
单环刺螠浮游幼体饥饿实验及不可逆生长点的确定
2020-11-19 05:04丁勇李赟孙阳刘彤陈文博张赛赛宋晓阳陈颖张玢
中国水产 2020年10期
文/丁勇 李赟 孙阳 刘彤 陈文博 张赛赛 宋晓阳 陈颖 张玢
单环刺螠(Urechis unicinctus)隶属螠虫动物门、螠纲、无管螠目、刺螠科、刺螠属。生活在海边潮间带,我国主要分布在黄海、渤海沿岸,在当地俗称海肠,是一种经济价值较高的海产品种,现阶段市场零售价格能达到320元/kg,是渔业经济品种中重要的捕捞对象。
饥饿是海洋生物在自然水域生态系统中经常面临的一种生理现象,也是影响海洋生物正常生长、发育及生存的一个重要环境因子。王庆志等认为饥饿条件对魁蚶眼点幼虫占幼虫总数比例有一定影响,随着饥饿时间延长,幼虫发育变慢,一直饥饿会导致生长停滞。单环刺螠浮游幼体内源性营养时间很短,幼体主动摄食能力很强,需要尽快对其进行投饵才能完成幼体从内源性营养向外源性营养期的转化,以维持正常的早期发育生长。刺参、扇贝等浮游幼体受自身的形态发育、生理状态以及环境因子影响,容易出现摄食器官发育迟缓、游泳器官机械损伤和幼体变态等可引发饥饿死亡的问题,导致后期浮游幼体生长迟缓,死亡率陡增。单环刺螠浮游幼体的相关研究未见报道。
近年来由于过度捕捞等原因,单环刺螠数量急剧下降,很多传统的产地已经捕捞不到单环刺螠。同时由于近几年刺参育苗行业惨淡,越来越多的养殖户放弃刺参育苗,导致大量工厂化育苗车间一直处于闲置状态。为保护大连本地特有的单环刺螠种质资源,同时能够将闲置的工厂化育苗车间利用,推进农业现代化的科技供给侧改革,促进产业转型升级,本项目探索了单环刺螠工厂化生态养殖模式。本文主要探讨饥饿环境对单环刺螠浮游幼体生长的影响,并初步确定单环刺螠浮游幼体PNR(不可逆点)。
一、材料与方法
(一)浮游幼体及培育条件
1.初孵浮游幼体来源
2019年5月,于大连市金普新区荞麦山海区捕捞种螠,清洗干净表面后解剖,人工授精获得受精卵,于孵化池中充气孵化出担轮幼虫。
2.浮游幼体培育条件
初孵浮游幼体于80L圆柱形水桶中培育,砂滤海水,盐度30~31,溶解氧8.4mg/L~8.9mg/L,水温18.0℃~19.5℃,pH7.9~8.0。白天为自然光,光强范围为200lx~1000lx。每天记录水桶内水温,吸底换水一次,换水量50%。昼夜以微充气饲养。观察浮游幼体的生活习性及生长发育变化。
3.饵料
实验用饵料为新月藻、角毛藻和海洋红酵母,混合投喂。
(二)方法
1.浮游幼体饥饿实验
取当天孵化担轮幼虫,分别放养于标号为A、B、C的三个80L的水桶中。从担轮幼虫开口摄食前一天开始,每天定时往A水槽中投喂新月藻、角毛藻和海洋红酵母,使水槽中饵料密度保持在正常浓度,作为正常生长担轮幼虫。B、C水槽内不投喂任何饵料,作为饥饿担轮幼虫。每个水桶的担轮幼虫放养密度为6个/mL,每个水桶放养总个数48万个。
每天随机分别从A、B两个水槽中各取担轮幼虫10个,在显微镜下测量并记录其体长、体高、胃径。体积在担轮幼虫发育过程中为椭球形。体积计算公式为:V=(4/3)·π·(R/2)2·L/2,其中R为体高,L为体长。
2.初次摄食率和初次摄食强度
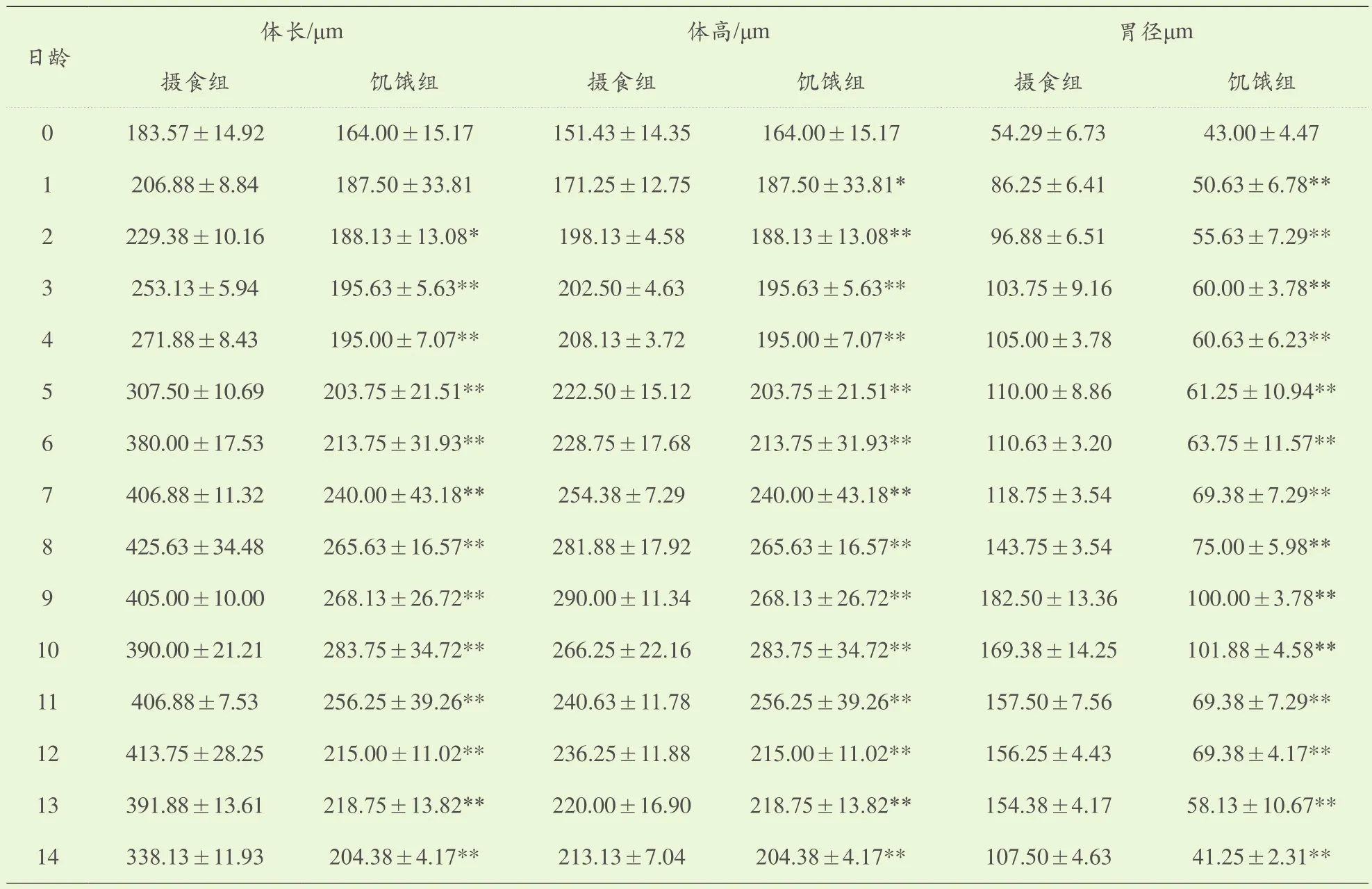
表1 摄食组和饥饿组单环刺螠浮游幼体测量表
单环刺螠担轮幼虫开口后每天从C水桶中取担轮幼虫20个,放入有效水体为40mL的透明塑料瓶中,不充气(单环刺螠耐低氧),投喂一定量新月藻、角毛藻和海洋红酵母,测定波长427nm下吸光光度值作为初始光度值。1h后将担轮幼虫全部取出,在显微镜下逐一观察个担轮幼虫的摄食情况,同时测量计算吸光光度值,计算初次摄食率和初次摄食强度,初次摄食率=摄食担轮幼虫数/样本担轮幼虫数×100%。每天从C组中取20个幼体于40mL透明塑料瓶中记C1,另一空40mL透明塑料瓶记C2。C1投饵后立刻测吸光值查询浊度记N1初,1h后测吸光值查询浊度记N1末,△N1=N1初-N1末。同理得△N2=N2初-N2末,初次摄食强度F=△N=△N1-△N2。
3.PNR(不可逆点)的确定
单环刺螠浮游幼体PNR确定,以单环刺螠孵化后日龄表示,每日测定饥饿组单环刺螠担轮幼虫的初次摄食率,当所测定的饥饿组担轮幼虫的初次摄食率低于最高初次摄食率的一半时,即为PNR(不可逆点)的时间。
(三)数据处理与计算
实验所得数据用SPSS17.0进行统计分析,结果以平均值±标准误表示。比较饥饿组担轮幼虫与摄食组担轮幼虫的生长指标采用t检验方法,以P<0.05为差异显著,P<0.01为差异极显著,用Excel2007运算并绘图。
二、结果
(一)摄食组和饥饿组单环刺螠浮游幼体生长的变化
在培育水温为18.0℃~19.5℃条件下,初孵浮游幼体体长(173.8±10.5)μm,体高(157.7±7.4)μm,胃径(48.6±7.1)μm。初孵浮游幼体游泳能力相对较弱,漂浮于水体表层,依靠体中部一圈纤毛环摆动进行运动。口位于体中部纤毛环初,消化道未分化。躯体呈椭球状。浮游幼体孵出后第2d开口摄食。15日龄饥饿组浮游幼体全部死亡。由表1可以看出饥饿组浮游幼体与摄食组浮游幼体在0~1日龄体长、体高及胃径变化差异均不明显,摄食组浮游幼体摄食后第2d开始加速增长,与饥饿组相比差异极显著(P<0.01)在摄食后的第7d,浮游幼体的体长和体高增长显著,与初孵浮游幼体相比分别增长了(232±10.5)μm和(96±7.4)μm,而与此对应的饥饿组浮游幼体的体长和体高分别增长了(66±5.2)μm和(82±4.8)μm。随着日龄增长,摄食组与饥饿组浮游幼体发育程度也不同,9日龄时,摄食组浮游幼体出现体环纹,开始发育至体节幼虫。而饥饿组浮游幼体直到15日龄,幼体全部死亡也未出现体环纹,未能发育到体节幼虫。随着饥饿时间的持续,饥饿组浮游幼体生长减慢、停止直至死亡。
(二)浮游幼体体积变化
浮游幼体的体积随日龄增加而先增加后减小(图1),初孵担轮幼虫的体积为2028174μm3,随着幼体的发育,摄食组体积增长很快,饥饿组体积增长缓慢。由图1可以看出,在18.0℃~19.5℃的条件下,5日龄到9日龄体积增长速度最快,10日龄开时体积开始减小,到实验结束时15日龄摄食组幼体体积8040022μm3,饥饿组幼体体积2313109μm3。9日龄担轮幼虫腹部分节明显,进入体节幼虫,体积开始逐渐减小。
(三)浮游幼体初次摄食率与PNR的确定
浮游幼体初次摄食率变化如图2所示。浮游幼体孵化后第二天开始摄食,也就是开始饥饿实验后第1d,此时浮游幼体初次摄食率为100%,一直维持在这个水平。直至第12日龄初次摄食率才开始下降95%,至第15日龄初次摄食率达到0%。根据PNR点定义,孵化后第15日龄,也就是饥饿15d,进入PNR期。图3为浮游幼体随着饥饿天数增加摄食能力的变化趋势。饥饿7d内,仔鱼摄食能力提高,饥饿7d后,仔鱼摄食能力降低。
三、讨论
(一)饥饿对单环刺螠浮游幼体生长及形态发育的影响
浮游幼体的生长发育除自身遗传性外,会受到一系列外界因素的影响,外源营养是其中重要的影响因子,外源营养缺乏会直接导致浮游幼体生存能力减弱,成活率降低。如果浮游幼体长期处于饥饿状态,则生长将会受到很大影响。饥饿后单环刺螠浮游幼体出现纤毛环萎缩,体壁皱缩,体高比摄食组低,体长比摄食组短,胃径比摄食组小,这与哲罗鱼、中华绒螯蟹、曼氏无针乌贼等海洋经济生物幼体饥饿实验结果基本相同。在螠虫类饥饿方面,目前国内外有关饥饿发育的研究较少。本实验测定了饥饿和摄食条件下单环刺螠生长的变化及发育状况。由结果可以看出,饥饿对其幼体的发育和摄食均有很大影响,海洋生物幼体阶段饵料充足是满足幼体正常生长发育的重要因子。
(二)初次摄食率与PNR
不同种类海洋生物幼体的初次摄食时间差异很大,冷水鱼类太平洋鳕仔鱼在破膜后第5日龄开始摄食,比高温鱼类黄鲷的4日龄、黑鮸的3日龄、大黄鱼的3日龄、囊鳃鲶3日龄晚,原因可能与温度有关。单环刺螠产卵孵化温度为18℃,温度较高,并且生物相对低等,消化结构简单,所以浮游幼体初次摄食时间更早。
摄食率的高低和最高摄食率持续的长短可以用来判断幼体的摄食能力,而PNR可以判定幼体耐受饥饿的能力。单环刺螠的最大初次摄食率为100%,出现在第2日龄,保持了近10d时间,与金乌贼仔乌PNR9.5d情况接近,而三疣梭子蟹的PNR出现在39h,这主要是受物种类别和温度的影响。
饵料是否充足将决定生物体后期的变态发育,甚至生长和存活,在最为恰当的时期进行合理投喂是十分必要的。由以上结果可以说明单环刺螠的摄食能力和耐饥饿能力很强。在人工育苗过程中,较强的耐饥饿能力能大大提高浮游期人工育苗的成活率,为探索成本最低的饵料配方提供了依据。单环刺螠开口较早,及时投喂饵料,确保初孵幼体尽快吃到食物,降低人工养殖过程中的死亡率,才能创造更大的经济效益。
(三)初次摄食最佳投喂时间
海洋生物幼体的摄食能力和耐饥饿能力需要掌握好,在人工育苗投喂适口天然饵料时,过早、过晚和过量投喂都会导致浮游幼体出现生长发育问题,甚至明显大量死亡。单环刺螠浮游幼体耐饥饿能力较强,开口摄食时间早,在18.0℃~19.5℃的条件下,单环刺螠担轮幼虫孵出后第2d即开口摄食,混合营养期很短,为1d。在混合营养期阶段,必须及时满足其外源性营养的需求,补充正常生理活动所需的饵料。若在此时期得不到食物,幼体后期的生长发育会受到明显影响。因此,单环刺螠浮游幼体最佳投喂时间为2日龄。
猜你喜欢
四川蚕业(2022年2期)2022-11-19
中国海洋大学学报(自然科学版)(2022年9期)2022-09-05
当代水产(2022年6期)2022-06-29
特种经济动植物(2022年2期)2022-02-15
当代水产(2021年1期)2021-03-19
四川蚕业(2021年1期)2021-02-12
流行色(2019年10期)2019-12-06
装饰装修天地(2019年9期)2019-10-21
河北渔业(2018年4期)2018-05-08
安徽农学通报(2016年24期)2017-01-12
