
基于水产动物视觉敏感性对人工光源选择的分析
2020-11-18 08:45:12宋昌斌
照明工程学报 2020年5期
宋昌斌, 李 贤, 史 策
(1.中国科学院半导体研究所,北京 100083;2.中国科学院海洋研究所,山东青岛 266071;3.宁波大学,浙江宁波 315832)
引言
2018年,中国渔业经济总产值为 25864.47 亿元,其中海水与淡水养殖产值达 9456.27 亿元[1],水产养殖是中国农业结构中发展最快的产业之一。然而,随着该产业的快速发展,粗放的养殖模式已经不适应消费者对优质水产品逐年增量的需求。水产养殖业一大特点是品种众多,以海水养殖为例,除藻、虾、蟹、贝,据不完全统计,我国现有海水养殖鱼类品种达30科、48属、80余种[2]。每种水生动物分别在内陆、近岸、大洋开阔[3]复杂的物理生化系统水域环境生存。在上亿年的适应环境进化中,形成了独特形态构造的视觉器官与神经细胞。每种水产动物,甚至在其不同的生命周期需要的光照环境不尽相同。目前大部分渔业生产者已经认识到LED光照对其捕捞、养殖品种的新陈代谢会起到积极作用,但对作用其品种的光生物学机理与效用规律知之甚少,LED制造、工程商家对此跨行业光照领域也缺乏相关知识。改变渔业传统照明观念,遵循实验数据技术支持的设计是今后发展方向[4]。比如实验结果表明蓝圆鲹(Decapterusmaruadsi)对绿光趋光率最高,竹荚鱼(Trachurusjaponicus)对橙光的反应大,红色光能使秋刀鱼(Cololabissaira)稳定地诱集在光照区,鲅鱼(Scomberomorusniphonius)对红、紫光有很强烈的兴奋作用[5]。Gao等[6]通过实验证明皱纹盘鲍(Haliotisdiscushanna)在红色或橙色光处理下胃蛋白酶、纤维素酶活性均显著高,生长率高。这些科学实证结果对渔船捕捞诱集、设施工厂化水产动物养殖人工光源的设计提供了有力的技术支撑。渔业用LED光照迫切需要光谱选择、灯具设计等一般性技术原则来有效地推动LED产业链跨行业的技术合作。
1 硬骨鱼类视觉的光谱敏感性概述
硬骨鱼类(Osteichthyes)是水中生活最繁盛的脊椎动物,有2万余种,占鱼类总数90%以上[7]。我国大宗水产养殖鱼类几乎都是头部有膜骨、内骨骼或多或少硬骨化的硬骨鱼类[8]。每一个硬骨鱼类品种对光的感觉与需求有很大差异,分析硬骨鱼类视觉的光谱敏感性及生长最适光环境有助于设计时采用针对性的科学依据,避免机械照搬一般性照明的设计原则。
1.1 硬骨鱼类视觉器官生理结构简述
硬骨鱼类视觉器官进化的形态构造与栖息的水域光环境密切相关,例如海水鱼眼角膜对光的折射系数为1.37,与海水对光的折射系数1.33很接近[9],因此,光线几乎可以从水中直接透过角膜进入晶状体,不会发生反射或折射损失,有利于更多光线投入视觉器官。硬骨鱼类眼结构与高等脊椎动物基本相似。鱼眼视网膜结构十分复杂,内层分布视杆与视锥二种视觉细胞,视杆细胞感受光线强弱,光强敏感性高,无辨色和精细分辨能力;视锥细胞感受光波长,有辨色与精细分辨能力[7,10]。鱼类对颜色光的反应,主要是视锥细胞的视觉功能。当视网膜受到光刺激时,根据光强与频率,光子带来的能量引起视色素分解等化学变化,生成的产物刺激视觉细胞,神经电脉冲沿神经纤维传到鱼脑视觉中枢,形成视觉。生活在光照条件好和依靠视觉捕食的鱼类,大部分可辨别颜色。Siebeck等[11]实验证明视锥细胞发达的珊瑚礁鱼类通过学习可辨别颜色。而视觉不发达的深海与夜间活动的鱼类,光刺激阈值低,光敏性强,大多数没有辨色能力[9]。鱼的视觉细胞特殊结构保障其在白天光线明亮和夜间昏暗环境中完成捕食生活,且在同一段时间垂直洄游于光线较强和较弱的水层。
1.2 光敏感性的一般规律
硬骨鱼类在长期进化过程中视觉器官必须适应栖息水域复杂的光学条件,完成各种生命活动。通过视网膜与松果体的光感受器接收光信号,以同步感知并适应自然栖息地的光环境。一般来说,经过长期进化鱼类视网膜所具有的细胞视色素和种群所在栖息地的光波峰是一致的。蓝刻齿雀鲷(Chrysipteracyanea)游弋围绕珊瑚礁,广泛分布在西太平洋的浅水中。Bapary等[12]实验证明利用长波长的光可诱导其卵巢发育。Tsutsumi 等[13]在4种光色饲养条件下,对太平洋蓝鳍金枪(Thunnusorientalis) 稚鱼的生长性能适合的波长范围进行实验,结果表明相对于红光,蓝绿对稚鱼体长体重有明显促进作用,同时用视网膜电流图(electroretinogram,ERG)法得到其稚鱼光谱适应的敏感性在449~503 nm波长范围内,这与其稚鱼阶段迁移到适宜的深海栖息地的光谱分布环境是一致的。蓝刻齿雀鲷、黑线鳕(Melanogrammusaeglefinus)、塞舌尔鳎(seychellessole)、大西洋鳕(Gadusmorhua)幼鱼视觉选择的敏感波长也与其生活环境光谱成分一致[14]。硬骨鱼类对光谱的响应很大程度上取决于物种栖息地的光环境:深海鱼的光谱敏感性通常在蓝色波段,而沿海及珊瑚礁鱼类在长波长波段呈现最大敏感度[11,12,15]。事实上,鱼类视觉色素已经适应他们栖息所在的或者清澈、或者浑浊、或者海洋深处及过渡带不同水域的光谱环境[15]。
1.3 鱼类光敏感性具有可塑性
概括来说,鱼类视觉色素由视蛋白组成,以视黄醛1为基础形成的视觉色素称为视紫红质,以视黄醛2为基础形成的视觉色素称为视紫质。前者以蓝紫色层活动的海鱼类为主,吸收光谱的范围为430~562nm,后者以在浮游植物和颗粒物质较多的近岸生活的,偏重于在可见光中长波长范围内的淡水鱼类为主,吸收光谱范围570~620nm,并且产卵场的盐度是决定视觉色素的主要因素[10]。需要考虑的是有一些鱼类视蛋白组成具有可塑性,呈现不同生长阶段随着栖息光环境的不同而有所变化具有二种色素的混合物,洄游及包括过河口性鱼类在不同的生命周期生活史的某一阶段,比如溯河与降海过程中一种色素可能超过另一种色素。欧洲舌齿鲈(Dicentrarchuslabrax)是欧洲海岸最常见丰富的鱼类物种之一,刘鹰团队[16-18]研究了不同LED 光色对平均体长(13.78±0.35)cm的欧洲舌齿鲈幼鱼分别就摄食、生长、消化能力与营养品质的影响,结果表明可见光长中长波长更适合欧洲舌齿鲈的养殖。但另一项实验表明欧洲舌齿鲈成鱼对434~447nm的蓝光敏感,并对褪黑激素有抑制作用[19]。崔鑫[20]从其栖息迁移角度分析这种现象,法国里昂湾欧洲舌齿鲈的体长12~22mm时,从产卵地洄游至近岸索饵场,当体长达到63~112mm时,再游回深海。我国大菱鲆(Scophthalmusmaximus)目前年产8~10万吨的产业规模,占我国鲆鲽类总产量的80%以上。大菱鲆为海鱼浮性卵,发育后从水上层游泳转变为底栖生活,光谱成分随之主要为蓝光。李贤研究小组[21,22]实验证明大菱鲆仔稚鱼最适光谱具有阶段特异可塑性,孵化后约为1~20天,大菱鲆最适光谱全光谱作为光源,摄食活跃,显著提高存活率。随着变态过程的进行,逐渐由全光谱向短波长的蓝光迁移。这些发现与大菱幼体在野外变态过程中所经历的光谱变化是一致的。鱼类的趋光性与生命周期有密切关系,绝大多数海水鱼幼鱼都有很强的趋光性。体长1~2寸的蓝圆鲹,能成群结队地趋光游泳,可是半斤以上的蓝圆鲹就很少来到光亮区。它们多半生活在光线微弱的底层水域。食物也由浮游动物转向海底介形类和硅藻类[23]。加利福尼亚的裂吻平鲉(Sebastesdiploproa)到了1龄开始迁徙,从表面迁徙到深海250~500 m,在垂直迁移过程中视网膜随光环境的变化而发生变化,仔鱼从高视觉分辨度发育到幼鱼高视觉灵敏度,剩下一点锥细胞仅接收捕食动物发出的生物光[24]。这些洄游迁徙往往伴引起鱼类视觉感光细胞形态、生理和生化的显著改变,比如迁移到的水生环境决定色素中维生素A1/A2比例[15]。因此鱼类光谱敏感性划分的界限不是绝对的,跟其生长阶段、生态位、视蛋白可塑性相关。
1.4 光谱对仔鱼开口期的重要性
鱼类的生活史是指精卵结合直至衰老死亡的生命周期过程,一般分为:胎胚、仔鱼、稚鱼、幼鱼、成鱼及衰老期[25]。在很多硬骨鱼类养殖中,仔鱼存活率高低与随后健康的稚鱼是养殖的瓶颈问题。所有仔鱼开口时期都是关键点,仔鱼是视觉摄食者,如果其开口阶段不能探知和抓到饵料,在卵黄吸收完后死去。对养殖者来说,得到仔鱼正确的光谱参数重要性不言而喻,因为对一个养殖户提高几个,甚至几十个百分点的仔鱼存活率不是一个小数目。2004年中山大学大规模人工孵化2900多万粒鞍带石斑鱼(Epinepheluslanceolatus)受精卵,结果成活5~8 cm的仔鱼34万多尾,成活率只有1.1%[26]。杨薇等[27]和魏开建[28]分析必须及时提供良好的开口饵料和光环境,才能保证仔鱼辨认运动中的浮游生物并成功地捕获与吞咽,得到足够的营养顺利地开口。Blanco~Vives等[29]记述塞内加尔的舌鰨孵化后11 d蓝光条件下仔鱼成活率为46.8%,而在红光条件下仅为27.8%。吴乐乐等[22]实验证明孵化后2天的大菱鲆仔鱼在绿光条件下的畸形率显著高于红光、橙光、蓝光、全光谱的畸形率。仔鱼通过背景光谱环境与食物反射光谱的对比,看见并捕捉食物。仔稚鱼开口及快速生长期,必须营造最适宜仔鱼的视觉系统光谱环境,以减少仔鱼的死亡率及后期品质的降级[30]。
1.5 鱼类光敏感性的数值
眼睛对不同波长光的相对敏感性随着亮度的刺激而发生适应状态变化称为普尔金耶现象(Purkinje phenomenon)[31]。对鱼类视网膜光谱敏感性数值的了解有助于设施养殖光环境的设计与远洋捕捞光诱技术中光源的选择。视网膜电流图是视觉电生理诊断学中应用最早的一种方法。杨雄里等[32]测定了蓝圆鲹(Decapterusmaruadsi)和鲐鱼(Pneumatophorusjaponicus)ERG b波的光谱敏感性, 表明两种鱼的暗视光谱敏感曲线峰值分别在490nm和480nm,当背景光强达到一定水平时, 两种鱼的光谱敏感曲线均发生普尔金耶现象,分别移至520nm和525nm。
20世纪70年代初,厦门大学、中科院上海生理研究所和上海水产研究所等科技工作者运用视网膜电流图方法和光-运动反应的行为取得一些淡水与海水硬骨鱼类光谱明、暗敏感峰值[33],尽管现在看来许多实验仪器不够精密, 如暗视仪、超微电极等。大多数硬骨鱼类都有辨别颜色的能力,当视网膜从暗视觉向明视觉过渡时,光谱敏感曲线的峰值向长波段移动[5,34]。
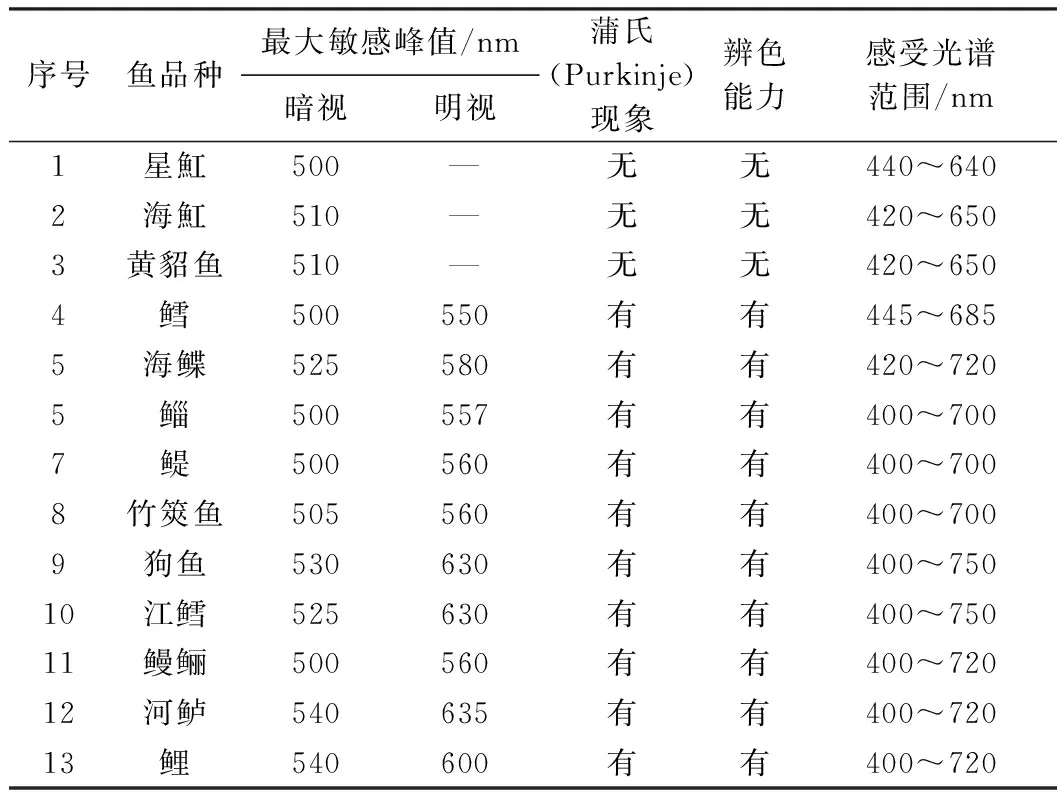
表1 部分海鱼光敏感数值表来源(参考文献[34])

图1 河鲈的光敏感曲线(来源:参考文献[34],根据印刷图重新绘制)Fig.1 Light sensitive curve of Perca Fluviatilis
目前对视网膜组织学及视色素的研究做得不多,更多研究聚焦研究珊瑚礁鱼类的视觉多样性以解析鱼类适应进化历程。水产动物对光的生理和行为响应是一个复杂的过程, 由其先天进化的视觉生理结构及其后天视觉可塑性共同决定。所以,只有利用现代精密的科研仪器、电生理学、生物化学及水产动物行为学等综合分析, 才能得到科学的结论。
2 虾蟹类视觉的光谱敏感性概述
2.1 虾蟹类视觉器官
和多数甲壳动物一样,虾蟹类的感光器官为复眼,复眼由多个小眼组合而成,成体复眼的外部形态为球形、半球形、心形、梨形及马鞍形,少数特化呈扁平形[35]。小眼由折光系统、感光系统和反光系统三部分组成。折光系统由角膜、角膜生成细胞和晶锥三部分组成,其中晶锥由晶状体细胞构成,或形成晶状体束。晶锥的作用可能类似于瞳孔,可以根据光线强弱通过形态变化控制光子进入,晶状体束能够将图像传导至感杆束。感光系统由小网膜细胞和感杆束组成,如刀额新对虾[36]、三疣梭子蟹[37]的小眼小网膜都是由4个远端小网膜细胞和7个近端小网膜细胞组成的“4+7”结构,感杆束由小网膜细胞伸出的微绒毛组成,包括远端感杆束和近端感杆束。反光系统由色素构成,包括远端色素、近端色素和反射色素[38]。总体来说,浮游生活种类、滤食性种类和生活在光线强的环境或白天活动的种类,复眼结构较简单;而游泳及底栖爬行种类、猎食性种类和生活在水域底层光线较暗或者昼伏夜出的种类,复眼结构较复杂,感光功能较强[36]。
2.2 虾蟹类视觉敏感性
关于经济虾蟹类的光谱敏感性研究较少,这可能与主要经济虾蟹类仍然主要采用池塘养殖模式,缺乏进行光调控的条件有关。甲壳动物中视觉敏感性研究较为深入的是口足类动物(虾蛄),因为其具有动物界最复杂的感光系统,有12种光感受器,视觉范围覆盖深紫外到远红外光(300~720 nm)。但并非所有的甲壳动物都具有如此精细的色彩分辨能力。Frank等[39]记录了12种栖息海洋中层的甲壳动物对光谱的电生理反应,发现所有测试物种的光谱敏感度峰值都出现在470 nm~500 nm之间。除了端足目动物(Phronimasedentaria)以外,其他11个物种的光谱敏感性与深度分布、迁徙行为无关。
早期研究认为蟹类复眼是一个单一的感受系统,缺乏颜色分辨能力[40]。但后期越来越多的证据都支持蟹类复眼具有颜色视觉。如Horch 等[41]发现招潮蟹(Ucathayeri)复眼中存在两种视觉色素,吸收峰值分别出现在430nm和500~540 nm。Premrajet等[42]在招潮蟹(Ucapugilator)的眼柄中鉴定了视蛋白编码基因(UpRh1、UpRh2和UpRh3),UpRh1和UpRh2氨基酸序列与其他节肢动物中长波敏感视蛋白相似,而UpRh3与其他节肢动物紫外敏感视蛋白相似。在行为学方面,雌性招潮蟹(Ucamjoebergi)偏爱黄色大螯的雄性[43],而雄性蓝蟹(Callinectessapidus)在交配中偏好红色指节的雌蟹[44]。雪蟹(Chionoecetesopilio)对蓝光和白光有显著偏好,布置蓝光的捕捉装置能显著提高雪蟹的渔获量[45]。但是与鱼类相比,针对虾蟹类视网膜电流图的研究仍然较少。
2.3 光谱对虾蟹类生长的影响
光谱对虾蟹生长和摄食具有显著性影响,且随着个体发育阶段不同而不同。例如蓝光延长了凡纳滨对虾幼体的发育时间[46],但是对于凡纳滨对虾成体,蓝光则显著增加了其抗氧化能力,并提升了存活和生长表现[47]。这可能与凡纳滨对虾不同生活史阶段栖息的水层不同有关,随着凡纳滨对虾的生活方式由浮游向底栖转变,适宜其生长的光谱也不断向短波光移动。但是另一方面,个体发育导致栖息地光谱环境的变化,并不能解释所有养殖水产动物的光谱偏好和生长表现,例如蓝光降低了中华绒螯仔蟹的摄食率与生长率[48],但是却对中华绒螯蟹幼体具有显著的诱集效应[49]。这表明与鱼类一样,虾蟹的视觉敏感光谱和选择偏好并不一定与生长最优的光谱一致,需要通过实验分别验证。
3 光环境设计应遵循的一般原则
由于众多品种的鱼、蟹及虾栖息环境的复杂性而进化出生活繁殖习性的多样性。有的鱼类,如日本鳗鲡,在淡水中生长,到繁殖期则要降海长距离洄游至大海深处产卵;而另一些鱼类,如大马哈鱼,生活大海中,繁殖期却要洄游到江河上游,甚至小溪中生殖[2]。所以要求水产养殖动物人工光照环境设计不能机械照搬适宜人眼的建筑照明设计,也不能套用家禽几个单一品种的光照设计模式。罗会明与郑微云[50]实验得出海上作业用黄橙光可使幼小鳗鲡聚集,而用紫色灯光诱捕成鳗可以提高渔获量。设计时宜遵循:(1)以鱼、蟹及虾的视觉器官的生理构造鱼视觉机能为基础依据。比如硬骨鱼类,需分析其视锥细胞与视杆细胞的结构、分布数量与视网膜明视觉与暗视觉光谱灵敏度,并参照其野生生存栖息水体的自然光环境,最终以科学验证的参数为准。(2)对不同生长发育期的水产动物,应考虑不同的光谱环境,比如一些品种的鱼在卵发育、仔鱼、稚鱼、成鱼期阶段需要的光环境时不同的。而不同的鱼在同一发育阶段也有其特殊性,如受精鱼卵在发育过程中的光照宜注意卵的类型,海水鱼多考虑浮性卵,淡水鱼多考虑沉性卵、漂浮卵[2]。(3)养殖池颜色选择在水产动物生活中非常重要[51],直接影响视觉器官识别并捕捉饵料,目前养殖户选择养殖池颜色带有很大的随意性,应考虑人工光辐照养殖池侧壁与底部被吸收后补色一体的效果。(4)宜用普尔金耶现象来确定鱼是否有辨别颜色的能力,如果养殖某种鱼的视网膜中只有视杆细胞,宜仅考虑光源的辐射强度。(5)设计时若采用白光需加色温及显色指数说明,目前白光LED发出的主波峰大部分是蓝光。若采用单一波长光谱需标明光源峰值的波长。若采用全光谱,注意LED光源的显色指数高及色温低不能代表该光源是全光谱光源[14],而是要比对与太阳光谱的相似程度。(6)若养殖水产动物品种不同生命周期需要不同的光谱,光源最好采用适宜波长混合灯珠,并用智能控制系统,以便在需要时变换光谱,因为在水面上安装更换灯具十分麻烦。
4 二个典型水产品种光照设计范例
4.1 大菱鲆养殖光谱设计
大菱鲆是世界公认的优质比目鱼之一。自1992年引进我国以来,大菱鲆产业经历了孵化与养殖技术攻关、产业技术熟化到目前年产8~10万吨的产业规模,占我国鲆鲽类总产量的80%以上,占世界大菱鲆总产量的75%左右。大菱鲆的发育过程经历变态,从水上层游泳生活转变为底栖生活,光照环境随之发生变化。光照作为重要的环境因子,对大菱鲆的胚胎发育,仔鱼、稚鱼和幼鱼的生长、代谢及非特异性免疫均有重要影响。
2.李贤团队证明从大菱鲆孵化后到变态前养殖阶段(约为孵化后1~20 d),应选择全光谱作为光源,光周期设定为24 L∶0 D,有助于其分布均匀、摄食活跃,减少应激反应,显著提高存活率[22]。
3.在大菱鲆开始变态及变态完成后,采用蓝光作为光源,有助于减少应激反应,提高特定生长率,促进视觉器官发育。在养殖期间,红光、橙光下大菱鲆幼鱼肌肉乳酸脱氢酶活力显著高于蓝、绿光,而血清乳酸含量差异不显著,表明红、橙光下大菱鲆幼鱼无氧代谢占的比重更大[21]。
4.2 皱纹盘鲍养殖光谱设计
皱纹盘鲍(Haliotisdiscushannai)是我国重要的海产经济贝类之一,2018年鲍的养殖产量达到16.32万吨,约占全球总产量的90%。作为昼伏夜出型生物的典型代表,鲍的运动和摄食行为多发生于夜间,而白天避光躲避于暗处。光照作为重要的环境因子之一,对成鲍、稚鲍的生长、代谢及幼虫的胚胎发育过程均具有重要影响。
高霄龙与刘鹰的研究发现在皱纹盘鲍的幼虫培育过程中,选择蓝、绿光且从担轮幼虫期起始光照可以提高幼体孵化率、变态率和存活率[52]。在成鲍和稚鲍的循环水养殖过程中,选择红、橙光作为光源,鲍从食物中获取的能量多,在粪便和排泄中损失的能量少,以致累积更多的能量用于生长[6]。但在蓝、绿光下时,稚鲍的耗氧率、排氨率升高,且随着鲍个体增大,其单位体重耗氧率、排氨率显著降低。机体通过糖酵解和糖异生作用来维持血糖平衡,但长期在此光质下必然对机体生长产生不利影响。
5 结论
光作为重要的环境因子,对水产动物整个生活史(从胚胎发育到性成熟)产生直接而广泛的影响[30]。在渔业生产中,无论是诱集捕捞,还是网箱与工厂化设施,人们已经意识到采用人工光照方式可以增加产量。由于水产动物视觉器官复杂多样性及作业现场光的次生影响能被其它的如温度、养殖池颜色及饲料等因素混淆,找到正确的光照环境参数不是一件容易的事情。因此,研究水产动物光敏感性对合理选择LED光源光谱分布,建立生物学的适宜光照调控策略,促进渔业产业持续、健康发展具有现实的指导意义。
致谢:本文得到大连海洋大学刘鹰教授指导,皱纹盘鲍光照设计范例由厦门大学海洋与地球学院高霄龙博士提供,在此表示感谢。
猜你喜欢
渔业研究(2022年5期)2022-10-28 01:56:14
海洋信息技术与应用(2022年1期)2022-06-05 07:38:24
食品工业(2021年9期)2021-04-16 09:49:47
中国水产(2019年3期)2019-03-25 09:27:20
探索科学(2017年5期)2017-06-21 21:16:16
金色少年(奇趣科普)(2016年8期)2016-09-21 02:04:58
浙江农业学报(2016年7期)2016-06-15 20:30:11
广东海洋大学学报(2015年4期)2016-01-13 08:39:40
集美大学学报(自然科学版)(2015年4期)2015-02-28 01:13:34
小星星·阅读100分(低年级)(2014年5期)2014-06-24 00:42:45
