPa时葛藤种苗的可溶性糖含量显著降低(P<0.05);与对照组相比,PEG为-0.4MPa时对葛藤种苗的可溶性糖含量影响无显著变化(P>0.05);与-0.2MPa相比,-0.4MPa时葛藤种苗的可溶性糖含量显著提高(P<0.05)。即随着干旱胁迫强度增加,可溶性糖含量呈先降低后上升的趋势。
2.6 干旱胁迫对葛藤种苗可溶性蛋白含量的影响
由图6可知,干旱胁迫显著影响了种苗的可溶性蛋白含量。与对照组相比,PEG为-0.2MPa时葛藤种苗的可溶性蛋白含量影响无显著变化(P>0.05);与对照组相比,PEG为-0.4MPa时葛藤种苗的可溶性蛋白含量显著提高(P<0.05)。即随着干旱胁迫强度增加,可溶性蛋白含量呈上升趋势。
3 讨论与结论
水分在植物的生命活动中起着关键作用,据统计,全世界由于缺水造成的减产超过其它因素导致减产的总和[8]。种子萌发阶段的耐旱情况可在一定程度上反映该物种的耐旱程度[9]。过氧化氢酶(CAT)、过氧化物酶(POD)、超氧化物歧化酶(SOD)是植物细胞在长期进化过程中形成的主要的抗氧化酶类,可清除及减少植物体内的自由基,维持活性氧代谢平衡,有效减轻外界胁迫对植物细胞的伤害[10,11]。
CAT是植物遭受非生物胁迫时细胞内所产生的活性氧清除系统中的主要的抗氧化酶,可清除植物体内产生的H2O2,从而保护细胞不受伤害[12]。
POD广泛分布于植物组织细胞中,其活性能够在一定程度上反映植物对活性氧自由基的清除能力[13]。本研究表明,CAT、POD活性在干旱胁迫时随着胁迫强度增加显著上升(P<0.05),可能是因为随着干旱胁迫强度的增加,植物体内所有的活性氧清除酶系统和具生理抗性特征的生理活动被诱导而加快;CAT、POD在此诱导下活性逐渐增加,用以清除环境胁迫导致的植物体内过多的超氧阴离子自由基,这与张宇君等[14]对燕麦种子萌发的研究结果类似。
SOD可以催化超氧化物的歧化反应,是活性氧清除系统中第一个发挥作用的抗氧化酶。植物在逆境胁迫下会产生对细胞有害的活性氧和自由基,SOD通过催化歧化反应使活性氧生成过氧化氢和氧气,保护细胞避免或减轻活性氧伤害[15]。本研究表明,SOD活性在干旱胁迫时随着胁迫强度增加呈现先上升后下降的趋势,可能是因为前期SOD活性升高用以清除体内过多的活性氧自由基,而随着胁迫浓度进一步增大,SOD已经不能完全清除存在的活性氧自由基,平衡被打破,因此活性降低,这与杨利艳等[16]对藜麦种子萌发的研究结果类似。
丙二醛(MDA)是膜脂过氧化的产物,植物在受到外界环境刺激时会加剧膜脂过氧化程度,通常把MDA含量作为衡量植物在逆境下受害程度的指标[17]。本研究表明,与对照相比,无论PEG-6000为-0.2MPa还是-0.4MPa,干旱胁迫都对葛藤种子MDA含量影响不显著(P>0.05),说明保护酶降低了细胞内的自由基,一定程度上维持了活性氧代谢平衡,有效减轻了细胞膜脂过氧化。而与-0.2MPa相比,-0.4MPa时葛藤种子MDA含量显著增加(P<0.05),这是植物膜质过氧化水平加剧的表现,这与李新蕾等[18]对扁核木种子萌发的研究结果类似。
可溶性糖能够维持细胞的含水率和渗透势,保护生物大分子,降低因渗透失水所带来的损伤[19]。植物在逆境条件下通过积累可溶性糖等有机物来调节细胞渗透压,以增加适应环境的能力[20]。本研究表明,在PEG-6000为-0.2MPa时,与对照相比可溶性糖含量显著下降(P<0.05),这可能是因为干旱胁迫刺激葛藤的应激反應,使其可溶性糖消耗量升高。在PEG-6000为-0.4MPa时,与-0.2MPa时相比可溶性糖含量显著增加(P<0.05),这可能是因为葛藤体内保护酶系统作用使其适应该胁迫,从而可溶性糖含量升高。
蛋白质是生命的物质基础。植物在逆境条件下通过增加可溶性蛋白的合成,直接参与其适应逆境的过程[21]。本研究表明,随着胁迫浓度的增加,葛藤种子可溶性蛋白含量显著提高(P<0.05),这可能是因为在干旱胁迫诱导下,蛋白酶活性升高,促进贮藏蛋白水解为可溶性蛋白,增加了种苗中可溶性蛋白含量,这与付士磊等[22]、朱利君等[23]的研究结论相似。
本研究分析了葛藤种苗适应干旱的生理生化机制,发现在干旱胁迫均有利于提高葛藤种苗活性氧清除系统中的CAT、POD和SOD的活性,提高可溶性蛋白和可溶性糖的含量,来缓解干旱胁迫对葛藤种苗造成的膜脂过氧化伤害。
参考文献
[1]张海程,夏振强.石漠化治理新模式——种葛藤养豆丹[J].中国水土保持,2017(08):39-41.
[2]Gama A C D,Cadima A.Effect of NPK application and burning on the root system of oil palm in a yellow latosol in Bahia,Brazil [J].Turrialba,1990(8):368-373.
[3]Jayasinghe C K.The role of leguminous cover crops in soil improvement with special reference to the nitrogen economy of tropical rubber soils [J].Bulletin of the Rubber Research Institute of Sri Lanka,1991(6):23-26.
[4]袁道先.新形势下我国岩溶研究面临的机遇和挑战[J].中国岩溶,2009,28(04):329-331.
[5]倪隆康,顾大形,何文,黄玉清,陈中义.岩溶区植物生态适应性研究进展[J].生态学杂志,2019,38(07):2210-2217.
[6]陈莹,王普昶,赵丽丽,杨春燕,宋高翔.外源钙对干旱胁迫下木豆种苗生理特性的影响[J].草地学报,2014,22(05):1051-1055.
[7]孔祥生,易现峰.植物生理学实验技术[M].北京:中国农业出版社,2008:259-260.
[8]孙梅霞,祖朝龙,徐经年.干旱对植物影响的研究进展[J].安徽农业科学,2004(02):365-367,384.
[9]陈莹,钟理,赵丽丽,王普昶,莫本田,龙忠富.截叶铁扫帚种子萌发期对岩溶生境高钙干旱的生理生化反应[J].江苏农业科学,2014,42(09):335-339.
[10]Yin H, Chen Q, Yi M.Effects of short-term heat stress on oxidative damage and responses of antioxidant system inLilium longiflorum [J].Plant Growth Regulation,2008,54(1):45-54.
[11]Edreva A.Generation and scavenging of reactive oxygen species in chloroplasts: a submolecular approach[J].Agriculture Ecosystems & Environment,2005,106(2):119-133.
[12]蓋胜男,赵训超,魏玉磊,徐晶宇.玉米过氧化氢酶(CAT)基因家族生物信息学分析[A].中国作物学会.2019年中国作物学会学术年会论文摘要集[C].中国作物学会:中国作物学会,2019:1.
[13]郭慧娟,胡涛,傅金民.苏打碱胁迫对多年生黑麦草的生理影响[J].草业学报,2012,21(01):118-125.
[14]张宇君,赵丽丽,王普昶,陈超.盐旱交互对燕麦种子萌发及幼苗生理特性的影响[J].草业学报,2018,27(05):141-152.
[15]夏民旋,王维,袁瑞,邓粉妮,沈法富.超氧化物歧化酶与植物抗逆性[J].分子植物育种,2015,13(11):2633-2646.
[16]杨利艳,杨小兰,朱满喜,杨雅舒,王创云.干旱胁迫对藜麦种子萌发及幼苗生理特性的影响[J].种子,2020,39(09):36-40.
[17]刘俊祥,魏树强,翟飞飞,李伟,周晓星,孙振元.Cd2+胁迫下多年生黑麦草的生长与生理响应[J].核农学报,2015,29(03):587-594.
[18]李新蕾,李叶芳,李凤荣,关文灵.干旱胁迫对扁核木种子萌发及幼苗生理特性的影响[J].云南农业大学学报(自然科学),2020,35(04):682-687.
[19]肖泽华,李欣航,潘高,吴耀文,杨灿鑫,匡雪韶,刘文胜.锰胁迫对黄花草种子萌发及幼苗生理生化特征的影响[J].草业学报,2019,28(12):75-84.
[20]刘家尧,衣艳君,张其德.盐胁迫对不同抗盐性小麦叶片荧光诱导动力学的影响[J].植物学通报,1998,15(02):47-50.
[21]张明生,谢波,谈锋,张启堂.甘薯可溶性蛋白、叶绿素及ATP含量变化与品种抗旱性关系的研究[J].中国农业科学,2003(01):13-16.
[22]付士磊,周永斌,何兴元,陈玮.干旱胁迫对杨树光合生理指标的影响[J].应用生态学报,2006(11):2016-2019.
[23]朱利君,闫秋洁.外源氯化钙对大蒜幼苗盐胁迫伤害的缓解作用[J].江苏农业科学,2016,44(08):242-244.
(责任编辑 李媛媛)
收稿日期:2020-10-27
基金项目:贵州省科技计划项目(项目编号:黔科合支撑[2018]2258号、黔科合支撑[2019]2295号)
作者简介:谭景晨(1997-),男,硕士在读;通讯作者席溢。
猜你喜欢
润·文摘(2021年9期)2021-09-22
儿童时代·幸福宝宝(2019年9期)2019-10-28
山东农业科学(2016年11期)2016-12-17
现代园艺(2016年2期)2016-03-15
少儿科学周刊·儿童版(2015年1期)2015-07-07
