
灌浆初期高温影响水稻籽粒碳氮代谢的机理*
2020-12-18 02:15:40王军可王亚梁陈惠哲张义凯朱德峰张玉屏
中国农业气象 2020年12期
王军可,王亚梁,陈惠哲,向 镜,张义凯,朱德峰,张玉屏
(中国水稻研究所/水稻生物学国家重点实验室,杭州 311400)
气候变暖导致全球温度升高是近年来气候变化的明显特征[1],随着全球气温升高,水稻高温热害频发,尤其南方水稻在灌浆期遇到高温已成为制约稻米品质的重要因素。水稻灌浆结实期最适平均温度为21℃左右,35℃以上高温则会显著降低稻米的产量和品质[2-3],籽粒碳氮代谢的生理过程对籽粒灌浆起着关键作用。作为籽粒干重的重要组分,淀粉和蛋白质的代谢受碳氮代谢生理过程影响[4−5]。
高温影响籽粒碳氮代谢,淀粉合成受阻及垩白含量显著增加是高温影响稻米品质的显著特征[6−7]。作为叶片光合产物向各器官运输的主要形式,蔗糖是籽粒中淀粉合成的主要碳前体,蔗糖转运体(SUTs)在通过质外体途径转运蔗糖的过程中起重要作用[8−9]。高温影响蔗糖代谢过程,Zhong等[10]研究指出,32℃以上高温导致稻米食味和加工品质下降与高温抑制淀粉和蔗糖代谢存在关系,Zhang等的[11]研究指出,高温通过破坏韧皮部胞间连丝而抑制蔗糖转运。陈燕华等[12]在穗发育的研究中指出,40℃高温显著降低蔗糖转运蛋白相关基因的表达,然而高温对籽粒中蔗糖转运基因表达的影响需进一步研究。淀粉的进一步合成依赖于ATP与各种酶促反应的协调进行,高温降低淀粉合成关键酶(ADPG焦磷酸化酶、可溶性淀粉合成酶、颗粒结合型淀粉合成酶及去分支酶活性)活性导致淀粉合成降低已成为基本共识[13−14],而对籽粒中能量代谢与淀粉合成关系的研究相对较少。糖酵解和三羧酸循环是主要的产能途径,α-酮戊二酸作为三羧酸循环重要的中间产物,也参与氮代谢中谷氨酸的合成,研究表明[15],高温抑制了穗发育过程中糖酵解和三羧酸循环过程中关键酶的活性,而在籽粒中这种现象是否存在并不清楚。高温促进蛋白质的合成[16],但高温下氮代谢与三羧酸循环的关系并不清楚。转录组分析表明,灌浆期高温影响表达的基因包括抑制蔗糖水解和淀粉合成,同时促进淀粉水解和储藏蛋白质合成[17−18]。然而蛋白质和淀粉是否受到统一途径代谢的影响并不清楚。为此,本研究拟以长江中下游优质粳稻浙禾香2号为材料,选择水稻种子发育5、10及15d这3个时期,在常温和高温处理下,通过籽粒发育过程中淀粉和蛋白积累的变化,分析蔗糖转运及分解、糖酵解及三羧酸循环关键酶活性、淀粉代谢关键酶活性以及氮代谢关键酶活性,以期深化籽粒灌浆期高温热害理论,明确高温对碳氮代谢的影响,并为稻米品质的调控提供参考。
1 材料与方法
1.1 试验设计
试验供试品种为嘉兴市农业科学院选育的优质稻品种浙禾香2号,于2019年在中国水稻研究所富阳试验基地进行盆栽试验,塑料种植盆规格为24cm×22.5cm×21.5cm,每盆装过筛土8kg。5月25日播种,大棚基质育秧,6月15日选取生长状况一致的秧苗进行移栽,每盆2穴,每穴2苗,共120盆。每盆施复合肥(N∶P∶K=15%∶15%∶15%)4.0g作基肥,而后施0.5g尿素作分蘖肥,穗分化始期施2.25g复合肥作穗肥,水稻生长期间其它管理措施与高产栽培一致。
1.2 温度处理及籽粒样本采集
设置适温(NT)和高温(HT)两个处理,均在人工气候箱(PGV-36型CONVIRON)内进行,具体温度设置如表1。人工气候箱具有自动控温控湿的功能,气候箱透光面所用材质为超透明的超白钢化玻璃,具有低自爆率、颜色一致、可见光透过率高及通透性好等特点,其光照以外界自然光为主,光谱损失少,基本保持箱内光照与外界自然光照一致,同时人工气候箱底部有鼓风装置,以此保证二氧化碳浓度与室外基本一致。箱内环境条件能达到试验处理要求。在水稻灌浆始期当天正午移入气候箱开始进行高温和适温处理,共持续15d,结束后移至自然环境下生长至成熟,收获时间为10月21日。
移入气候箱后选择大小基本一致的单穗挂牌标记,每处理标记80穗,供取样分析。温度处理期间,分别于5d(第6天)、10d(第11天)、15d(第16天)和成熟收获时,各取稻穗20个,将穗子按长度三等分,留取穗中部籽粒作为样本,用铝箔纸包裹,经液氮冷冻后,装入自封袋,置于−80℃冰箱保存,然后统一测定。

表1 人工气候箱温度设置Table 1 Temperature setting of different treatments in climate chamber
1.3 项目测定
1.3.1 粒重
在每次收集的籽粒样本中,各取大小均匀的籽粒50颗去壳,置于电热恒温鼓风干燥箱(DHG-9070A)中105℃杀青30min,80℃烘至恒重,后用德国sartoriys BS 224S电子天平测定粒重,算术平均计算单粒重。
1.3.2 蔗糖代谢关键酶活性及相关代谢物含量
在每次收集的籽粒样本中,各取大小均匀的冷冻去壳籽粒0.1g检测。蔗糖含量测定采用蒽酮比色法[19],葡萄糖和果糖含量、可溶性酸性转化酶和蔗糖合酶活性利用苏州某公司提供试剂盒进行测定,重复3次,取平均值。
1.3.3 能量代谢关键酶活性及相关代谢物含量
在每次收集的籽粒样本中,各取大小均匀的0.1g冷冻籽粒,利用己糖激酶、丙酮酸激酶、柠檬酸合酶、异柠檬酸脱氢酶、α-酮戊二酸脱氢酶、琥玻酸脱氢酶及苹果酸脱氢酶活性试剂盒进行测定,利用ATP和ADP含量测定试剂盒进行含量测定,试剂盒由苏州某公司提供,重复3次,取平均值。
1.3.4 淀粉代谢关键酶活性及相关代谢物含量
在每次收集的籽粒样本中,各取大小均匀的0.1g冷冻籽粒。腺苷二磷酸葡萄糖(ADPG)含量采用双抗体夹心法利用ELISA试剂盒测定,测定原理为,用纯化的植物腺苷二磷酸葡萄糖(ADPG)捕获抗体包被微孔板,制成固相抗体,往包被的微孔中依次加入植物腺苷二磷酸葡萄糖(ADPG),再与HRP标记的检测抗体结合,形成抗体−抗原−酶标抗体复合物,经过彻底洗涤后加底物TMB显色。TMB在HRP酶的催化下转化成蓝色,并在酸的作用下转化成最终的黄色。颜色的深浅和样品中的植物腺苷二磷酸葡萄糖(ADPG)呈正相关。用酶标仪在450nm波长下测定吸光度(OD值),通过标准曲线计算样品中植物腺苷二磷酸葡萄糖(ADPG)含量。利用腺苷二磷酸葡萄萄糖焦磷酸化酶(AGP)、颗粒结合型淀粉合成酶、可溶性淀粉合成酶、淀粉分支酶、淀粉去分支酶、α-淀粉酶、β-淀粉酶及淀粉磷酸化酶活性测定试剂盒(苏州某公司提供)进行测定。淀粉含量测定参考Hansen等[20]的方法。各指标测定均3次重复,取平均值。
1.3.5 氮代谢关键酶活性及相关代谢物含量
全氮含量测定:在每次收集的籽粒样本中,各取烘干后的籽粒0.2g,通过全自动样品快速研磨仪(Tissuelyser-48,上海)磨成细粉,采用H2SO4- H2O2消煮,并用FOSS全自动凯氏定氮仪测定含氮量。
蛋白质含量测定:参考GB 5009.5-2016食品安全国家标准食品中蛋白质的测定方法。
其它氮代谢相关指标测定:选取0.1g冷冻籽粒。利用氨基酸含量、α-酮戊二酸含量、亚硝酸还原酶活性、硝酸还原酶活性、谷氨酰胺合成酶活性及谷氨酸合酶活性试剂盒进行相关指标含量和活性的测定。各指标测定均3次重复,取平均值。
1.3.6 籽粒RNA提取和实时荧光定量
取冷冻保存的籽粒样品0.1g,用于蔗糖转运蛋白相关基因OsSUT1和OsSUT2相关表达量的测定。籽粒总RNA参照Trizol法[21]提取,利用TOYOBO的Rever Tra Ace quantitative PCR RT Master Mix(Toyobo, Osaka, Japan)试剂盒反转录。荧光定量PCR所采用的设备为7500 Real Time PCR System(Applied Biosystems System)。反转录后的产物稀释后作为模板cDNA。用内参基因OsUBQ引物检测反转录产物,引物采用Primer5软件设计,基因表达分析所用引物序列如表2所示;根据样品特定荧光阈值的Ct值,采用2−ΔΔCt法[22]计算不同样品间基因的相对表达量。
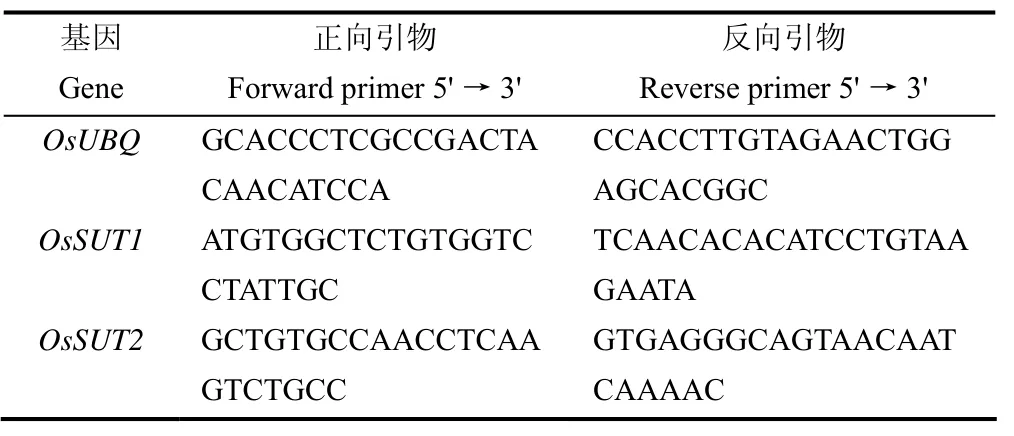
表2 实时荧光定量PCR引物序列Table 2 Primers used for quantitative real-time PCR
1.4 数据分析
采用Microsoft Excel统计数据,用SAS9.1软件进行数据分析,处理间的差异分析采用student单尾t检验法。
2 结果与分析
2.1 高温对籽粒重及其淀粉/蛋白质含量的影响

图1 灌浆初期高温(HT)与适温(NT)处理持续5d、10d和15d后采样以及成熟时采样观测的籽粒重及其淀粉/蛋白质含量的比较Fig.1 Comparison of grain weight and its starch/protein content sampled after 5d, 10d and 15d treatment, during the early stage of filling and maturity stage between high temperature(HT)and normal temperature(NT)
由图1可知,高温抑制水稻籽粒干物质积累,但对淀粉和蛋白质积累的影响存在差异。高温处理15d导致成熟期籽粒粒重降低16.9%,处理期间高温下淀粉相对含量降低,蛋白质相对含量却呈现上升的趋势。
2.2 高温对籽粒碳代谢的影响
2.2.1 对蔗糖转运的影响
蔗糖是籽粒中淀粉积累的重要来源,蔗糖的转运与卸载对淀粉合成有重要意义,作为蔗糖转运家族的重要成员,OsSUT1和OsSUT2在籽粒灌浆过程中起着重要作用;葡萄糖和尿苷二磷酸葡萄糖(UDPG)是籽粒中淀粉合成的基础物质,其来源于光合产物蔗糖的分解。
由图2可见,与适温(31℃/24℃)条件下相比,灌浆初期高温(35℃/28℃)持续5d、10d和15d后,籽粒中蔗糖转运基因OsSUT1(图2a)和OsSUT2(图2b)的相对表达量均极显著降低,而且高温处理超过10d后,OsSUT2的相对表达量下降更多,从而造成高温处理5d、10d和15d后籽粒中蔗糖含量极显著低于同时段适温条件下(图2c)。可见,灌浆初期高温使籽粒发育过程中蔗糖供应受到明显抑制。
而由图3可见,高温条件下持续5d和10d处理中,籽粒中可溶性酸性转化酶活性(S-AI)(图3a)和蔗糖合成酶(SS-I)活性(图3b)极显著高于适温处理(P<0.01),持续15d后该两种酶的活性明显下降,与适温处理一致或明显降低。SS-I主要催化蔗糖和UDP生成游离果糖和UDPG,而S-AI催化蔗糖不可逆地分解为葡萄糖和果糖,由图3c、图3d可见,高温条件下持续5d和10d处理中籽粒中果糖含量明显高于适温处理,持续15d后果糖含量与适温处理差异不显著;而高温条件下各处理时段籽粒中葡萄糖含量均明显高于适温处理。可见,灌浆初期高温使籽粒发育过程中S-AI和SS-I活性增加,加速了蔗糖向UDPG和葡萄糖的转化。
2.2.2 对淀粉代谢的影响
蔗糖的分解产物在淀粉体中经ADPG焦磷酸化酶(AGPase)催化形成淀粉的直接合成前体腺苷二磷酸葡萄糖(ADPG),后经颗粒结合淀粉合成酶(GBSS)、可溶性淀粉合成酶(SSS)、淀粉分支酶(SBE)、淀粉去分支酶(DBE)等一系列酶的作用下分别合成直链淀粉和支链淀粉。
由图4可见,随着高温处理时间的延长,AGPase活性降低,而ADPG含量却呈现上升的趋势,同时GBSS和SSS活性显著下降,说明直链淀粉合成受阻。同时随着处理时间的延长,SBE活性先升高后降低,DBE活性呈现先下降后上升的趋势。说明灌浆前期15d高温对淀粉合成的影响主要通过降低AGPase、GBSS、SSS和SBE的活性以及前10d的DBE的活性来降低淀粉的合成,高温处理下ADPG含量上升的原因是其进一步合成淀粉的途径受阻。

图2 高温(HT)与适温(NT)处理籽粒蔗糖转运基因表达量(a、b)和蔗糖含量(c)的比较Fig.2 Comparison of the expression of sucrose transporter genes(a,b) and sucrose content(c) in grains treated with high temperature (HT) and normal temperature (NT)

图3 高温(HT)与适温(NT)处理籽粒可溶性酸性转化酶活性(a)、蔗糖合成酶活性(b)、果糖含量(c)以及葡萄糖含量(d)的比较Fig.3 Comparison of soluble acid invertase activity(a), sucrose synthase activity(b), fructose content(c) and glucose content(d) in grains treated with high temperature (HT) and normal temperature (NT)

图4 高温(HT)与适温(NT)处理籽粒AGPase(a)、GBSS(b)、SSS(c)、SBE(d)、DBE活性(e)及ADP-葡萄糖含量(f)的比较Fig.4 Comparison of the activity of AGPase(a), GBSS(b), SSS(c), SBE(d), DBE(e) and the ADPG(f) content in grains treated with high temperature (HT) and normal temperature (NT)
淀粉合成的同时也在不断地被利用,α-淀粉酶、β-淀粉酶和淀粉磷酸化酶是主要的淀粉降解酶。由图5可见,与适温处理相比,随着高温处理时间的延长,ɑ-淀粉酶,β-淀粉酶及淀粉磷酸化酶活性呈上升的趋势,说明高温促进了籽粒淀粉水解。
可见,灌浆初期高温使籽粒发育过程中淀粉合成减少,而淀粉的水解增强,最终导致淀粉含量的降低。
2.3 高温对水稻籽粒糖酵解及三羧酸循环关键酶活性的影响
ATP参与ADPG和淀粉的合成,淀粉体中ATP的含量与ADPG和淀粉含量呈正相关[23],糖酵解及三羧酸循环是主要的产能过程。对不同温度处理下籽粒中糖酵解及三羧酸循环关键酶的分析可知,高温下糖酵解关键酶活性受到抑制,随着处理时间的延长,己糖激酶和丙酮酸激酶活性显著下降(图6),说明糖利用受阻。由图7可见,三羧酸循环关键酶活性的变化在高温下表现出明显的差异,柠檬酸合酶、α-酮戊二酸脱氢酶及琥珀酸脱氢酶活性显著下降,而随着处理时间的延长,异柠檬酸脱氢酶和苹果酸脱氢酶活性显著升高。高温下ATP和ADP总量与适温下并无显著差异。
可见,灌浆初期高温致使能量代谢紊乱,最终导致高温下ATP和ADP总量与适温下并无显著差异。

图5 高温(HT)与适温(NT)处理籽粒α-淀粉酶(a)、β-淀粉酶(b)以及淀粉磷酸化酶(c)活性的比较Fig.5 Comparison of the activity of α-amylase(a), β-amylase(b) and starch phosphorylase(c) in grains treated with high temperature (HT) and normal temperature (NT)

图6 高温(HT)与适温(NT)处理己糖激酶活性(a)和丙酮酸激酶(b)活性的比较Fig.6 Comparison of the activity of hexokinase(a) and pyruvate kinaset(b) in grains treated with high temperature (HT) and normal temperature (NT)

图7 高温(HT)与适温(NT)处理籽粒三羧酸循环关键酶活性(a−e)及ATP和ADP含量总和(f)的比较Fig.7 Comparison of the activity of key enzymes in the tricarboxylic acid cycle(a-e),sum of ATP and ADP content(f) in grains treated with high temperature (HT) and normal temperature (NT)
2.4 高温对水稻籽粒氮代谢的影响
硝酸还原酶(NR)、亚硝酸还原酶(NiR)、谷氨酰胺合成酶(GS)和谷氨酸合成酶(GOGAT)是氮素同化过程中的关键酶,其活性显著影响着蛋白质含量。α-酮戊二酸是三羧酸循环过程的中间产物,也是氮代谢中形成谷氨酸的重要原料,谷氨酸则可转化成其它氨基酸和酰胺进而形成蛋白质。
由图8可见,随着高温处理时间的延长,籽粒总氮含量较常温处理有上升趋势,但差异未达显著水平。α-酮戊二酸含量显著增加,使谷氨酸的合成原料增加。同时谷氨酸和氨基酸含量在高温处理下显著提高。图9显示,与适温处理相比,随着高温处理时间的延长,硝酸还原酶和亚硝酸还原酶活性呈现下降的趋势,使谷氨酸的另一合成原料氨减少,而高温下谷氨酸合成酶和谷氨酰胺合成酶活性与适温相比差异不明显。可见,灌浆初期高温使籽粒发育过程中异柠檬酸脱氢酶活性增加和α-酮戊二酸活性的降低,加速了α-酮戊二酸的积累,促进了α-酮戊二酸向谷氨酸的转换,加速了籽粒中形成蛋白质的原料即氨基酸含量的增加,进而导致蛋白质含量升高。
3 讨论与结论
本实验条件下,高温抑制水稻籽粒发育,导致干物质积累显著下降。试验表明,随着高温处理时间的延长,淀粉含量显著下降,而蛋白质含量显著上升,导致稻米品质变差,这与前人的研究结论是一致的[24−25]。但高温下淀粉和蛋白质的绝对含量低于适温处理,说明高温对籽粒的物质代谢造成影响。碳代谢方面,籽粒干物质积累主要来源于光合作用的蔗糖供应,试验发现,高温下蔗糖转运表达基因OsSUT1和OsSUT2表达量下调,说明转运卸载到籽粒中的蔗糖受到抑制,Zhang等[11]研究表明,高温下籽粒干物质积累受阻的主要原因是蔗糖分解受阻,而在本研究中,随着处理时间的延长,与适温相比,可溶性酸性转化酶和蔗糖合成酶在高温下呈现先上升后下降的趋势,可见蔗糖分解受阻并不是干物质积累降低的主要原因。与前人研究结果的差异可能是由于设置处理温度不同导致,不同温度条件下,蔗糖转运和卸载对籽粒干物质积累的影响程度有待进一步明确。

图8 高温(HT)与适温(NT)处理籽粒全氮(a)、α-酮戊二酸(b)、谷氨酸(c)和氨基酸(d)含量的比较Fig.8 Comparison of the content of total nitrogen(a), ɑ-Ketoglutarate(b), glutamate(c)and amino acid(d) in grains treated with high temperature (HT) and normal temperature (NT) treatments

图9 高温(HT)与适温(NT)处理籽粒硝酸还原酶(a)、亚硝酸还原酶(b)、谷氨酰胺合成酶(c)及谷氨酸合酶(d)活性的比较Fig.9 Comparison of the activities of nitrate reductase (a), nitrite reductase (b), glutamine synthetase (c) and glutamate synthase (d) in grains treated with high temperature (HT) and normal temperature (NT)
高温下葡萄糖和果糖含量有所上升,对糖酵解关键酶己糖激酶和丙酮酸激酶活性进行分析发现, 高温下两种酶活性均呈显著下降趋势,说明糖利用受阻。另一方面,试验发现,高温导致淀粉水解酶活性增加,这也是葡萄糖和果糖含量增加的原因之一,研究表明淀粉水解导致可溶性糖积累可以提高籽粒的抗氧化能力[26−27]。能量代谢方面,本实验中三羧酸循环过程关键酶活性在高温下的变化不一致,可能是由反馈调节导致,作为三羧酸循环重要的中间产物,α-酮戊二酸的代谢与蛋白质合成密切相关,异柠檬酸脱氢酶活性上升促进了α-酮戊二酸的合成,同时α-酮戊二酸脱氢酶活性受到抑制减少了α-酮戊二酸的利用,进一步提高了α-酮戊二酸含量。虽然高温导致三羧酸循环代谢紊乱,但ATP和ADP含量在高温下并没有发生显著变化,说明能量供应并未受阻,α-酮戊二酸含量的增加也进一步促进氮代谢谷氨酸形成。氮代谢方面,高温下氮代谢相关酶活性受高温的影响较淀粉代谢相关酶活性受高温的影响小,高温下谷氨酸合成酶和谷氨酰胺合成酶活性受高温影响不大,与梁成刚等[28]的结果有所差异,原因可能是不同遗传背景材料的氮代谢过程对温度的响应不同。试验表明,α-酮戊二酸含量增加是蛋白质相对量积累提高的原因之一。试验中,与适温处理相比,高温下氨基酸积累显著增加,一方面与氨基酸合成增加有关,另一方面,可能由于高温促进了氨基酸向籽粒的转运,试验发现高温下铵态氮和硝态氮转运相关基因表达量较低,更详细的氨基酸转运途径有待于进一步研究。
综上所述,高温主要通过阻碍蔗糖转运抑制籽粒发育,同时高温导致糖酵解和三羧酸循环紊乱;高温下籽粒淀粉含量下降的主要原因是淀粉合成受阻且淀粉水解增强;高温会导致三羧酸循环过程紊乱,α-酮戊二酸积累增加是蛋白质含量上升的原因之一。α-酮戊二酸参与的能量代谢的相关研究可为高温下稻米品质的改良提供新思路,α-酮戊二酸能否作为外源调节剂减弱灌浆前期高温对水稻生产的影响,还有待进一步研究。
猜你喜欢
云南化工(2021年10期)2021-12-21 07:33:28
建材发展导向(2021年24期)2021-02-12 02:00:02
广西糖业(2020年3期)2020-09-25 08:32:44
浙江工业大学学报(2017年5期)2018-01-22 02:03:33
三门峡职业技术学院学报(2017年1期)2017-06-05 10:17:30
中国洗涤用品工业(2017年2期)2017-04-16 05:07:45
中国比较医学杂志(2017年5期)2017-01-17 06:17:05
中国糖料(2016年1期)2016-12-01 06:49:06
医学研究杂志(2015年12期)2015-06-10 06:57:46
中国果业信息(2015年11期)2015-01-23 19:19:58
