
辣椒疫病致病因子及防治研究进展
2020-11-30 09:00彭玉梅黄玲芝裴冬丽韩霜刘冬梅周庆峰朱晓琴
中国瓜菜 2020年9期
彭玉梅 黄玲芝 裴冬丽 韩霜 刘冬梅 周庆峰 朱晓琴
摘 要:辣椒(Capsicum annuum)是世界上重要的蔬菜作物之一,然而辣椒疫霉菌(Phytophthora capsici)引起的辣椒疫病给我国乃至世界的辣椒生产造成了严重的经济损失。目前,对辣椒疫病的有效防治方法还很有限,主要是化学药物防治,往往造成辣椒果的农药残留和环境污染。随着科学技术的进步,人们对辣椒疫病的研究已经深入到分子水平,以期从分子水平研究辣椒疫病抗性品种。综述了国内外关于辣椒疫病致病因子研究进展情况,并简述了辣椒疫病的检测及防治方面的研究现状,以期为辣椒疫病的分子育种研究与科学防治提供理论参考。
关键词:辣椒疫病;致病因子;检测方法;防治措施
中图分类号:S641.3 文献标志码:A 文章编号:1673-2871(2020)09-007-05
Abstract: Pepper is one of the most important crops in China. But pepper phytophthora blight (Phytophthora capsica) causes serious economic losses to the pepper production in China and even in the world. With the advance of research, the research on pepper phytophthora blight had reached the molecular level. This paper summarized the research progress, detection methods, and preventive/control measures of pepper blight.
Key words: Pepper phytophthora blight; Pathogenic factors; Detection methods; Preventive measures
辣椒属于茄科辣椒属,是一种原产于南美洲的一年生草本植物,果实成熟时通常会变成红色,含有较多的维生素C,味道鲜美,是不可或缺的美食原料之一。辣椒疫病是一种具有高度破坏性的植物病害,主要经土壤传播,其致病菌是辣椒疫霉菌,于1918年首次在美国发现,现普遍分布于全球,在我国很多地方也都有发现。该病害可侵染辣椒叶、果实、茎等部位,引发整株萎蔫甚至死亡。如若防治不彻底,会造成巨大的经济损失乃至绝收,這将对辣椒生产造成很大的影响[1]。为了减少辣椒疫病带来的损失,人们进行了诸多试验研究,在对该病的致病机制研究方面,因为该病原菌可以侵染本氏烟和拟南芥,故人们常利用这两种模式植物来进行相关分子水平试验。由于病原体的扩散途径复杂多样,病害的发生通常呈集中暴发性,并且单一的控制方法又往往无法达到预期效果[1]。因此,从植株与病原菌互作的角度,尤其是从分子水平切入,继而深入研究发病机制,探究多种措施相结合的综合防治策略,就显得尤为重要。笔者从辣椒疫病致病因子、检测方法及防治措施几方面综述了辣椒疫病的研究进展,以期为该病害的防治提供理论参考。
1 致病因子
1.1 RXLR效应子
为了阻断病原微生物的侵染,植物体内也进化出相应的免疫防卫系统。首先是基本的防御反应,病原体相关分子模式PAMP(pathogen-associated molecular pattern)可以激发植株产生先天免疫反应PTI(PAMP triggered immunity),该反应可以阻止多种类型病原物的侵染;与此同时,病原菌又会通过分泌效应子的形式来抑制植物的PTI反应,于是,植株也进化出了第二种特异性的防御反应,即植物识别病原菌的效应子后会产生相应的效应蛋白如NB-LRR类抗性蛋白(resistance protein,R),并由此激发植株产生后天诱导反应ETI (effector triggered immunity)[2],这时往往还会激起植株的超敏反应(hypersensitive reaction,HR);病原物则又通过效应子抑制ETI反应[2]。
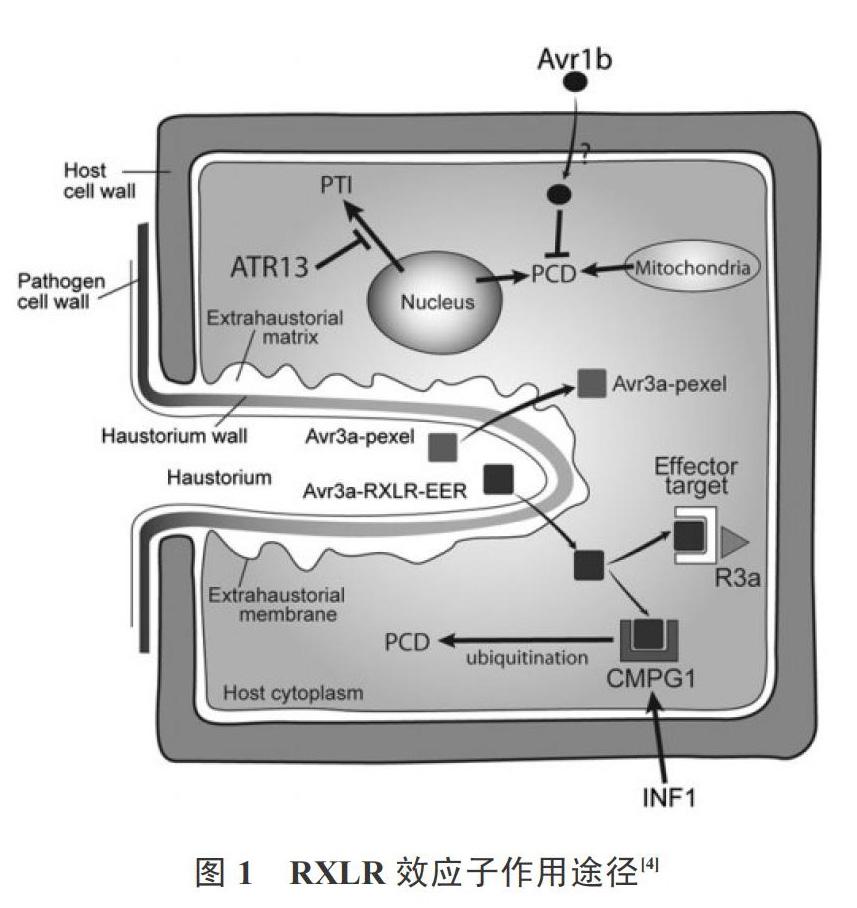
辣椒疫霉菌是一种植物病原卵菌,在侵染宿主时会通过RXLR效应子来抑制植物的防御反应,因此,对这些效应子进行深入研究可为防治辣椒疫病奠定基础[3]。RXLR效应子的N端是相对较保守的RXLR基序,主要功能是将效应蛋白转运到宿主细胞内,RXLR-EER基序是效应子进入宿主细胞所必需的(图1)[4-5],RXLR基序两侧是具有明显保守性的包含有数十个氨基酸的序列,而位于效应子C端区域的氨基酸序列变异性相对较大,后续深入研究发现该区域也具一定的保守域[6]。RXLR效应子在疫霉菌的进化和侵染植株阶段起着至关重要的作用,因此,深入了解RXLR效应子为研究辣椒疫病提供了有利的理论依据[7]。笔者选取了4种RXLR效应子进行分析,以期为辣椒疫病的研究提供参考。
陈孝仁等[8]的研究表明,RXLR型效应子PcAvh2在辣椒疫霉菌侵染植株的过程中不可或缺。其研究结果表明,PcAvh2具有10个等位基因,其编码的多肽在数十个氨基酸位点有多态性;在辣椒疫病发病早期阶段,该基因会特异上调表达,能抑制各种效应子引起的植物防御反应;并且,在原生质体转化试验中,如若沉默PcAvh2,则会显著降低辣椒疫霉的侵染力,说明PcAvh2在辣椒疫霉菌的侵染过程中起重要作用。
李秀奇等[3]应用从辣椒疫霉菌株中克隆获得的RXLR22034效应子进行蛋白质的表达与纯化试验,结果表明,在37 ℃和0.5 mmol·L-1 IPTG的诱导条件下,该基因在21 kDa处有大量蛋白表达,利用镍柱亲和层析及分子筛层析以纯化蛋白,最后再利用坐滴法得到较高质量的蛋白晶体。该研究为深入研究RXLR效应子的三维结构提供了重要的内容,有利于进一步研究辣椒疫霉RXLR效应子的功能。
杨楠等[7]分析研究了辣椒疫霉菌的RXLR121504效应子。试验结果表明,该效应子核酸序列381 bp,其编码产物包括126个氨基酸,其中信号肽包括24个氨基酸序列,第28~31位氨基酸是RXLR基序。研究结果还表明,该效应子在激活寄生植株超敏反应的同时,还能阻止由INF1激活的植物细胞坏死,即RXLR121504能激活或阻止植物的防御反应。该效应子类似于PiAvr3a,能结合并稳定细胞中的E3泛素连接酶,从而阻止由INF1激发的植物细胞死亡,协助辣椒疫霉菌的侵染[9]。由此推测RXLR121504在病原菌侵染寄主植株的过程中起着关键作用,这对探究RXLR121504的作用机制提供了有利的参考。
张丽等[10]通过实时荧光定量PCR技术来检测在辣椒疫霉菌侵染过程中效应子RXLR115890的表达情况,结果表明,RXLR115890在病原菌侵染植物前期的表达量显著上调,但随着时间的推移,其表达量很快又减少,之后又略微增加。由此推测RXLR115890在辣椒疫霉病的发病前期起着关键作用,为研究辣椒疫霉菌效应子RXLR115890的功能提供了有利的参考。
1.2 CRN效应子
植物细胞程序性死亡(programmed cell death,PCD)是植物抵抗病原物入侵的防卫机制,病原物通过延缓或抑制植物PCD的发生来实现自身的入侵和定殖[11]。CRN (crinkling and necros-inducing protein)蛋白是卵菌如辣椒疫霉菌特有的一类效应子,研究表明,多数CRN效应子在很大程度上会进入植物细胞核并阻止植物细胞的PCD反应,以协助辣椒疫霉菌的侵染[12]。陈孝仁等[13]研究发现,辣椒疫霉菌CRN效应子Pc506611能够抑制本氏烟的PCD,之后又利用序列分析和RT-PCR技术发现了3个在病原菌生长发育阶段表达上调的基因,分别是Pc559084、Pc570403和Pc20879;经过克隆和测序后发现其编码的蛋白产物均含有HVLVVVP和LFLAK基序;在本氏烟上的基因瞬时表达结果表明,编码基因Pc570403和Pc559084能够抑制植物PCD,并且Pc559084能够促进辣椒疫霉菌的侵染[14]。因此,利用效应子如CRN等来阻断植物细胞的程序性死亡或许是辣椒疫霉菌的作用机制之一。这些试验结论使人们更进一步了解CRN效应子,同时也为更深层次地了解植物与疫霉菌互作的机制提供了有利的理论依据。
1.3 elicitins效应子
疫霉菌可分泌一种被称为elicitins的胞外蛋白激发子并诱导一些植物产生超敏反应[15],进而使植物产生抗性反应。邹金城[16]等经过转录分析、系统发育分析等技术操作后,获得了2个elicitins基因,分别是Pc508770和Pc508763,二者可刺激本氏烟产生HR;还获得了在辣椒疫霉病发病时期表达上调的6个elicitins基因,值得一提的是它们在病原菌不同的侵染阶段差异上调表达,如Pc11949在整个感染过程中上调表达,而Pc127268只在侵染后期上调表达。辣椒疫霉菌elicitins有多种生物学功能,对该基因的研究为了解辣椒疫霉的分子致病机制提供了重要数据,也为综合治理辣椒疫病提供了科学依据。
1.4 MAPK基因
MAPK(mitogen-activated protein kinase,MAPK)信号途径在细胞信号转导网络中处于重要地位,参与多种生理生化反应,调控着细胞的生长、分化,在辣椒疫霉侵染宿主的过程中也不可或缺。朱彤彤等[17]利用生物信息学技术,从基因组库中筛选出了十几个MAPK基因,它们大多数位于基因组负链中。这些基因所编码的蛋白产物各不相同,最终长度在200~1 000个氨基酸之间。进化关系分析表明,这些基因与真菌保持相对独立,但在疫霉属间相对保守,说明MAPK在疫霉菌的信号调控网络中仍起重要作用。通过对辣椒疫霉菌MAPK基因的分析,为进一步了解该基因家族提供了有利的参考。
2 辣椒疫霉的分子生物学检测
传统检测植物病害的方法为观察植株的发病部位症状,并进行形态学鉴定和分离纯化致病菌,鉴定过程相对耗时,再加上辣椒疫霉菌引起的病害与腐霉菌等引起的病害症状相似,二者难以分辨,给防治工作造成一定程度的困难[18]。这些年随着科学技术的进步,病原菌的检测技术已深入到分子水平。
聚合酶链式反应(PCR)是一种快速灵敏的分子检测技术,最常见的方法是利用核糖体转录间隔区域(ITS)序列的不同来进行鉴定,然而,这种方法很难区别一些具有相似ITS序列的分类地位很近的物种。李毛毛等[19]通过分析对比疫霉属的多种Ypt1基因序列,设计出了一对能检测辣椒疫霉的特异性引物PcYpt1F/PcYpt1R,该体系可检测至少10 pg含量的基因组,并且灵敏度高,可用于快速检测含有辣椒疫霉菌的样品。
程颖超等[20]根据辣椒疫霉菌和尖孢镰刀菌等近10种常见传播方式类似的病原菌基因序列的差异,设计出了1对辣椒疫霉菌的特异性引物YM2F/YM2R,并依此建立了一套實时定量PCR检测体系,试验证明该体系具有良好的线性关系,灵敏度远高于普通PCR,为1×10-1 pg·μL-1。他们对土壤样品的检测试验表明,在一定范围内病害的发生以及流行程度都与病原微生物的密度成正比。这些试验结果可以为辣椒疫病的监测以及早期防治提供有利的理论依据。
3 综合治理
3.1 选用抗病品种
孙文秀等[21]、王丽慧等[22]研究表明,大部分辣椒疫霉菌菌株分化与地理位置没有直接关系,因此,选用抗病的辣椒品种显得至关重要。除了常规育种方法外,还可以采用分子育种等手段快速创造新的辣椒抗病品种。
3.2 农业防治措施
3.2.1 间作 试验证明,辣椒和玉米间作在一定程度上可以控制辣椒疫霉菌在作物之间的传播,作用机制主要是玉米发达的根系能够吸附致病菌的游动孢子,同时会分泌一些化学物质以抑制孢子的萌发,甚至裂解疫霉菌产生的孢子,从而有效控制辣椒疫病[23]。另外,江冰冰等[24]的试验证明,利用辣椒和韭菜间作在一定程度上也可以防治辣椒疫病,在该体系中,可以每2行辣椒之间种植2~3行韭菜,以控制辣椒疫病的传播。这种间作模式可显著抑制辣椒疫病的扩散,其作用机制可能是韭菜根系分泌的某种化学物质降低了辣椒疫霉孢子的传播速率。
3.2.2 土壤强还原 辣椒疫病也是一种土传病害,因为辣椒疫霉菌可存在于土壤中,当条件适宜时能够萌发继而侵染植物[25]。因此,杀死土壤中的病原微生物在预防和控制疫病中显得特别重要。早在2000年,人们就发明了土壤强还原法(Reductive soil disinfestation,RSD),以减少土壤中病原微生物的数量。RSD的关键环节是在土壤中加入易分解的有机物以及保证土壤的厌氧状态,有机物在厌氧状态下的分解产物能对病原菌产生毒害作用,以达到有效杀灭土传病原菌的目的。黄新琦等[26]的研究结果表明,当土壤中的有机碳源主要来源于玉米秸秆时,土壤强还原的过程中主要产生了丁酸等物质;用50 mmol·L-1丁酸溶液处理的土壤中辣椒疫霉菌的数量为对照的38.9%,表明丁酸在整个还原过程中扮演着重要角色。
3.2.3 科学管理 高温高湿会加剧辣椒疫病的发生,因此合理灌水是预防疫病发生的关键措施,避免漫灌,要做好田间排水并随时留意田间湿度,防止田间积水。对于已经发病的植株,应进行集中清理,不能随意堆放,以避免病害的传播。王光飞等[27]发现,生物炭用量在1.33%时各项指标如群落结构、土壤酶活性等稳定在最佳值或者保持在其范围内,有利于增强土壤的抑病性。
3.3 化学防治
何烈干等[28]通过试验发现,68%精甲霜·锰锌水分散粒剂和50%烯酰吗啉可湿性粉剂可有效防治辣椒疫病,并且在农业生产中可采用多种不同的杀菌剂轮换使用的方式,这样可在有效治理辣椒疫病的同时避免抗药性的产生。
薛春生等[29]认为,好普(2.0%氨基寡糖素水剂)和肃克(碘与寡糖衍生物的络合物,是20%寡聚酸碘水剂)是防治辣椒疫病的有效生物制剂,其中肃克防治效果最高可达80%,略高于好普。
曹云等[30]认为,DMPP与碳酸氢铵的结合可以减少土壤中氨氧化细菌的数量,从而降低硝态氮含量,增加铵态氮含量,使泥土的pH值增大,进而使土壤中辣椒疫霉菌的数目下降,从而能有效抑制辣椒疫病的发生。
另有研究发现,肉桂精油[31]、山苍子精油[32]等植物精油,金丝草的乙酸乙酯萃取物[33],黄帚橐吾的提取物[34]等对辣椒疫霉菌均有抑制作用,因此这些物质对防治辣椒疫病有一定的应用潜力。
3.4 生物防治
陈林等[35]从辣椒根的韧皮部中分离出对辣椒疫霉菌拮抗作用显著的辣椒植株内生菌株,即Peanibacillus polymyxa sp.,并且它还可固定大气中的氮。张娜等[36]将含有地衣芽胞杆菌chiMY基因的重组质粒pDM转化到具有较高杀虫活性的Bt519-1菌株,结果证明,工程菌对辣椒疫霉菌的抑制效率在90%以上,即Bt519(pDM)是一株潜在的生防菌株。杨宇红等[37]研究表明,hrp-菌株与BABA具有协同作用,将二者按比例混合使用对辣椒疫病有一定程度的防治作用,其中以0.1%BABA、hrp-菌株1×106 CFU·mL-1配比效果最佳。谈泰猛等[38]从健康辣椒植株的根际土壤中得到一株枯草芽孢杆菌IBFCBF-4,对辣椒疫病具有显著的拮抗作用。盆栽试验结果表明,该菌对辣椒疫病防治及促生效果显著。刘青等[39]分离得到绿色木霉Tv-1、Tv2,钩状木霉Tha-1、哈茨木霉Thz-2等4种菌株对辣椒疫霉菌的抑制率均在90%以上。这些高效菌株主要是通过降低致病因子酶活力、破坏细胞壁结构等方式起拮抗作用。杨定祥等[40]从数百种细菌中筛选出数十株可抑制辣椒疫霉菌的细菌菌株,其中以来自珊瑚的解淀粉芽孢杆菌(Bacillus amyloliquefaciens) SH-27菌株的抑菌效果最显著,室内盆栽试验表明,经过该菌株发酵液处理后的各项生理指标均显著优于对照,即海洋细菌SH-27菌株不仅对辣椒有促生长效果,而且对辣椒疫霉菌具有拮抗作用,具有作為生物防治菌株的潜力。张艳萍等[41]在研究马铃薯晚疫病菌的拮抗菌株时发现,从土壤中分离出的假单胞菌(Pseudomonas sp.)HC5对辣椒疫霉菌也有一定的抑制作用,而且还能产生氢氰酸和硝吡咯菌素这两种抗生素,所以该菌株也具有作为生物防治菌株的潜力。
生物防治由于具有高效且无毒无害等特点愈发受到人们关注,这些生防菌的发现为辣椒疫病的生物防治提供了理论支撑和技术准备。
4 展 望
随着科学技术的进步,人们对辣椒疫病发病分子机制的研究也愈发深入,对于辣椒疫病的了解也更加透彻。但是,仍然有一部分的研究暂时还处于试验室研究阶段,并未真正适用于大田试验;有的虽然应用于实际生产中,但尚未大范围推广。在辣椒疫病的防治措施中,化学试剂对环境都有一定的污染,而微生物及其产物就会在疫病防治方面发挥重要作用。预防比治理更重要,如果想最大程度减少辣椒疫病带来的损失,关键还是要选育抗病品种。因此,接下来要做的就是针对辣椒疫病的发病机制,如致病基因等来培育新的有效的抗病品种,并将其应用于生产实践中。
参考文献
[1] 易图永,谢丙炎,张宝玺,等.辣椒疫病防治研究进展[J].中国蔬菜,2002(5):52-55.
[2] JONES J,DANGL J.The plant immune system[J].Nature,2006,444(7117):323-329.
[3] 李秀奇,冯锐,黄振岭,等.辣椒疫霉效应分子RxLR22034的表达纯化及晶体生长[J].山东农业大学学报(自然科学版),2018,49(3):383-387.
[4] BIRCH P,ARMSTRONG M,BOS J,et al.Towards understanding the virulence functions of RXLR effectors of the oomycete plant pathogen Phytophthora infestans[J].Journal of Experimental Botany,2009,60(4):1133-1140.
[5] WHISSON S C,BOEVINK P C,MOLELEKI L,et al.A translocation signal for delivery of Oomycete effector proteins into host plant cells[J].Nature,2007,450(7166):115-118.
[6] WIN J,MORGAN W,BOS J,et al.Adaptive evolution has targeted the C-terminal domain of the RXLR effectors of plant pathogenic Oomycetes[J].The Plant Cell,2007,19(8):2349-2369.
[7] 杨楠,张丽,丁鹏,等.辣椒疫霉效应分子RxLR121504功能特性的研究[J].菌物学报,2017,36(9):1233-1242.
[8] 陈孝仁,张烨,黄沈鑫,等.辣椒疫霉RXLR型效应子PcAvh2的序列多态性、基因转录特征及功能[J].微生物学报,2018,58(12):2204-2215.
[9] BOS J,ARMSTRONG M,GILROY E.Phytophthora infestans effector AVR3 aisessential for virulence and manipulates plant immunity by stabilizing host E3 ligase CMPG1[J].Proceedings of the National Academy of Sciences of the United States of America,2010,107(21):9909-9914.
[10] 张丽,王俪颖,梁涛,等.辣椒疫霉效应因子RxLR115890不同侵染时期的差异性表达[J].山东农业大学学报(自然科学版),2018,49(3):388-392.
[11] CLEM R J.Baculoviruses and apoptosis:adiversity of genes and responses[J].Current Drug Targets,2007,8:1069-1074.
[12] STAM R,HOWDEN A J,DELGADO-CEREZO M,et al.Characterization of cell death inducing Phytophthora capsici CRN effectors suggests diverse activities in the host nucleus[J].Frontiers in Plant Science,2013,4387.
[13] CHEN X,XING Y,LI Y,et al.RNA-Seq reveals infection-related gene expression changes in Phytophthora capsici[J].Plos One,2013,8(9):e74588.
[14] 陈孝仁,李艳朋,邢玉平,等.辣椒疫霉CRN编码基因的克隆、转录特征及在与寄主互作中的作用[J].植物病理学报,2015,45(4):384-394.
[15] JIANG R H Y,TYLER B M,WHISSON S C,et al.Ancient origin of elicitin gene clusters in Phytophthora genomes[J].Molecular Biology and Evolution,2006,23(2):338-351.
[16] 邹金城,邢玉平,孙华伟,等.辣椒疫霉elicitins基因的克隆及瞬时表达分析[J].南京农业大学学报,2014,37(6):28-36.
[17] 朱彤彤,杨磊,孙文秀.辣椒疫霉MAPK基因家族的鉴定及生物信息学分析[J].生物技术,2018,28(5):460-466.
[18] 罗加凤,刘跃庭,廖芳,等.进境美国加州脐橙中丁香疫霉Phytophthora syringae的截获[J].菌物学报,2012,31(1):24-30.
[19] 李毛毛,赵伟,汪涛,等.以Ypt1基因序列为靶标的辣椒疫病菌快速分子检测[J].植物病理学报,2014,44(5):546-551.
[20] 程颖超,康华军,石延霞,等.辣椒疫霉菌RT-PCR检测技术的建立及应用[J].园艺学报,2018,45(5):997-1006.
[21] 孙文秀,张修国,贾永健,等.不同地区辣椒疫霉菌遗传多样性的RAPD分析[J].植物病理学报,2005,35(4):340-344.
[22] 王丽慧,高洁铭,陶丽婷,等.青海省辣椒疫霉菌SSR标记的遗传多样性分析[J].分子植物育种,2018,16(21):7068-7076.
[23] YANG M,ZHANG Y,QI L,et al.Plant-plant-microbe mechanisms involved in soil-borne disease suppression onamaize and pepper inter-cropping system[J].PLos One,2014,9(12):e115052.
[24] 江冰冰,张彧,郭存武,等.韭菜和辣椒间作对辣椒疫病的防治效果及其化感机制[J].植物保护学报,2017,44(1):145-151.
[25] 李世东,缪作清,高卫东.我国农林园艺作物土传病害发生和防治现状及对策分析[J].中国生物防治學报,2011,27(4):433-440.
[26] 黄新琦,温腾,孟磊,等.土壤强还原过程产生的有机酸对土传病原菌的抑制作用[J].植物保护,2015,41(6):38-43.
[27] 王光飞,马艳,郭德杰,等.不同用量秸秆生物炭对辣椒疫病防控效果及土壤性状的影响[J].土壤学报,2017,54(1):204-215.
[28] 何烈干,马辉刚,陈学军,等.辣椒种质疫病抗性鉴定及防治药剂的筛选[J].江西农业大学学报,2014,36(5):1054-1060.
[29] 薛春生,何瑞玒,肖淑芹,等.辽宁省辣椒疫霉菌生理小种的生物学特性及生物制剂对辣椒疫病的防效[J].植物保护学报,2017,44(4):650-656.
[30] 曹云,王光飞,郭德杰,等.DMPP增强碳酸氢铵防控辣椒疫病的效果与机制[J].中国生态农业学报,2016,24(10):1382-1390.
[31] 王海娇,王轶楠,高飞,等.肉桂精油对辣椒疫霉的生物活性[J].中國生物防治学报,2018,34(3):341-468.
[32] 王轶楠,赵特,高飞,等.山苍子精油对辣椒疫霉生长发育的影响及对辣椒疫病的防效[J].植物保护学报,2018,45(5):1112-1120.
[33] 雷绍南,倪林,吴晓芹,等.金丝草抑菌活性成分研究[J].农药药学学报,2017,19(1):119-124.
[34] 李晶,李娜,丁品,等.黄帚橐吾提取物对保护地辣椒4种病原真菌的抑制活性及其病害防效[J].草业学报,2018,27(4):56-68.
[35] 陈林,董兆麟,段康民.一株具有抗菌作用的P. polymyxa sp.菌株的分离鉴定[J].西北大学学报(自然科学版),2011,41(6):1010-1013.
[36] 张娜,李蓬飞,陈月华,等.转异源chi基因的Bt工程菌株的生防潜力评价[J].微生物学通报,2016,43(1):88-96.
[37] 杨宇红,茆振川,凌键,等.β-氨基丁酸和茄青枯菌hrp-突变体协同防治辣椒疫病研究[J].中国生物防治学报,2017,33(4):519-524.
[38] 谈泰猛,黎继烈,申爱荣,等.辣椒疫病拮抗菌的分离、鉴定及其生防效果[J].生态学杂志,2017,36(4):988-994.
[39] 刘青,李升,梁才康,等.贵州地区木霉菌分离鉴定及对辣椒疫霉的拮抗作用[J].微生物学通报,2019,46(4):741-751.
[40] 杨定祥,林巧玲,卢乃会,等.拮抗辣椒疫霉菌海洋细菌菌株SH-27的筛选鉴定及其防病促生作用[J].微生物学通报,2018,45(1):54-63.
[41] 张艳萍,令利军,赵瑛,等.土壤中细菌HC5的分离鉴定及抑菌活性测定[J].农业资源与环境学报,2018,35(6):527-532.
猜你喜欢
华夏医学(2016年4期)2016-12-12
中国市场(2016年36期)2016-10-19
科技视界(2016年21期)2016-10-17
科学与财富(2016年28期)2016-10-14
企业导报(2016年12期)2016-06-17
