
天然和人工固沙灌木林蜘蛛和甲虫分布与环境因子的关系
2020-11-24 09:30刘继亮赵文智李锋瑞巴义彬
生态学报 2020年21期
刘继亮,赵文智,*,李锋瑞,巴义彬
1中国科学院西北生态环境资源研究院,临泽内陆河流域研究站, 兰州 730000 2 河北大学博物馆, 保定 071002
人工绿洲是我国内陆河流域主要的自然景观之一,它的面积所占比重很小,确是人类活动的主要场所[1-2]。绿洲外围乔木林带的建立能有效阻止风沙侵害,但也需要大量的水资源来维持,而灌木林带与乔木林带相比水资源消耗较低,它主要依赖自然降雨来维持。因而,近年来绿洲外围营建了大面积的人工固沙灌木林,主要栽植耐旱的灌木梭梭和柽柳,它们在荒漠绿洲过渡区呈斑块状分布。人工栽植的灌木林与天然灌木林相比,植被盖度较高、种类单一,草本的盖度及物种丰富度也较低,土壤结构、养分及含盐量等也发生变化,这也会影响地表及土壤节肢动物群落结构及生态功能[3- 5]。地表蜘蛛和甲虫是干旱区天然和人工固沙林地两种主要的地表节肢动物类群,它们既是荒漠生态系统重要的消费者又是重要的捕食者,同时也是特殊的分解者,对环境变化响应十分敏感[6-9]。然而,我们对干旱区固沙灌木植被恢复过程中植被、土壤环境变化如何影响地表节肢动物群落营养结构及功能的认识还很有限,不清楚地表节肢动物群落中关键类群-蜘蛛和甲虫群落(荒漠食物网的重要组分)对植被和土壤环境变化的响应规律。
干旱区、半干旱区植被恢复改变地表生态水文环境,强烈影响飞行昆虫、地表及土栖节肢动物多样性[10-12]。对地上昆虫而言,植被恢复最主要的改变了植被群落结构从而影响了昆虫多样性[13]。天然植被转变为人工植被随着植被结构简单化,昆虫多样性也随之降低[14]。地表节肢动物(主要由蜘蛛和昆虫组成)与飞行昆虫不同,它们的变化既受植被变化的影响,同时也受植被驱动的土壤环境变化的影响,土壤环境改变影响地表节肢动物的栖居和繁殖环境。李岳诚等[15]在宁夏白芨滩的研究发现,土壤含水量、植被盖度和人为干扰强度是造成不同生境甲虫多样性差异的主要因素。娄巧哲等[16]在古尔班通沙漠南缘的研究发现,荒漠地表甲虫群落的物种丰富度、多样性指数、均匀度指数、优势度指数与植被盖度、土壤含水量及养分均存在显著线性相关,而甲虫的活动密度与土壤含水量呈负相关关系。由此可见,人工植被恢复过程中植被和土壤环境是影响地表及土壤节肢动物群落变化的关键环境因子,但因地域环境差异,不同动物类群对环境变化的响应模式不同,从而改变了以节肢动物为主的碎屑食物网的结构及功能。鉴于此,本文通过河西走廊中部荒漠-绿洲过渡区天然灌木林和人工梭梭、柽柳灌木林地表蜘蛛和甲虫群落组成与植被和土壤环境的关系定位监测,探讨人工固沙灌木及灌木种类差别对蜘蛛和甲虫群落结构及多样性的影响,确定植被和土壤环境要素对蜘蛛和甲虫分布的影响及相对贡献率,这有助于理解和认识人工固沙植被恢复对以碎屑或植物为基础的荒漠生态系统食物网结构及功能的影响机制。
1 材料与方法
1.1 研究区概况
研究区属于大陆性干旱气候,冬季漫长寒冷,春季干旱少雨,降雨主要集中在7、8、9三个月,夏季和秋季比较湿润。年平均降水量117.0 mm(主要集中在7—9月),年平均蒸发量2390 mm,年平均气温7.6℃,≥10℃年积温3085℃,无霜期165天。年均日照时数为3045 h,太阳辐射总量为611272.8 J cm-2a-1。本区冬季盛行西北风,年均风速3.2 m/s,最大风速可达21.3 m/s,≥8级以上大风年均15天左右,且集中于3—5月。
1.2 试验设计及样品采集
本研究区位于黑河中游张掖绿洲外围的荒漠-绿洲过渡区(39.37°—39.40°N,100.14—100.17°E)。张掖绿洲外围荒漠绿洲过渡区人工固沙植被区栽植的灌木主要有梭梭Haloxylonammodendron(C. A. Mey.) Bunge和柽柳TamarixchinensisLour.,梭梭主要栽植在沙丘上,而柽柳主要栽植在丘间相对低洼、平坦的区域,它们均呈斑块状分布。人工栽植的梭梭、柽柳林密度较高,随着灌木生长,部分灌木枯死,林地开始疏化。天然灌木林主要灌木种为沙拐枣CalligonummongolicumTurcz.、泡泡刺NitrariasphaerocarpaMaxim.和小果白刺NitrariasibiricaPall.,丘间灌木盖度较高,而沙丘上灌木盖度较低。
我们依据灌木栽植年限在荒漠绿洲过渡区选择了20—25年的人工梭梭林和柽柳林,每种人工灌木林我们选取9个相对独立的灌丛斑块,灌丛斑块之间相距1000—2000 m,每个灌丛斑块选取30 m×50 m的区域作为蜘蛛和甲虫样品采集区。天然灌木林选在人工固沙植被毗邻的丘间低地区,随机选取9个相对独立地点,每个地点选取30 m×50 m的区域作为蜘蛛和甲虫样品采集区。每个动物样品采集区布设10个陷阱收集器(其中5个位于灌丛下,5个位于灌丛间),陷阱之间距离≥10 m。2012年干季(5月下旬)和湿季(8月中旬)利用陷阱法对地表活动的蜘蛛和甲虫种类和数量进行系统调查,两个调查期动物样品采集时间均为7天。捕获到的蜘蛛和甲虫样品带回实验室,采用体视显微镜和生物显微镜进行种类鉴定和数量统计,蜘蛛样品参照分类资料鉴定至科,按科统计数量(部分蜘蛛为幼体无法准确鉴定至种,因而蜘蛛样品鉴定至科),甲虫样品参照分类资料鉴定至属或种,按照属或种统计数量[17- 20]。
采用样方法调查动物采样区植物群落组成,每个动物样品采集区随机选取1个10 m×10 m和5个1 m×1 m的灌木和草本样方,估算灌木盖度、草本物种数,利用烘干法测定草本地上生物量。随机选取3—5株灌木测定灌丛下和灌丛间凋落物的现存量。每个动物样品采集区随机选取3株灌木,测定灌丛下凋落物的现存量。5个采集5个0—20cm土壤样品混合后带回室内使用干筛法测定土壤含砂量(>0.05 mm),使用pH计和电导率仪测定土壤pH和电导率,使用重铬酸钾氧化—分光光度法测定有机碳含量,半微量开氏法测定全氮含量。植被和土壤环境特征见表1和表2。

表1 天然灌木林、人工梭梭和柽柳林植被环境特征
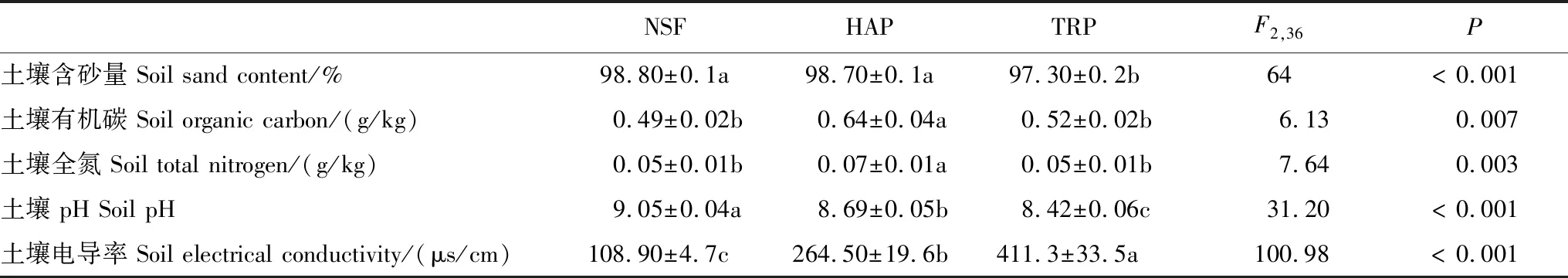
表2 天然灌木林、人工梭梭和柽柳林土壤环境特征
1.3 数据处理
首先将2个采样期每个动物样品采集区10个陷阱收集器合并统计,统计蜘蛛和甲虫的活动密度、类群丰富度和多样性指数(Shannon-Wiener)。采用单因素方差分析(GLM模型)比较蜘蛛和甲虫活动密度、类群丰富度和多样性指数的差异。同时,分析3种生境植被和土壤环境因子的差异。多样性指数计算使用BioDiversity Pro软件包[21],方差分析使用SPSS 21.0软件包。
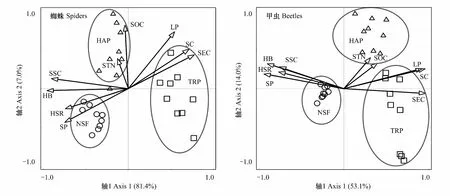
利用SIMPER相似性分析3种生境间物种组成的相似性和相异性,然后利用非参数的相似性检验(Analysis of similarities,ANOSIM)和非线性多维标定排序(Non-metric multidimensional scaling,NMDS)比较天然和人工固沙植被蜘蛛和甲虫群落差异[22]。ANOSIM用来检验不同群落间差异的显著性,R值通常为0—1,表示组内和组间的差异程度[22]。 NMDS排序结果采用胁强系数(Stress)衡量MDS分析结果的优劣,其中Stress<0.01,完全可信;0.01 采用RDA(冗余度分析,Redundancy analysis)和CCA(典范对应分析,Canonical correlation analysis)分析植被与土壤生态环境因子对蜘蛛和甲虫群落影响的贡献率。具体分析方法是:先采用DCA排序分析蜘蛛和甲虫群落确定第1轴梯度值(Gradient)。蜘蛛群落DCA排序的梯度值为1.4,选用RDA排序;甲虫群落DCA排序的梯度值为3.1,选用CCA排序。利用逐步向前选择(Interactive-forward-selection)确定9个植被和土壤环境因子的相对贡献量,利用蒙特卡洛置换检验(Monte-Carlo permutation test 999)。排序分析使用蜘蛛和甲虫个体数量的数据,分析前所有数据进行log (x+1)转换,统计分析使用通用的排序软件CANOCO 5.0[23]。利用Pearson相关分析主要蜘蛛和甲虫类群与排序确定重要植被和土壤环境因子的相关性,相关分析使用SPSS 21.0软件包。 天然灌木林与人工梭梭和柽柳林蜘蛛活动密度和多样性指数差异显著(F2,36=91.03,P<0.001;F2,36=20.02,P<0.001),而蜘蛛类群丰富度差异不显著(F2,36=2.71,P=0.087)。人工梭梭和柽柳林蜘蛛活动密度均显著高于天然灌木林,其活动密度约是天然灌木林的1.7倍和4.6倍(图1)。蜘蛛多样性指数与活动密度变化相反,天然灌木林蜘蛛的多样性指数显著高于人工梭梭和柽柳林(图1)。此外,研究还发现人工柽柳林蜘蛛的活动密度还显著高于人工梭梭林,蜘蛛多样性指数变化刚好相反(图1)。 天然灌木林与人工梭梭和柽柳林甲虫活动密度、类群丰富度和多样性指数均明显不同(F2,36=266.75,P<0.001;F2,36=12.77,P<0.001;F2,36=66.19,P<0.001)。天然灌木林甲虫的活动密度显著高于人工梭梭和柽柳林,而多样性指数变化相反(图1)。天然灌木林甲虫的活动密度约是人工梭梭和柽柳林的18.3倍和13.4倍(图1)。人工柽柳和梭梭林甲虫的多样性指数显著高于天然灌木林,而人工梭梭林甲虫类群丰富度显著高于天然灌木林和人工柽柳林(图1)。 图1 天然灌木林、人工梭梭和柽柳林蜘蛛和甲虫活动密度、类群丰富度和多样性指数比较Fig.1 The activity density, taxa richness and Shannon-Wiener diversity index of ground spiders and beetles among natural shrub forest (NSF), Haloxylon ammodendron (HAP) and Tamarix ramosissma plantation (TRP)字母不同代表不同生境间存在显著差异(P<0.05) 蜘蛛和甲虫群落的NMDS排序结果较好的反映了天然灌木林和人工梭梭、柽柳林间蜘蛛和甲虫群落相似性,天然灌木林和人工梭梭、柽柳林蜘蛛群落差异较小,而不同生境间甲虫群落相差较大(图2)。ANOSIM检验表明,天然灌木林和人工梭梭、柽柳林蜘蛛和甲虫群落结构存在显著差异(GlobalR=0.79,P<0.001;GlobalR=0.93,P<0.001),三种生境之间蜘蛛和甲虫群落组成不同。 图2 天然灌木林、人工梭梭和柽柳林蜘蛛和甲虫群落NMDS排序图Fig.2 NMDS plots indicating 2-dimensional distances on ground spiders and beetles among natural shrub forest (NSF), Haloxylon ammodendron (HAP) and Tamarix ramosissma plantation (TRP) SIMPER分析发现,天然灌木林和人工梭梭林蜘蛛和甲虫群落的平均相异性为28.0% 和38.4%。天然灌木林和人工梭梭林间蜘蛛群落差异的主要贡献类群为球蛛科(贡献率 32.7%)、跳蛛科(16.9%)、微蛛亚科(15.5%)和狼蛛科(14.8%);甲虫群落差异的主要贡献类群为东鳖甲属(12.8%)、戈壁琵甲(10.4%)、谢氏宽漠王(10.0%)。天然灌木林和人工柽柳林蜘蛛和甲虫群落的平均相异性为34.8% 和52.5%。天然固沙灌木林和人工柽柳林间蜘蛛群落差异的主要贡献类群为狼蛛科(35.0%)、球蛛科(23.7%)、跳蛛科(12.8%)和微蛛亚科(12.2%);甲虫群落的主要贡献类群为东鳖甲属(10.5%)。两种人工固沙植被蜘蛛和甲虫群落组成也存在一定差异,人工梭梭和柽柳林蜘蛛和甲虫群落的平均相异性为26.4%和32.9%。人工梭梭和柽柳林间蜘蛛群落差异的主要贡献类群为狼蛛科(44.1%)、微蛛亚科(12.5%)、皿蛛亚科(12.1%)和球蛛科(10.2%);甲虫群落的主要贡献类群为叩甲科(10.0%)。 天然灌木林和人工梭梭、柽柳林蜘蛛与环境因子的排序结果表明,9个植被和土壤环境因子解释了85.3%的蜘蛛群落变异(F=17.8,P=0.001)。pRDA分析发现,灌木盖度、草本生物量、凋落物量、土壤砂含量和电导率是影响蜘蛛分布的主要环境因子,它们解释了82.1%的蜘蛛群落变异(表3)。第1轴反映了天然灌木林、人工梭梭林地表蜘蛛群落组成与人工柽柳林不同,而第2轴反映了天然灌木林与人工梭梭林地表蜘蛛群落组成存在一定差异(图3)。不同类型蜘蛛与主要环境因子的关系不同,结网型的球蛛科与灌木盖度、凋落物量和土壤电导率呈显著负相关(P<0.05),而与草本生物量呈显著正相关(P<0.05);结网型的个体较小的皿蛛亚科与电导率呈显著正相关(P<0.05),而与草本生物量和土壤砂含量呈显著负相关(P<0.05)。游猎型蜘蛛狼蛛科、平腹蛛科和蟹蛛科与灌木盖度、凋落物量和土壤电导率呈显著正相关(P<0.05),而与草本生物量和土壤砂含量呈显著负相关(P<0.05);游猎型逍遥蛛科仅与凋落物量呈显著负相关(P<0.05)。伏击型蜘蛛跳蛛科和个体较小结网型蜘蛛微蛛亚科受5个环境因子的影响较小(P>0.05)。 表3 pRDA分析确定9个植被和土壤环境因子对蜘蛛分布的相对贡献率 图3 天然灌木林和人工梭梭、柽柳林蜘蛛和甲虫与主要环境变量关系的RDA和CCA二维排序图Fig.3 The RDA and CCA two-dimensional ordination diagram of ground spiders and beetles with explanatory variables among natural shrub forest (NSF), Haloxylon ammodendron (HAP) and Tamarix ramosissma plantation (TRP)SC:灌木盖度 Shrub cover;HB:草本生物量 Herbaceous biomass; HSR:草本物种数 Herbaceous species richness;LP:凋落物量 Litter production;SSC:土壤含砂量 Soil sand content;SOC:土壤有机碳 Soil organic carbon;STN:土壤全氮Soil total nitrogen; SEC:土壤电导率 Soil electrical conductivity;SP:土壤pH Soil pH 天然灌木林和人工梭梭、柽柳林甲虫与环境因子的排序结果表明,9个植被和土壤环境因子解释了66.5%的甲虫群落变异(F=6.70,P=0.001)。pCCA分析发现,灌木盖度、草本生物量、土壤pH和含砂量是影响甲虫分布的主要环境因子,它们解释了60.6%的甲虫群落变异(表4)。第1轴反映了天然灌木林与人工梭梭林、柽柳林地表甲虫群落组成不同,而第2轴反映了人工梭梭林与柽柳林地表甲虫群落组成存在一定差异(图3)。9个优势甲虫种与主要环境因子的相关分析结果表明,戈壁琵甲和阿小鳖甲均与4个因子的相关性较小(P>0.05),克氏扁漠甲、谢氏宽漠王、东鳖甲属种1、东鳖甲属种2、齿足象属、甜菜象属甲虫均与灌木盖度呈显著负相关,与草本生物量、土壤砂含量和pH呈显著正相关(P<0.05)。婪步甲属种1甲虫与灌木盖度呈显著正相关,与草本生物量、土壤砂含量和pH呈显著负相关(P<0.05)。 表4 pCCA分析确定9个植被和土壤环境因子对甲虫分布的相对贡献率 干旱区人工固沙植被恢复主要表现为防风固沙营建的人工乔木林和灌木林,而绿洲外围天然和人工灌木林是绿洲外围主要的固沙植被。目前的研究表明,利用自然降雨的人工固沙灌木恢复大幅提高了植被盖度、改善了土壤环境,而随着灌木盖度增加也导致土壤盐碱环境发生变化,草本盖度趋于降低[24- 26]。天然固沙植被与人工固沙植被相比,植被和土壤环境均发生明显变化,地表和土栖土壤动物群落组成及多样性也发生明显改变。前期的研究发现,利用灌溉维持的人工林地及农田显著提高了土栖节肢动物的密度及多样性,而地表节肢动物密度呈降低的趋势,但多样性有所提高[3,4,27]。无灌溉的人工灌木林地与天然灌木林地地表和土壤节肢动物群落组成相差不大,地表节肢动物群落组成及多样性明显不同。人工灌木林地替代天然林地提高了地表节肢动物的多样性,降低了地表节肢动物的活动密度,这与刘继亮等[12,27]、Liu等[28]在宁夏北部沙地的研究结果相近。此外,研究还发现灌木种类不同对土壤环境恢复的影响也不同。梭梭和柽柳均是耐旱、泌盐灌木,柽柳林草本生物量、物种数及凋落物量均低于梭梭林,柽柳林土壤有机碳、全氮含量和pH也低于梭梭林,而土壤电导率显著高于梭梭林(表1),人工梭梭和柽柳林地表节肢动物群落组成也明显不同[4,12]。 研究还表明,高盖度无灌溉人工梭梭林和柽柳林与低盖度天然灌木相比,蜘蛛和甲虫两种优势地表节肢动物群落结构明显不同。人工梭梭和柽柳林蜘蛛的活动密度显著高于天然灌木林,而甲虫的密度显著低于天然灌木林。以上结果与郑国等[30]在云南和任海庆等[31]在海南热带雨林区的研究结果相反,而与姚润枝等[32]在北京退耕还林区、蒯国锋和刘新民[33]在宁夏沙坡头和Herrmann等[34]在以色列的研究结果接近。蒯国锋和刘新民[33]在宁夏沙坡头的研究也发现,无灌溉固沙人工林土壤蜘蛛群落的个体数、物种丰富度和物种多样性随固沙时间的延长而增加。以上结果表明,干旱或退化生境植被恢复明显改善了蜘蛛栖居环境和丰富了食物资源,从而提高了一些蜘蛛种群的数量及多样性,而湿润生境天然林转变为人工林地生境结构趋于简单、人为干扰增强,从而降低了一些蜘蛛的数量及多样性[31,33]。此外,不同灌木种类不同对蜘蛛群落变化的影响也不同,故增加人工固沙灌木种类有利于维持绿洲外围灌木林带地表蜘蛛的多样性[30]。 甲虫群落变化与蜘蛛不同,研究区人工梭梭、柽柳林与天然灌木林相比,降低了甲虫的活动密度,但提高了地表甲虫多样性,这与刘云慧等[35]在河北坝上地区和杭佳等[36]在宁夏黄土区的研究结果相反,而与李岳诚等[15]在宁夏白芨滩国家级自然保护区和娄巧哲等[16]在古尔班通古特沙漠南缘和的研究结果接近。研究区地表甲虫以拟步甲科为主,人工梭梭、柽柳林与天然灌木林相比,拟步甲科活动密度显著降低,而随着其他甲虫种类及活动密度的增加,地表甲虫多样性亦显著增加。此外,研究还发现人工柽柳林甲虫类群丰富度显著高于人工梭梭林,这可能与林下环境改善导致一些捕食性甲虫增加有关。拟步甲科、步甲科和象甲科是研究区主要地表甲虫类群,人工梭梭、柽柳林与天然灌木林相比,克氏扁漠甲、谢氏宽漠王、东鳖甲属、伪葬步甲属、半猛步甲属、齿足象属和甜菜象属甲虫活动密度均显著降低,这可能与土壤物理性质和草本多样性降低有关;而一角甲属、婪步甲属、暗步甲属和青步甲属甲虫在人工梭梭、柽柳林密度较高,尤其在人工柽柳林,这可能与灌木盖度增加改善了栖居条件和增加了食物资源有关[4,37]。 蜘蛛是干旱区食物网中最活跃和重要捕食者,其种类和密度变化关系到生态系统平衡和稳定[38-39]。 人工植被恢复过程中蜘蛛的种类、数量变化与植被和土壤塑造的生境条件密切相关。人工植被恢复通常导致植被类型单一,生境结构趋于简单,从而导致蜘蛛密度或多样性降低[30,40]。研究发现,在干旱区天然植被受土壤水分和养分限制植被盖度较低,人工固沙植被恢复显著提高了灌木盖度和凋落物量,而土壤砂含量降低及养分含量增加可能为一些蜘蛛类群的栖居提供了条件,这导致人工梭梭、柽柳林蜘蛛的活动密度显著高于天然灌木林。此外,研究还发现人工柽柳林蜘蛛的数量显著高于人工梭梭林,这与土壤盐碱及碳氮含量环境变化密切相关。不同捕食类型蜘蛛对天然灌木林、人工梭梭和柽柳林主要环境变化的响应模式也存在一定差异。结网型的球蛛科偏好在天然灌木林活动,个体较小的结网型蜘蛛和游猎型蜘蛛倾向在人工梭梭、柽柳林活动,伏击型蜘蛛和一些个体小的结网蜘蛛对植被变化的响应不敏感。以上研究结果与任海庆等[31]在海南热带雨林和人工橡胶林的研究结果接近。结网型蜘蛛偏好选择灌木盖度较低的生境,而游猎型蜘蛛更多选择在植盖度较高的人工生境活动,伏击型蜘蛛受植被盖度变化影响较小,不同蜘蛛类群的取食及繁殖导致它们对生境条件变化的响应模式不同。总之,天然灌木林、人工梭梭和柽柳林蜘蛛群落组成明显不同,天然灌木林与人工梭梭林相比提高了灌木盖度、凋落物量、土壤肥力和电导率较高,这导致二者蜘蛛群落组成不同;天然灌木林与人工柽柳林相比提高了灌木盖度、凋落物量土壤肥力和电导率,降低了草本生物量和土壤砂含量,这些因素导致二者蜘蛛群落组成不同。 甲虫是干旱区数量或生物量最多和最活跃的节肢动物类群之一,它既是捕食者又是重要的分解者,它是地表节肢动物群落中的关键类群。干旱区人工植被恢复引起的植被和土壤环境变化强烈影响地表甲虫数量及多样性。研究表明,人工固沙植被替代天然固沙植被,灌木盖度、草本生物量、土壤pH和砂含量是影响甲虫分布的主要环境因子。研究结果与李岳诚等[15]在宁夏白芨滩国家级自然保护区、娄巧哲等[16]在古尔班通古特沙漠、杭佳等[36]在宁夏黄土区和杨贵军等[41]在宁夏盐池关于地表甲虫的研究结果接近,地表植被和土壤环境变化影响甲虫分布及多样性。总之,天然灌木林与人工梭梭和柽柳林相比,灌木盖度、凋落物量及电导率增加,而草本生物量、物种丰富度、土壤砂含量和pH降低,这导致天然和人工固沙植被地表甲虫群落不同;两种人工固沙植被土壤养分含量不同导致人工梭梭和柽柳林地地表甲虫群落组成又略有不同。 不同甲虫类群对植被和土壤环境变化的响应模式不同决定了地表甲虫群落结构及多样性。研究发现灌木盖度、草本生物量、土壤砂含量和电导率与腐食性或植食性拟步甲科甲虫活动密度呈正相关,这与娄巧哲等[16]、吕昭智等[37]和杨贵军等[41]关于拟步甲科等地表甲虫分布的影响要素研究结果接近。植被盖度增加降低了鸟类等捕食者的捕食强度,这为一些个体大的拟步甲科甲虫提供了躲避天敌的小生境,而一些被鸟类等天敌捕食风险较小的甲虫趋于在开阔生境活动[42- 43];土壤机械组成变化是一些白天活动拟步甲科甲虫种重要限制因子,这些甲虫在土壤硬度较低和土壤疏松的生境能快速挖掘巢穴躲避天敌和抵御高温[37]。叶甲科、叩甲科和象甲科均是植食性甲虫,叶甲科和叩甲科与灌木盖度呈正相关,偏好先择在盖度较高的天然灌木林,而象甲科与灌木盖度呈负相关,偏好先择在盖度较低的天然灌木林,这可能食物资源的数量及质量有关。如象甲科甲虫主要是齿足象属和甜菜象属甲虫,齿足象属甲虫主要取食白刺属灌木叶片,甜菜象属甲虫主要取食一些荒漠草本(如沙芥属、雾冰藜和盐生草等),故象甲科甲虫主要在灌木盖度较低天然灌木林活动。 人工梭梭和柽柳林与天然灌木林相比,地表蜘蛛的活动密度提高,而地表甲虫活动密度降低;地表甲虫和蜘蛛多样性变化均与活动密度变化相反。两种人工固沙灌木林之间蜘蛛和甲虫群落结构也存在一定差异,人工柽柳林地表蜘蛛活动密度、多样性和甲虫物种丰富度均显著高于人工梭梭林。不同地表蜘蛛和甲虫类群对天然和人工灌木林的响应模式不同,人工灌木林与天然灌木林相比,球蛛科密度降低,而皿蛛亚科、狼蛛科和平腹蛛科均增加,人工柽柳林狼蛛科和平腹蛛科的密度又高于人工梭梭林;天然灌木林伪葬步甲、半猛步甲、齿足象、甜菜象和克氏扁漠甲、谢氏宽漠王和东鳖甲属甲虫均明显高于人工梭梭、柽柳林,婪步甲属、暗步甲属甲虫变化相反且在柽柳林的密度较高。 地表蜘蛛和甲虫群落变化与地表植被和土壤环境因子密切相关,9个植被和土壤环境因子解释了85.3%和66.5%的蜘蛛和甲虫群落变异。pRDA分析发现,灌木盖度、草本生物量、凋落物量、土壤砂含量和电导率是影响蜘蛛分布的主要环境因子,它们解释了82.1%的蜘蛛群落变异;pCCA分析发现,灌木盖度、草本生物量、土壤pH和含砂量是影响甲虫分布的主要环境因子,它们解释了60.6%的甲虫群落变异。总之,人工固沙植被转变为天然固沙植被过程中显著改变灌木盖度并驱动凋落物、草本生物量、土壤砂含量、pH及电导率变化是影响蜘蛛和甲虫群落变化的主要环境因子,它们互馈互作影响了蜘蛛和甲虫群落组成及多样性,进而改变荒漠生态系统碎屑食物网的结构和功能。 致谢:蛛形纲节肢动物样品鉴定得到了西南大学生命科学学院张志升教授和王露雨博士的帮助,特此致谢。2 结果与分析
2.1 天然灌木林、人工梭梭和柽柳林蜘蛛和甲虫群落比较
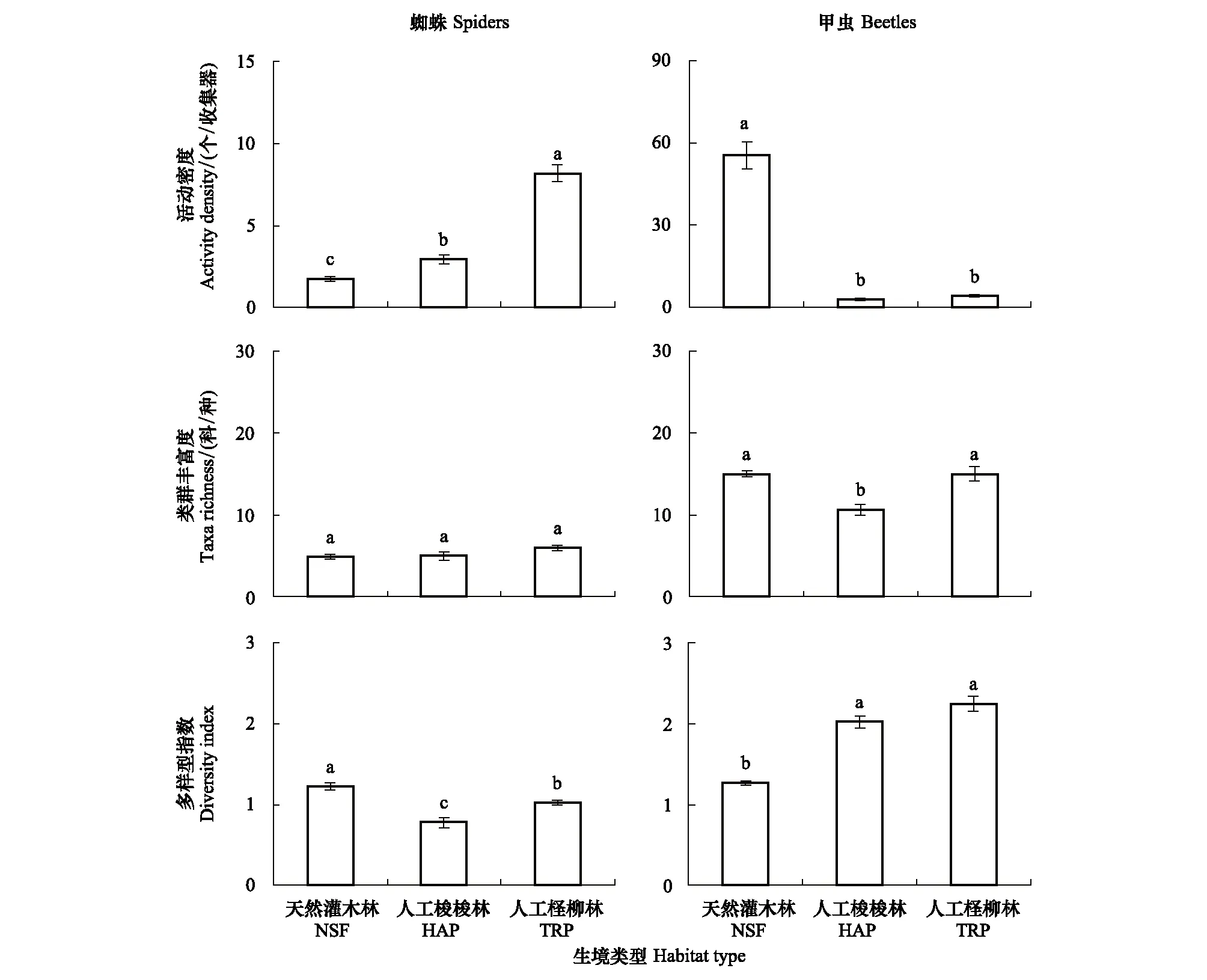
2.2 天然固沙植被、人工梭梭和柽柳林蜘蛛和甲虫群落相似性比较

3.3 天然灌木林、人工梭梭和柽柳林蜘蛛和甲虫分布与环境因子的关系


3 讨论
4 结论
猜你喜欢
哈哈画报(2022年4期)2022-04-19
农村科学实验(2021年21期)2021-12-25
科学大众(2020年17期)2020-10-27
河北林业科技(2020年3期)2020-03-23
山东林业科技(2019年6期)2020-01-02
农民致富之友(2019年31期)2019-11-23
中学生天地(B版)(2019年4期)2019-05-08
植物资源与环境学报(2019年1期)2019-04-02
农家科技下旬刊(2016年4期)2016-11-19
中国民族医药杂志(2016年8期)2016-05-09
