
闽江口芦苇与短叶茳芏空间扩展对湿地土壤磷赋存形态的影响
2020-11-24 09:29:42张鹏飞孙志高陈冰冰俞琳莺
生态学报 2020年21期
张鹏飞,孙志高,3,*,陈冰冰,何 涛,王 华,俞琳莺,李 晓,王 杰
1 福建师范大学湿润亚热带生态地理过程教育部重点实验室,福州 350007 2 福建师范大学地理研究所,福州 350007 3 福建师范大学福建省亚热带资源与环境重点实验室,福州 350007
磷是湿地中最为重要的限制元素之一[1],其含量及赋存形态的变化直接影响湿地生态系统的初级生产力。滨海湿地特别是河口湿地作为磷的重要源、汇或转化器,其对于河口湿地的稳定与健康可产生深刻影响[2]。河口湿地土壤中的磷养分供给状况对植物生长发育至关重要,特别是由于不同形态磷对植物的有效性存在较大差异,故其在土壤中的赋存状况直接影响着湿地植物的营养状况[3]。目前,国外关于河口湿地土壤磷的研究主要集中于不同磷形态或不同磷营养水平对湿地植物生态影响[4-5]、不同活性磷组分转化动态与生物可利用性[6]等方面。与之相比,国内关于河口湿地磷的相关研究虽已涉及辽河口湿地[7]、黄河口湿地[8]、珠江口湿地[9]、长江口湿地[10]和闽江口湿地,但研究内容主要侧重于湿地土壤全磷[7]、有效磷或无机磷含量的时空分布特征[11]、磷吸附-解吸及其影响因素[12]等方面,而对不同活性磷组分赋存的相关研究还不多见。
闽江是福建省最大的独流入海河流,闽江口湿地是闽江与东海相互作用形成的特殊生态类型,其对于维护区域生态安全发挥着重要作用。受河口水动力、海洋潮汐及植被演替等多因素影响,闽江口湿地的环境条件极为复杂[13],由此可能直接或间接影响到湿地土壤中不同形态磷的赋存及其转化。植被的空间扩展是指不同物种之间对光、水分、养分等环境资源以及生存空间的竞争[14]。芦苇(Phragmitesaustralis)和短叶茳芏(Cyperusmalaccensis)是闽江口两种重要的原生植被,其对区域环境具有良好的适应性。近年来,芦苇与短叶茳芏在闽江口鳝鱼滩北部形成了明显的交错带,宽度达100—150 m,且有逐年变宽的趋势[15]。Ewanchuk和Bertenes[16]的研究表明,湿地植被之间的相互扩展不仅可影响不同植被的生态特征和养分吸收利用状况,而且还可能显著改变湿地群落的结构和演替过程,进而影响到湿地系统的稳定性。何涛等[13]和王华等[15]的进一步研究表明,该区芦苇与短叶茳芏的空间扩展是双向过程,不同植被在空间相互扩展过程中可采用不同的生态竞争策略。当前,关于闽江口湿地土壤中不同活性磷组分赋存的相关研究还比较缺乏,而关于不同植被空间扩展影响下湿地土壤不同磷赋存形态的相关研究还鲜有报道。特别是芦苇与短叶茳芏的空间扩展对交错带湿地土壤中不同形态磷的赋存及变化究竟有何影响尚不明确。为此,本研究选取闽江口鳝鱼滩的芦苇湿地、短叶茳芏湿地以及芦苇与短叶茳芏空间扩展形成的交错带湿地为研究对象,探讨了两种植物空间扩展对湿地土壤磷赋存形态及其变化特征的影响。研究结果有助于揭示不同湿地植被之间的磷养分竞争影响,并可为该区的湿地生态保育提供重要科学依据。
1 研究区概况与研究方法
1.1 研究区概况
闽江口地处我国中亚热带和南亚热带过渡区,气候暖热潮湿,雨热同期,年平均气温19.3 ℃,年降水量超过1300 mm,河口区发育有良好的湿地生态系统。本研究区位于闽江口最大的一块湿地——鳝鱼滩(119°34′12″—119°40′40″E,26°00′36″—26°03′42″N),土壤类型为滨海盐土,主要植被包括短叶茳芏(Cyperusmalaccensis)、芦苇(Phragmitesaustralis)、互花米草(Spartinaalterniflora)和扁穗莎草(Cyperuscompressus)等,植被群落由陆向海整体呈条带状分布。芦苇是一种本地入侵物种,它是大约30—40年前由闽江中上游逐渐扩散至闽江口的鳝鱼滩[17]。现有研究表明,闽江口芦苇和短叶茳芏的竞争能力相当,二者的空间扩展是双向过程,但芦苇具有相对竞争优势[13,15]。伴随着二者的空间扩展,由此形成了两种植物的交错带。
1.2 研究方法
1.2.1样地布设与样品采集
采用定位研究方法,于2016年7月在鳝鱼滩西北部选择纯芦苇湿地(P.australis,记为PA)、纯短叶茳芏湿地(C.malaccensis,记为CM)以及两种植物空间扩展形成的交错带湿地(P.australis-C.malaccensis,记为PA-CM)为研究对象,采用土壤柱状采样器在上述典型样地内进行土壤样品采集,采集深度为60 cm(每10 cm一层)。每个样地采集3个土柱作为重复,共计54个样品。不同湿地表层土壤的理化性质见表1。

表1 不同湿地表层土壤理化性质
1.2.2样品处理与测定
将采集的土壤样品置于实验室自然风干,研磨后过100目筛后装袋待测。土壤磷组分采用Hedley 连续浸提法测定: 取0.5 g过100目筛的土,逐级加入阴离子树脂膜、0.5 mol/L NaHCO3、0.1 mol/L NaOH和1 mol/L HCl提取出土壤中稳定性由弱到强的各级土壤磷组分,最后用浓硫酸和高氯酸高温消煮测定残留在土壤中的磷。根据Maranguit等[18]的划分方法,主要磷组分包括:(1)易分解态磷:Resin-Pi(树脂磷)、NaHCO3-Pi(碳酸氢钠态无机磷)、NaHCO3-Po(碳酸氢钠态有机磷);(2)中等易分解态磷:NaOH-Pi(氢氧化钠态无机磷)、NaOH-Po(氢氧化钠态有机磷)、Sonic-Pi(超声态无机磷)、Sonic-Po(超声态有机磷)。本研究中Sonic-Po极低,未检出;(3)难分解态磷:HCl-Pi(盐酸态无机磷)、Residual-P(残留态磷)。本研究中HCl-Pi为稀盐酸提取态磷。上述8种磷组分之和为总磷(TP)。不同形态磷含量通过连续流动分析仪(SKALAR-SAN++荷兰)测定。采用HACH-sensION3和ECTestr11+原位测定土壤pH和EC;采用高温外热重铬酸钾容量法测定土壤有机质含量;采用MasterSizer 2000激光粒度分析仪测定土壤粒度,并采用国际制分类;采用环刀法测定土壤容重;采用土壤墒情速测仪测定(TZS- 1)土壤含水量;采用高压酸消解ICP-MS 测定土壤Fe、Al含量。
1.2.3指标计算
土壤P储量(SP, g/m2)采用下式计算:
式中,SPi为第i层土壤磷库储量(g/m2);dvi为第i层土壤容重(g/cm3);Pi为第i层土壤磷含量(mg/g),hi为土壤深度(cm)。
1.2.4数据处理与分析
运用 Origin 8.0 软件对数据进行作图和计算。采用SPSS 20.0 软件对数据进行Pearson相关分析、方差分析和逐步线性回归分析,显著性水平设定为P=0.05。运用Canono 5.0软件进行主成分(PCA)分析。
2 结果与分析
2.1 湿地土壤易分解态磷含量变化
芦苇与短叶茳芏空间扩展过程中,3种湿地0—10 cm土层的Resin-Pi含量整体表现为交错带湿地>芦苇湿地>短叶茳芏湿地,而在10—40 cm土层则表现为芦苇湿地>交错带湿地>短叶茳芏湿地(图1)。其中,芦苇湿地与短叶茳芏湿地在10—20 cm土层以及芦苇湿地与交错带湿地在20—30 cm土层均存在显著差异(P<0.05)。就NaHCO3-Pi和NaHCO3-Po而言,除0—10 cm土层外,其含量在3种湿地10—60 cm土层整体均表现为交错带湿地>短叶茳芏湿地>芦苇湿地。其中,芦苇湿地与交错带湿地10—20 cm土层的NaHCO3-Pi含量存在极显著差异(P<0.01),而芦苇湿地与短叶茳芏湿地10—20 cm土层的NaHCO3-Pi含量则存在显著差异(P<0.05)。芦苇湿地与交错带湿地、交错带湿地与短叶茳芏湿地20—30 cm土层的NaHCO3-Pi含量存在极显著差异(P<0.01);NaHCO3-Po含量仅在芦苇湿地与交错带湿地的20—30 cm土层存在显著差异(P<0.05)。

图1 湿地土壤磷形态水平分布特征Fig.1 Horizontal distributions of the contents of phosphorus forms in marsh soils 图中不同字母表示数据间在P <0.05水平上差异显著; PA: 芦苇湿地 P. Australis marsh; PA-CM: 交错带湿地 P. Australis-C. Malaccensis marsh; CM: 短叶茳芏湿地 C. Malaccensis marsh
芦苇与短叶茳芏的空间扩展亦对不同湿地土壤中易分解态磷含量的垂直变化产生较大影响(图2)。3种湿地土壤的Resin-Pi含量均自表层向下呈先增加后降低变化,但其在不同湿地间的差异并不显著(P>0.05)。与之不同,3种湿地土壤的NaHCO3-Pi含量整体均自表层向下呈降低变化,且其在芦苇湿地与交错带湿地之间存在极显著差异(P<0.01)。就NaHCO3-Po而言,其含量在芦苇湿地与交错带湿地整体均随深度增加呈先增加后降低变化,而在短叶茳芏湿地则呈逐渐降低趋势。比较而言,芦苇湿地与交错带湿地NaHCO3-Po含量存在极显著差异(P<0.01),与之不同,芦苇湿地与短叶茳芏湿地的NaHCO3-Po含量存在显著差异(P<0.05)。与芦苇湿地和短叶茳芏湿地相比,交错带湿地土壤中的NaHCO3-Pi和NaHCO3-Po平均含量均呈增加趋势,增幅分别为25.3%、21.3%(NaHCO3-Pi)和 167.3%、11.0%(NaHCO3-Po)。与之不同,交错带湿地土壤中的Resin-Pi含量较芦苇湿地降低了12.3%,而较短叶茳芏湿地升高了20.3%。易分解态磷含量的变异系数整体表现为芦苇湿地>短叶茳芏湿地>交错带湿地,均为中等变异(表2)。
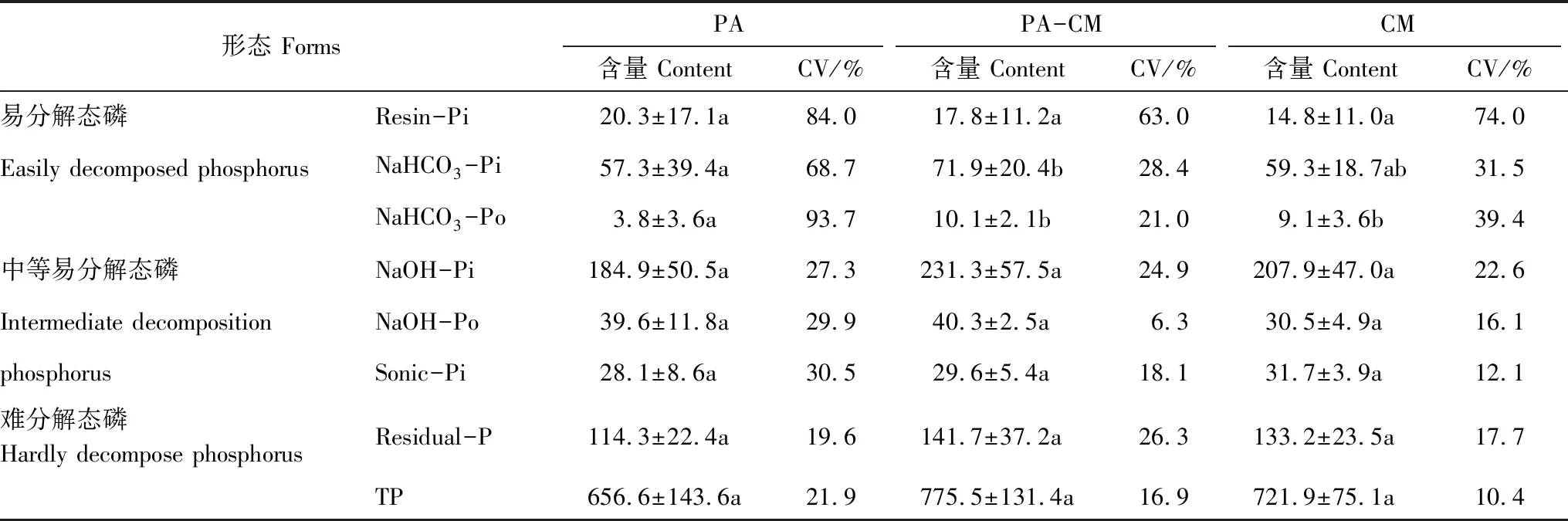
表2 不同湿地土壤中各形态磷含量均值及垂直变异系数
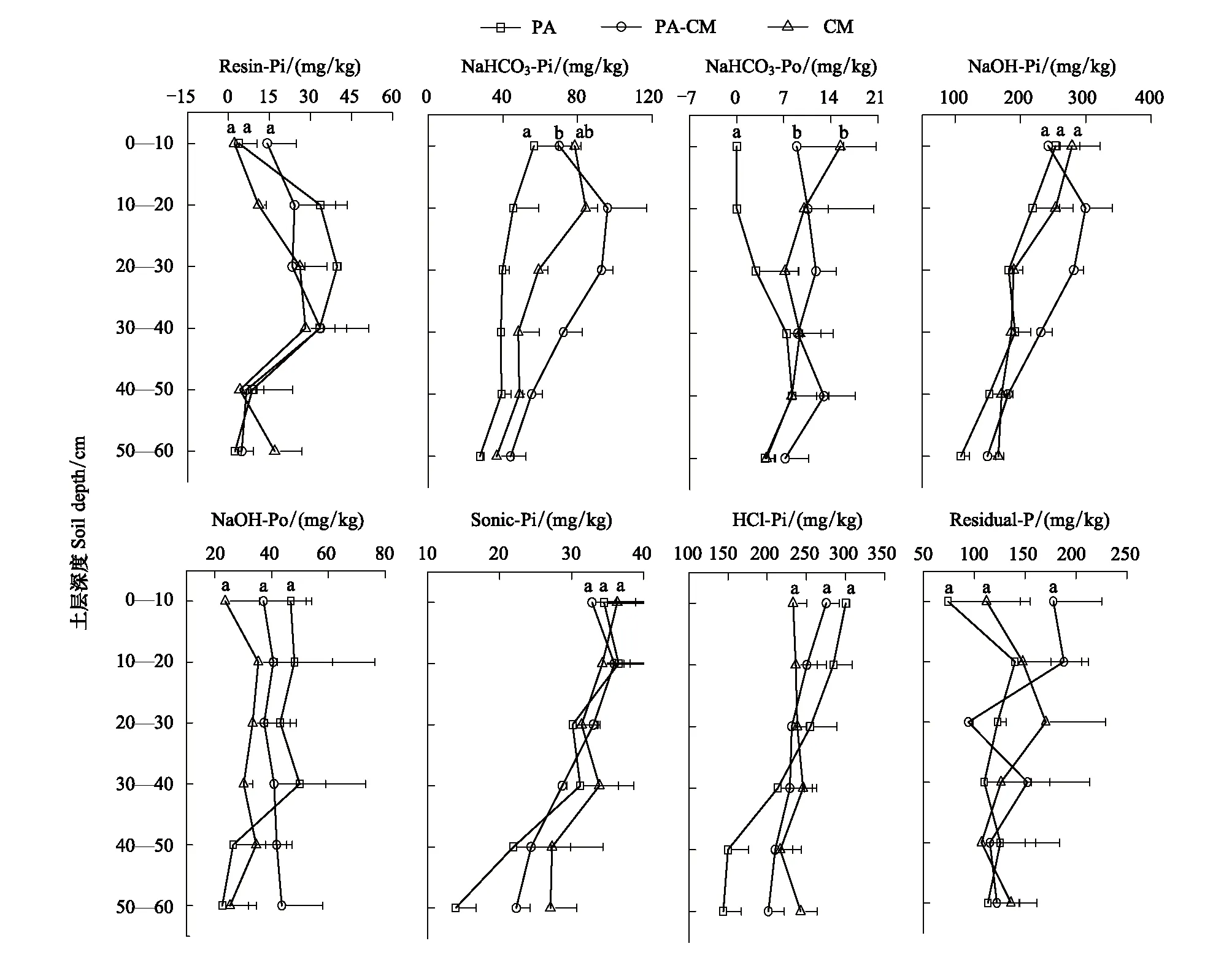
图2 湿地土壤磷形态垂直分布特征Fig.2 Vertical distributions of the contents of phosphorus forms in marsh soilsResin-Pi: 树脂磷 NaHCO3-Pi: 碳酸氢钠态无机磷; NaHCO3-Po: 碳酸氢钠态有机磷; NaOH-Pi: 氢氧化钠态无机磷; NaOH-Po: 氢氧化钠态有机磷; Sonic-Pi: 超声态无机磷; HCl-Pi: 盐酸态无机磷; Residual-P: 残留态磷
2.2 湿地土壤中等易分解态磷含量变化
芦苇与短叶茳芏空间扩展过程中,3种湿地0—10 cm土层的NaOH-Pi含量整体表现为短叶茳芏湿地>芦苇湿地>交错带湿地,而在10—60 cm土层则表现为交错带湿地>短叶茳芏湿地>芦苇湿地(图1)。其中,芦苇湿地与交错带湿地以及交错带湿地与短叶茳芏湿地20—30 cm土层的NaOH-Pi含量均存在极显著差异(P<0.01)。就NaOH-Po而言,其含量在0—40 cm土层表现为芦苇湿地>交错带湿地>短叶茳芏湿地,但在40—60 cm土层则表现为交错带湿地>短叶茳芏湿地>芦苇湿地。不同湿地各土层的NaOH-Po含量均不存在差异(P>0.05)。
在垂直方向上,3种湿地土壤中NaOH-Pi和Sonic-Pi含量的变化趋势较为一致,整体均自表层向下呈不断降低变化;但三者NaOH-Po含量的变化趋势并不一致(图2)。其中,芦苇湿地土壤中的NaOH-Po含量整体随深度增加呈降低趋势,而交错带湿地与短叶茳芏湿地土壤中的NaOH-Po含量均随深度增加而变化不大。比较而言,3种湿地土壤中NaOH-Pi、NaOH-Po和 Sonic-Pi含量在垂直方向上均不存在差异(P>0.05)。与芦苇湿地和短叶茳芏湿地相比,交错带湿地土壤中的NaOH-Pi平均含量均呈增加趋势,增幅分别为25.1%和11.2%。与之不同,交错带湿地土壤中的NaOH-Po含量较芦苇湿地降幅不大(仅为0.02%),而较短叶茳芏湿地增加了32.2%。此外,交错带湿地土壤中的Sonic-Pi含量较芦苇湿地升高了5.4%,而较短叶茳芏湿地降低了6.8%。尽管3种湿地土壤中中等易分解态磷含量的变异系数均较易分解态磷有所降低,但仍为中等变异(表2)。
2.3 湿地土壤难分解态磷含量变化
芦苇与短叶茳芏空间扩展过程中,3种湿地土壤中的HCl-Pi含量在0—20 cm土层表现为芦苇湿地>交错带湿地>短叶茳芏湿地,但在40—60 cm土层则表现为短叶茳芏湿地>交错带湿地>芦苇湿地(图1)。其中,芦苇湿地与短叶茳芏湿地的HCl-Pi含量在0—10 cm土层上存在极显著差异(P<0.01),而芦苇湿地与交错带湿地以及芦苇湿地与短叶茳芏湿地的HCl-Pi含量在40—50和50—60 cm土层均存在显著差异(P<0.05)。不同湿地土壤的Residual-P含量较为接近,且其之间差异亦不显著。
在垂直方向上,芦苇湿地与交错带湿地土壤中的HCl-Pi含量整体均自表层向下呈骤然降低变化,而短叶茳芏湿地土壤中的HCl-Pi含量则呈较小波动变化(图2)。就Residual-P而言,其在3种湿地土壤中虽均呈较大波动变化,但其垂直变化特征并不一致(P>0.05)。与芦苇湿地和短叶茳芏湿地相比,交错带湿地土壤中的Residual-P含量整体均呈增加趋势,增幅分别为24.0%和6.4%。与之不同,交错带湿地土壤中的HCl-Pi含量较芦苇湿地增加了3.8%,而较短叶茳芏湿地却降幅不大(仅为0.01%)。难分解态磷含量的变异系数整体以芦苇湿地最高,交错带湿地次之,短叶茳芏湿地最低(表2)。
2.4 湿地土壤总磷含量与储量
芦苇湿地、交错带湿地和短叶茳芏湿地土壤的TP平均含量分别为(656.6±143.6)mg/kg、(775.5±131.4)mg/kg和(721.9±75.1)mg/kg(表2)。相对于芦苇湿地和短叶茳芏湿地,交错带湿地土壤的TP含量增幅分别为20.03%和7.12%。整体而言,0—50 cm土层的TP含量均表现为交错带湿地>交错带湿地>短叶茳芏湿地,而在50—60 cm土层则表现为芦苇湿地>交错带湿地>短叶茳芏湿地(图3)。交错带湿地土壤中,易分解态磷、中等易分解态磷和难分解态磷含量分别占TP含量的12.9%、38.8%和48.3%,而在芦苇湿地和短叶茳芏湿地中,三者比例分别为10.0%、38.4%、51.6%和11.5%、37.4%、51.1%。与芦苇湿地与短叶茳芏湿地相比,交错带湿地土壤中的易分解态磷占比分别增加了2.9%与1.4%,中等易分解态磷的占比分别增加了0.4%与1.4%,而难分解态磷则降低了3.3%与2.8%。具体到某一形态,3种湿地土壤中的NaOH-Pi、HCl-Pi和Residual-P占TP的比例较高,而Resin-Pi、NaHCO3-Pi、NaHCO3-Po、NaOH-Po和Sonic-Pi占TP的比例较低。TP含量的垂直变异以芦苇湿地最高,短叶茳芏湿地最低(表2)。3种湿地土壤的磷储量在垂直方向上均随深度增加呈降低趋势,且不同土层的磷储量整体均以交错带湿地较高(图3)。总体而言,3种湿地0—60 cm土层的磷储量整体表现为交错带湿地(70.6 g/m2)>短叶茳芏湿地(62.3 g/m2)>芦苇湿地(59.5 g/m2)。与芦苇湿地与短叶茳芏湿地相比,交错带湿地土壤的磷储量分别增加了12.0%和18.0%。其中,NaOH-Pi、HCl-Pi和Residual-P在TP储量中的占比相对较高,分别介于28.1%—30.6%、29.7%—34.0%和17.5%—18.5%;而NaHCO3-Po、Resin-Pi和Sonic-Pi的占比相对较低,分别介于0.9%—1.3%、2.0%—3.0%、3.8%—4.2%(图3)。

图3 湿地土壤TP含量、储量及其组成Fig.3 Total phosphorus contents, phosphorus stocks and its composition in marsh soils
3 讨论
3.1 本研究与国内外相关研究对比
与国外相关研究相比,闽江口湿地土壤的TP含量处于中等水平(表3)。就不同磷组分而言,印度卡尔帕克金海岸盐沼土壤中易分解态磷含量与闽江口相当,中等易分解态磷含量明显低于闽江口,而难分解态磷含量明显高于闽江口,这主要与该区盐沼土壤砂粒含量占比较高,而闽江口湿地土壤粉粒含量占比较高(表1)有关。Stone和English[19]的研究证实,中等易分解态磷含量与土壤粒度呈反比,而这可能是导致印度卡尔帕克金海岸盐沼土壤中等易分解态磷含量较低的一个重要原因。另外,巴西瓜纳巴拉海湾盐沼土壤中各组分磷含量的变幅较大,且与闽江口相比含量值偏低,原因可能与其采样于冬季和夏季进行,而本研究仅在夏季进行有关。不同采样季节的温度差异较大,由此可能导致有机物分解和有机磷矿化速率存在较大差异[20],进而使得土壤中磷组分含量在不同采样季节的变幅较大。
与国内的相关研究相比,闽江口湿地土壤的TP含量在我国整体处于中等水平。其中,易分解态磷和中等易分解态磷含量高于黄河口湿地和松嫩平原湿地,但明显低于三江平原淡水湿地;难分解态磷含量明显高于松嫩平原湿地,但明显低于黄河口湿地,而与三江平原淡水湿地相当(表3)。三江平原淡水湿地由于地处东北,相对较低的温度条件使得土壤有机质降解缓慢,长期的养分积累导致本地区的土壤肥力较高[21],加之近年来该区湿地得到有效保护,由此导致其土壤中的TP含量很高。与之相比,松嫩平原地处半干旱地区,生态环境脆弱,土壤贫瘠[22],故其土壤中的TP含量较低。滨海湿地特别是河口湿地由于受河流与海洋的双重影响,故表现出与淡水湿地不同的磷组分分布状况。对比研究发现,随着纬度的降低,黄河口湿地、杭州湾湿地、闽江口湿地和珠江口湿地土壤中的TP含量整体呈增加趋势。本研究中,闽江口湿地受成土母质的影响,土壤中富含大量Fe、Al酸性沉积物。Fe、Al可与磷酸盐结合,导致本研究区土壤中的中等易分解态磷含量远高于北方的黄河口湿地。另外,由于北方黄河口湿地土壤中富含Ca、Mg离子,使得磷通常与其结合而构成难分解态磷的主要成分,而这是导致其难分解态磷含量及其占TP比例明显高于本研究区湿地土壤的一个重要原因(表3)。

表3 国内外湿地表层土壤磷含量对比/(mg/kg)
3.2 空间扩展对湿地土壤磷赋存形态的影响
本研究表明,芦苇与短叶茳芏的空间扩展明显改变了不同形态磷的分布特征,二者形成交错带湿地土壤中的易分解态磷、中等易分解态磷和难分解态磷含量整体均要高于相应的纯群落湿地。相对于芦苇湿地和短叶茳芏湿地,交错带湿地土壤中的难分解态磷含量分别提高了10.6%和1.6%,中等易分解态磷含量分别提高了19.2%和11.5%,而易分解态磷含量分别提高了22.6%和16.6%。原因可能与二者空间扩展过程中交错带湿地土壤的理化性质特别是粒度组成、pH以及Fe、Al含量均较纯群落湿地均发生明显改变有关(表1)。粒径分布不仅影响TP含量,而且影响其形态组成;粒径越小,生物可利用磷含量一般越高,其在TP中的比例也越高[25]。本研究中,交错带湿地土壤中较高的易分解态磷、中等易分解态磷和难分解态含量可能与芦苇与短叶茳芏扩展过程中二者对悬浮颗粒物的拦截能力较纯群落明显增强,进而导致其粒径组成发生明显改变有关。已有研究表明,在芦苇和短叶茳芏形成的交错带,二者因高度差异,芦苇占据高处空间,而短叶茳芏占据低处空间,而这种高低空间搭配导致其对悬浮颗粒物的拦截能力相比纯群落更强[13]。本研究中,交错带湿地表层土壤中的黏粒和粉粒含量相对于芦苇湿地增加了11.8%,而相对于短叶茳芏湿地升高了29.1%,表明两种植物扩展增加了土壤中的黏粒和粉粒含量(表1)。交错带湿地土壤中细颗粒的增加一方面可提高对易分解态磷和中等易分解态磷的吸附能力,另一方面大量的易分解态磷、中等易分解态磷和难分解态磷养分亦可随细颗粒被拦截而输入到湿地土壤中。pH也是影响湿地土壤中不同磷组分赋存的重要因素,特别是在一定酸碱度范围内,酸碱度变化可导致磷离子从矿物表面脱附并重新吸附到其他位置,从而改变土壤中的磷组分[26]。本研究中,交错带湿地土壤的pH值略高于短叶茳芏湿地,但却低于芦苇湿地。类似的,3种湿地土壤中的Fe、Al含量整体表现为交错带湿地>芦苇湿地>短叶茳芏湿地,与以Fe、Al吸附态磷为主的中等易分解态磷含量的变化较为一致,说明芦苇与短叶茳芏的空间扩展极有可能会通过改变土壤pH及Fe、Al含量来影响土壤中中等易分解态磷的赋存。
就不同磷组分而言,其在湿地土壤中整体表现为HCl-Pi(26.49%—36.99%)>NaOH-Pi(25.00%—35.79%)>Residual-P(11.67%—25.92%)>NaHCO3-Pi(5.55%—11.53%)>NaOH-Po(3.05%—7.39%)>Sonic-Pi(3.18%—4.78%)>Resin-Pi(0.28%—5.56%)>NaHCO3-Po(0.68%—2.00%)。其中,难分解态磷占TP的比例最高(48.3%—51.1%),中等易分解态磷次之(37.4%—38.8%),而易分解态磷最低(11.5%—12.9%)。虽然易分解态磷含量的占比较低,但却是生物容易获取且有效性较高的重要磷组分。由于磷的溶解性有限,且易于吸附到有机物和矿物表面,几乎所有植物都进化出专门的机制来增强从土壤中获取磷的方式,故其磷的获取量在不同生境中往往差异较大[27]。本研究中,交错带湿地土壤中的Resin-Pi含量相对于芦苇湿地降低了12.3%,而相对于短叶茳芏湿地增加了20.2%,说明短叶茳芏在交错带中相比纯群落可获得更多的Resin-Pi养分,从而有助于提高其在交错带中的竞争优势。NaHCO3提取态磷包括NaHCO3-Pi和 NaHCO3-Po。由于这两部分磷均是有效的,可在Resin-Pi被植物吸收利用后及时补充转化,因此对植物生长具有重要意义。本研究中,相对于芦苇湿地和短叶茳芏湿地,交错带湿地土壤中的NaHCO3提取态磷含量分别增加了34.2%和19.9%,说明交错带中的两种植被均可通过大量吸收NaHCO3提取态磷来保持各自竞争优势。NaOH提取态磷主要是以化学吸附作用吸附于土壤Fe、Al表面的磷,即中等易分解态磷,包括有机(NaOH-Po)和无机(NaOH-Pi和Sonic-Pi)两部分。本研究中,其含量大致占TP的33%。相对于芦苇湿地和短叶茳芏湿地,交错带湿地土壤中的NaOH提取态磷含量分别增加了19.2%和11.5%。尽管NaOH提取态磷较难被植物吸收利用,但其在交错带湿地土壤中的增加对于保障芦苇和短叶茳芏在交错带湿地中的竞争或空间扩展提供了充足的潜在磷养分。磷灰石型磷(HCl-P)是在高度风化的土壤中可提取出来的部分闭蓄态磷,残留磷(Residual-P)为不能提取的非常稳定的磷。本研究中,相对于芦苇湿地和短叶茳芏湿地,交错带湿地土壤中的两种磷含量分别增加了10.6%和1.6%。虽然难分解态磷含量较为稳定,但可通过解吸、风化和矿化过程而被植物吸收利用,从而充当潜在磷库的作用。综上,相较于纯群落,芦苇和短叶茳芏的空间扩展导致了交错带湿地土壤中绝大部分磷形态含量的明显增加,而这主要与前述交错带群落对悬浮颗粒物的较强拦截作用导致大量细颗粒和磷养分输入密切相关。
本研究还表明,芦苇与短叶茳芏的空间扩展整体改变了湿地土壤中的TP含量(图4)。相对于芦苇湿地和短叶茳芏湿地,交错带湿地土壤的TP含量分别增加了20.0%和7.1%,而磷储量分别增加了12.0%和18.0%。由于土壤TP含量特别是上述易分解态磷和中等易分解态磷含量可较好地指示土壤速效磷水平[28],而速效磷又是评价土壤供磷能力的重要指标[6],加之磷是本研究区湿地的重要限制性养分,故交错带湿地土壤中较高的TP以及易分解态磷和中等易分解态磷含量在一定程度上体现了其具有较强的供磷潜力。据此可知,本研究中交错带湿地的磷赋存状况有助于缓解芦苇与短叶茳芏在空间扩展过程中对磷养分的竞争压力,从而可在一定程度上维持交错带湿地系统的相对稳定。
3.3 湿地土壤不同磷形态赋存的关键因子识别
为进一步明确影响芦苇与短叶茳芏空间扩展过程中湿地土壤不同形态磷含量分布的关键因素,对3种湿地土壤磷组分与环境因子进行主成分分析。结果表明,芦苇湿地提取的两个主成分(PCA1和PCA2)累计贡献率为88.62%(图4)。其中,PCA1的贡献率为50.10%,可看作是土壤颗粒组成(黏粒、粉粒和砂粒)的代表;PCA2的贡献率为38.52%,可看作是可溶性盐(EC)和pH的代表。进一步分析发现,Resin-Pi和NaOH-Po与PCA1存在较强相关性,而NaHCO3-Pi和NaOH-Pi与PCA2存在较强相关性。交错带湿地提取的两个主成分(PCA1和PCA2)累计贡献率达92.07%。其中,PCA1的贡献率为52.30%,可看作是土壤颗粒组成(粉粒、砂粒和黏粒)的代表;PCA2的贡献率为39.77%,可看作是可溶性盐(EC)和有机质的代表。进一步分析发现,NaHCO3-Pi和NaOH-Pi与PCA1存在较强相关性,而Sonic-Pi与PCA2存在较强相关性。短叶茳芏湿地提取的两个主成分(PCA1和PCA2)累计贡献率达92.97%。其中,PCA1的贡献率为85.55%,可看作是土壤颗粒组成(粉粒和砂粒)组成和可溶性盐(EC)的代表;PCA2的贡献率为7.42%,可看作是有机质的代表。进一步分析发现,NaHCO3-Pi和NaOH-Pi与PCA1存在较强相关性,而Resin-Pi与PCA2存在较强相关性。

图4 湿地土壤磷赋存形态与环境因子主成分分析Fig.4 Principal component analysis for phosphorus forms and environmental factors in soils of different marshesEC: 电导率 Electrical conductivity; SOM: 土壤有机质Soil organic matter; BD: 土壤容重Soil bulk density
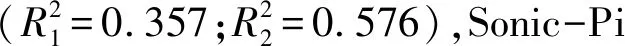
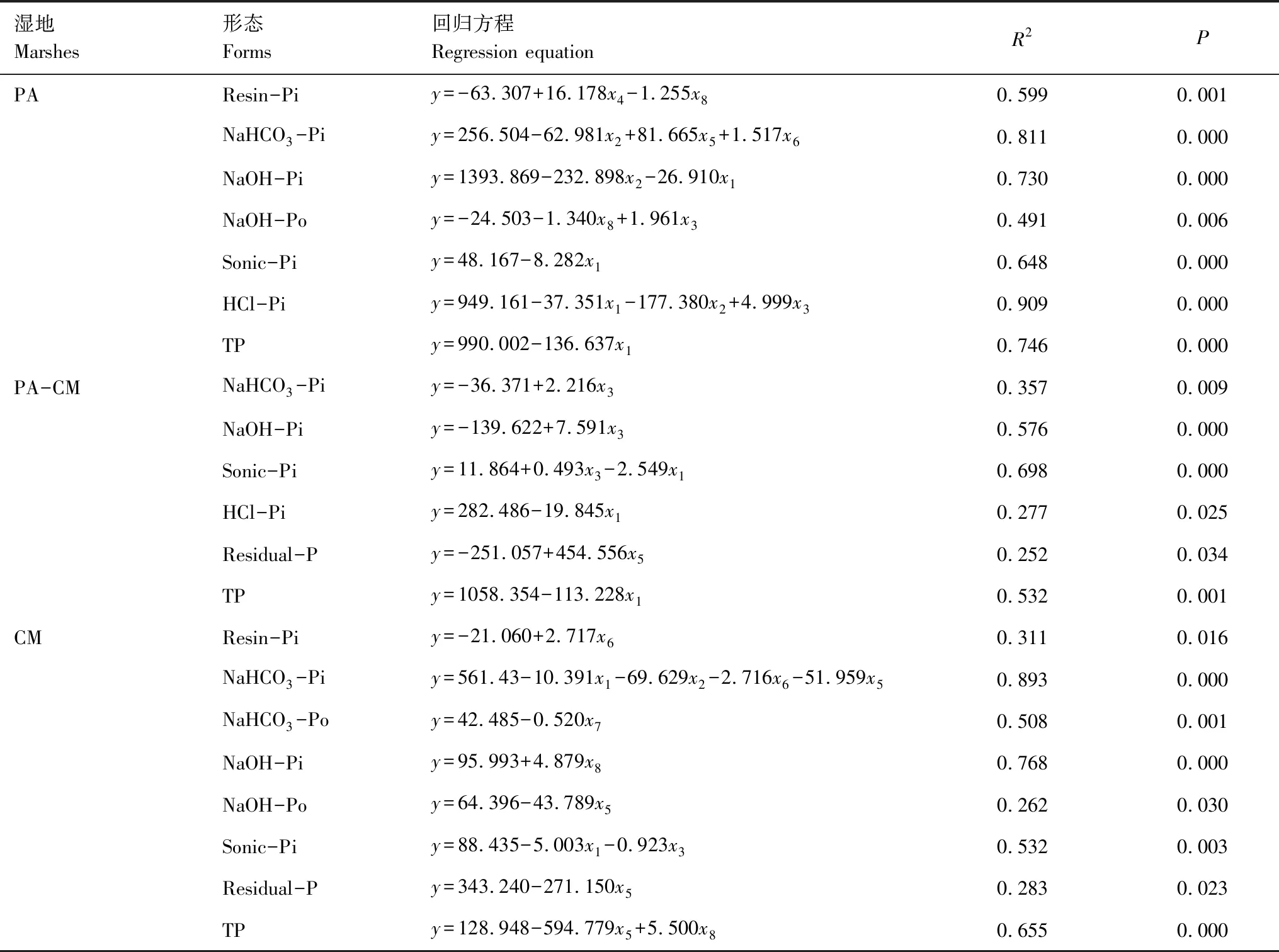
表4 湿地土壤中不同磷组分与环境因子之间逐步线性回归分析
不同磷赋存形态间的相关分析表明,3种湿地中NaHCO3-Pi与NaOH-Pi均呈极显著正相关(P<0.01),NaOH-Pi与Sonic-Pi均呈显著正相关(P<0.05),Sonic-Pi与HCl-Pi均呈显著相关(P<0.05)。与之不同,芦苇湿地和交错带湿地中的NaHCO3-Pi与HCl-Pi均呈极显著正相关(P<0.01),而短叶茳芏湿地并不存在相关规律。上述结果表明,不同磷形态之间关系密切且可能来自同一个源,即磷矿化过程。据表1可知,芦苇与短叶茳芏的空间扩展改变了湿地土壤理化性质,特别是交错带湿地土壤的颗粒组成、pH值以及Fe、Al含量与芦苇湿地或短叶茳芏湿地存在显著差异。前述主成分分析又进一步发现,芦苇湿地主要受土壤颗粒组成(黏粒、粉粒和砂粒)、EC和pH的影响,短叶茳芏湿地主要受土壤颗粒组成(粉粒和砂粒)、EC和有机质的影响,而交错带湿地主要受土壤颗粒组成(粉粒、砂粒和黏粒)、EC和有机质的影响,说明两种植物的空间扩展在一定程度上改变了影响土壤中不同磷形态的关键因素,而这种改变与表1显示的湿地土壤理化性质的改变基本一致。因此,芦苇与短叶茳芏的空间扩展极有可能会通过影响湿地土壤的磷矿化过程而改变土壤中不同形态磷的赋存以及其之间的转化(表5)。

表5 湿地土壤中不同磷赋存形态之间的相关分析
4 结论
(1)不同湿地土壤中各形态磷含量整体均表现为HCl-Pi>NaOH-Pi>Residual-P>NaHCO3-Pi>NaOH-Po>Sonic-Pi>Resin-Pi>NaHCO3-Po。其中,难分解态磷占TP的比例最高(48.3%—51.1%),中等易分解态磷次之(37.4%—38.8%),而易分解态磷最低(11.5%—12.9%)。
(2)芦苇与短叶茳芏在空间扩展形成的交错带湿地土壤中易分解态磷、中等易分解态磷和难分解态磷含量相对于纯群落湿地均发生了明显改变,其值相比芦苇湿地分别提高了10.6%、19.2%和22.6%,相比短叶茳芏湿地分别提高了1.6%、11.5%和16.6%,原因主要与二者空间扩展过程中交错带湿地土壤理化性质特别是粒度组成、pH以及Fe、Al含量均较纯群落湿地发生明显改变有关。
(3)芦苇与短叶茳芏的空间扩展整体改变了湿地土壤的TP含量和储量,相对于芦苇湿地和短叶茳芏湿地,交错带湿地土壤的TP含量分别增加了20.0%和7.1%,而磷储量分别增加了12.0 %和18.0%,说明交错带湿地土壤具有较强的供磷能力。
(4)芦苇与短叶茳芏的空间扩展不但改变了湿地土壤中磷的赋存状况,而且亦可能改变不同磷形态之间的转化。交错带湿地土壤的磷赋存状况有助于缓解芦苇与短叶茳芏空间扩展过程中对磷养分的竞争压力,从而可在一定程度上维持交错带湿地系统的相对稳定。
猜你喜欢
海外文摘·文学版(2022年4期)2022-04-14 21:55:16
心声歌刊(2021年4期)2021-12-02 01:14:20
黄河之声(2021年19期)2021-02-24 03:25:24
闽都文化(2020年5期)2020-08-25 10:10:44
福建基础教育研究(2020年2期)2020-05-28 08:39:55
岁月(2018年2期)2018-02-28 20:40:58
珠江水运(2017年17期)2017-10-10 08:16:03
幼儿100(2016年28期)2016-02-28 21:26:18
中央民族大学学报(自然科学版)(2015年4期)2015-06-11 03:41:42
新疆大学学报(自然科学版)(中英文)(2014年2期)2014-11-06 07:49:06
