
PSMB5蛋白对牛骨骼肌卫星细胞增殖与成肌分化的影响
2020-11-16 02:49来裕婷朱菲菲王轶敏郭宏张林林李新郭益文丁向彬
中国农业科学 2020年20期
来裕婷,朱菲菲,王轶敏,郭宏,张林林,李新,郭益文,丁向彬
(天津农学院动物科学与动物医学学院/天津市农业动物繁育与健康养殖重点实验室,天津 300384)
0 引言
【研究意义】 肌肉是动物躯体的重要组成部分,骨骼肌大约占体重的40%,骨骼肌在调节动物新陈代谢、机体运动、能量储存和健康等方面至关重要,是机体功能正常运转的必要组分。骨骼肌卫星细胞作为肌源性的干细胞,在肌肉发育分化和肌损伤修复中都发挥了重要的作用。泛素-蛋白酶体系统(Ubiquitinroteasome system, UPS)是真核生物降解蛋白的主要途径。泛素(ubiquitin, Ub)先标记要降解的蛋白质,然后由蛋白酶体识别和降解[1]。泛素蛋白酶体介导的蛋白降解由泛素(Ub)、泛素活化酶(ubiquitin-activating enzyme, E1)、泛素结合酶(ubiquitin-conjugating enzyme, E2)、泛素-蛋白连接酶(ubiquitin-protein ligating enzyme,E3)、去泛素化酶(deubiquitinating enzymes,DUBs)、26S蛋白酶体及其底物(蛋白质)构成[2-3]。泛素-蛋白酶体对底物的降解过程主要包括泛素化、蛋白酶体降解、去泛素化3个过程。由于底物蛋白在细胞内广泛存在,泛素化参与调节细胞周期进程、细胞增殖与分化、以及信号传导等多种生物学过程[4]。在肌肉发育分化过程中,泛素-蛋白酶体系统可能也发挥了重要的调节作用,具体调控机制亟需进行深入研究。【前人研究进展】26S蛋白酶体是降解泛素化底物的一个ATP依赖型蛋白水解复合体,由20S核心蛋白酶体和19S调节颗粒构成[5-7]。19S调节颗粒可以识别泛素化的蛋白,20S是泛素-蛋白酶体系统的中枢蛋白水解结构[8-9]。20S是一个圆筒形结构,由α亚基和β亚基构成的4个环层叠组成,内侧的2个β环具有蛋白酶催化水解活性[10],β1、β2和β5为活性亚基,具有特异性肽切割位点,是整个泛素-蛋白酶体通路降解蛋白的关键位点[11-12]。其中,蛋白酶体β5亚基(PSMB5)具有糜蛋白酶样活性[13]。在肌肉中,泛素-蛋白酶体系统与肌原纤维蛋白的降解密切相关[14-15]。有研究表明,过表达PSMB5可增强人骨髓间充质干细胞20S蛋白酶体活性,提高细胞增殖能力[16]。也有研究表明,PSMB5过表达能够促进人骨髓间充质干细胞向神经元样细胞分化[17]。还有研究表明免疫蛋白酶体 β5i 亚基可通过升高钙调神经磷酸酶的蛋白水平和活性,促进 DOCA 盐诱导的心肌肥厚[18]。沉默 PSMB5基因可降低神经干细胞蛋白酶体活性抑制新生小鼠神经干细胞的增殖分化能力[19]。也有研究表明在心脏肥大的动物模型中,心脏蛋白酶体亚基的蛋白表达水平与心肌肥厚呈正相关,可能是由于蛋白酶体促进了心脏肥大信号通路中相关蛋白的表达[20]。【本研究切入点】上述研究提示蛋白酶体亚基 PSMB5可能在细胞分化和肌肉发育过程中发挥了重要的调控作用,但PSMB5对牛肌肉发育分化是否具有调控作用还不清楚。【拟解决的关键问题】本研究利用牛骨骼肌卫星细胞体外诱导成肌分化模型,模拟牛骨骼肌生长发育过程,通过改变牛骨骼肌卫星细胞中PSMB5基因的表达水平,研究 PSMB5蛋白对牛骨骼肌卫星细胞体外增殖和成肌分化的影响。
1 材料与方法
1.1 细胞
牛骨骼肌卫星细胞由天津市农业动物繁育与健康养殖重点实验室分离冻存。
1.2 试验进行时间及地点
试验于2019年1—9月在天津市农业动物繁育与健康养殖重点实验室完成。
1.3 主要仪器
二氧化碳培养箱(SANYO);荧光倒置显微镜(Leica);通用酶标仪(Biotek)电泳仪、转膜仪、高灵敏化学发光成像分析系统(Bio-Rad)等。
1.4 主要试剂
DMEM;FBS(Fetal Bovine Serum),HS(Horse Serum),Opti-MEM,0.25%胰蛋白酶购自美国Gibco公司;si-RNA和EdU细胞增殖检测试剂盒购自广州锐博公司;RIPA裂解液,电泳液,电转液,TBS和发光液购自北京索莱宝公司;Protease inhibitor cocktail购自美国SIGMA公司; BCA试剂盒购自北京康为公司;LipofectamineTM3000 Reagent和 proteasome 20S Antibody抗体购自赛默飞公司;MyHC一抗购自DSHB公司;GAPDH一抗,羊抗鼠二抗和羊抗兔二抗购自北京中杉金桥公司;pcDNA3.1购自武汉淼灵生物科技有限公司;RNA快速提取试剂盒购自艾德莱生物科技有限公司;反转录试剂盒购自 Takara biotechnology Corporation;All-in-OneTMqPCR Mix,购自GeneCopoeia。
2 方法
2.1 牛骨骼肌卫星细胞的复苏、传代及诱导分化
牛骨骼肌卫星细胞体外诱导成肌分化模型的建立参考天津市农业动物繁育与健康养殖重点实验室已建立的方法进行[21]。细胞在37 ℃水浴中复苏,用1 mL完全培养基(含20% FBS的DMEM)中和,室温,1 000 r/min 离心 10 min,移除上清,加入适量的完全培养基重悬,接种于60 mm的培养皿中。待细胞融合度达到 80%时传代,用 37 ℃预热的 PBS清洗细胞两次,加入 1 mL 0.25%胰酶室温消化 2 min,加入1 mL完全培养基终止消化,室温1 000 r/min离心5 min,移除上清,加入适量的完全培养基重悬,接种于细胞培养板中。待细胞汇合度达到80%,加入分化培养基(含2% HS的DMEM)进行体外诱导分化。
2.2 试验设计及处理
牛骨骼肌卫星细胞分化前后 PSMB5表达量的检测:将牛骨骼肌卫星细胞传代后接种于24孔板,在完全培养基中培养24 h,诱导分化培养72 h,利用RNA快速提取试剂盒分别提取增殖期和分化期牛骨骼肌卫星细胞总 RNA,qRT-PCR检测牛骨骼肌卫星细胞分化前后PSMB5的mRNA表达差异,Western blotting法检测 PSMB5在牛骨骼肌卫星细胞分化前后蛋白表达水平的差异。
PSMB5的干扰及过表达效果检测:设计合成PSMB5 小干扰 RNA si-RNA-PSMB5 (si-PSMB5,序列为:AGAGGAGCCAAGAATTGAA);构建PMSB5的表达载体 pcDNA3.1-PSMB5(pcDNA-PSMB5)。将牛骨骼肌卫星细胞在24孔培养板内进行增殖培养,当细胞融合度达 80%时,按生产商家说明书利用Lipofectamine 3000转染已构建的si-PSMB5及过表达载体pcDNA-PSMB5,以si-NC及空质粒pCDNA3.1(pcDNA-NC)转染细胞作为对照。qRT-PCR及Western blotting法检测转染处理后48 h的过表达和干扰效果。
PSMB5对牛骨骼肌卫星细胞增殖的影响:转染后24 h,按 Cell-Light EdU Apollo In Vitro Imaging Kit试剂盒说明书检测牛骨骼肌卫星细胞增殖期细胞数。
PSMB5对牛骨骼肌卫星细胞成肌分化的影响:光学显微镜观察干扰和过表达 PSMB5后诱导分化 72h牛骨骼肌卫星细胞肌管形成状态,Western blotting检测分化标记因子MyHC的蛋白水平表达。
2.3 qRT-PCR检测PSMB5的基因表达水平
取 5 μL 总 RNA 利用 All-in-OneTMFirst-Strand cDNA Synthesis Kit试剂盒将RNA反转录为cDNA,再采用qRT-PCR检测PSMB5基因的表达水平。
qRT-PCR 反应体系:10 μmol·L-1上游引物、10 μmol·L-1下游引物、2.0 μL 5 倍稀释 cDNA、10 μL 2×All-in-OneTMqPCR Mix,RNase-free water 将体系补至20 μL。qRT-PCR反应引物信息如表1所示。
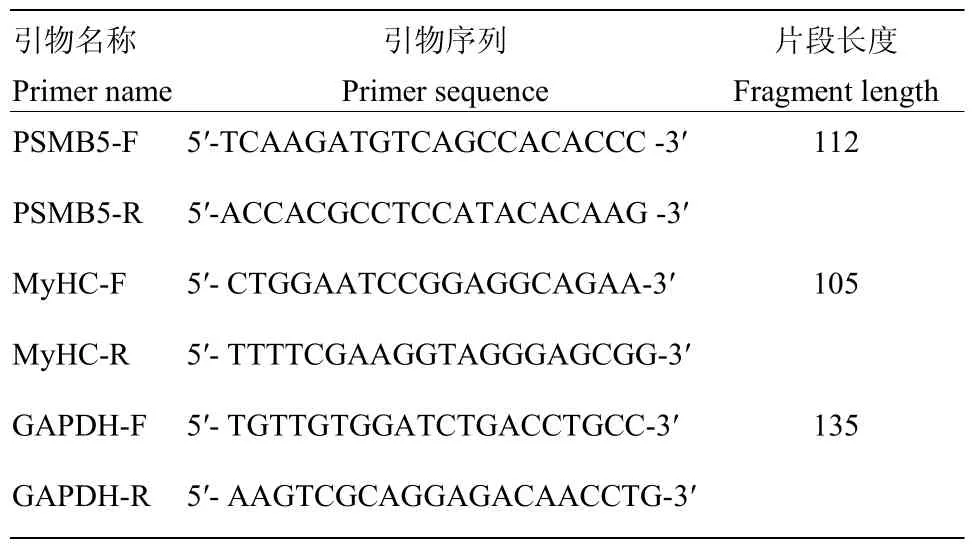
表1 qRT-PCR引物信息Table 1 The primer information of qRT-PCR
2.4 Western blotting检测 PSMB5和牛骨骼肌卫星细胞分化标记基因蛋白表达水平
用蛋白裂解液裂解6孔板中增殖期细胞或分化期肌管细胞,收集蛋白,用 BCA法测定蛋白浓度,再通过 Western blotting检测 PSMB5和分化标志基因MyHC的蛋白的表达量,具体步骤参考本实验室已建立的方法进行[22]。
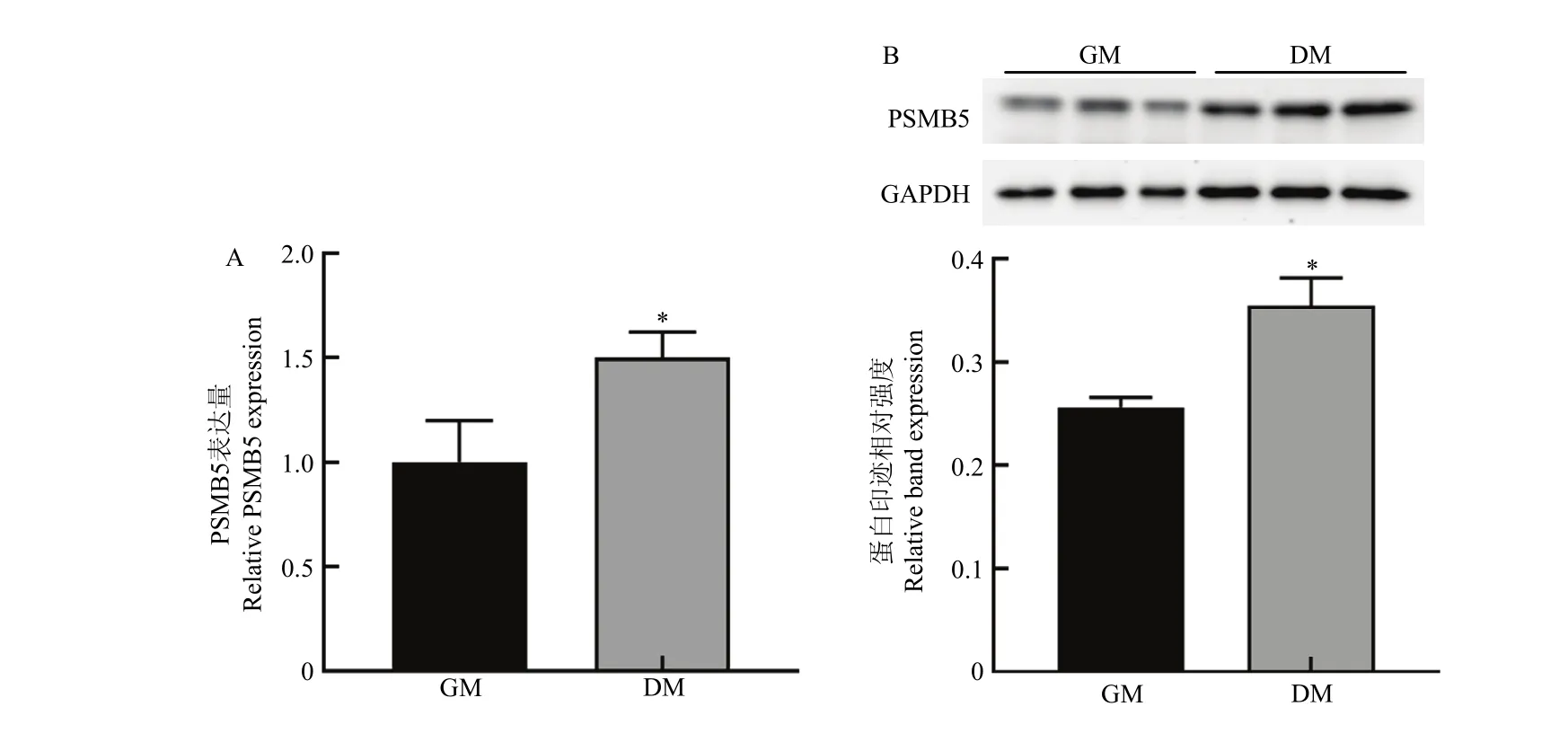
图1 牛骨骼肌卫星细胞分化前后PSMB5表达量的检测Fig. 1 Expression of PSMB5 in proliferation and differentiation of bovine skeletal muscle satellite cells
2.5 EdU法检测牛骨骼肌卫星细胞增殖状态
将牛骨骼肌卫星细胞接种在 96孔板中,转染后24 h,按 Cell-Light EdU Apollo In Vitro Imaging Kit试剂盒说明书检测各处理细胞的增殖期细胞数。每组试验均设置3个生物学重复,每个生物学重复至少采集3个技术重复或视野。EdU增殖效率=EdU阳性细胞数/Hoechst 33342标记细胞数,分别记数3个200×视野的细胞总数及EdU阳性细胞数。
2.6 统计分析
每组试验均设置3个生物学重复,所有数据均以“平均数±标准差”表示。EdU结果采用χ2检验进行分析;qRT-PCR结果按 2-ΔΔCt法计算,采用t检验进行差异显著性分析,以GAPDH基因作为内参基因对检测的目的基因进行归一化。“*”表示差异显著(P<0.05),“**”表示差异极显著(P<0.01)。
3 结果
3.1 牛骨骼肌卫星细胞分化前后PSMB5表达量的检测
将牛骨骼肌卫星细胞传代后接种于24孔板,在增殖培养基中培养24 h,诱导分化培养72 h,qRT-PCR和 Western blotting检测牛骨骼肌卫星细胞分化前后 PSMB5的表达差异,(图1)。可知,PSMB5在牛骨骼肌卫星细胞分化前后表达水平存在显著差异,诱导分化后牛骨骼肌卫星细胞中PSMB5的mRNA和蛋白表达量均显著高于增殖期(P<0.05)。结果提示 PSMB5可能对牛骨骼肌卫星细胞的成肌分化有一定的调控作用。
3.2 PSMB5干扰及过表达效果检测
当24孔培养板内增殖期细胞融合度达80%时,转染小干扰 RNA si-PSMB5和 PSMB5过表达质粒pcDNA-PSMB5,转染处理后 48 h后,qRT-PCR及Western blotting检测细胞中PSMB5的表达水平变化。结果如图2和图3所示,转染si-PSMB5后,PMSB5的mRNA表达水平(图2-A)及蛋白表达水平(图2-B)显著低于对照组(P<0.05);转染 pcDNA-PSMB5过表达质粒后,PSMB5的mRNA水平(图3-A)及蛋白表达水平(图3-B)显著高于对照组(P<0.01)。结果表明小干扰RNA以及构建的PSMB5过表达质粒分别对牛骨骼肌卫星细胞具有明确的干扰和过表达效果。
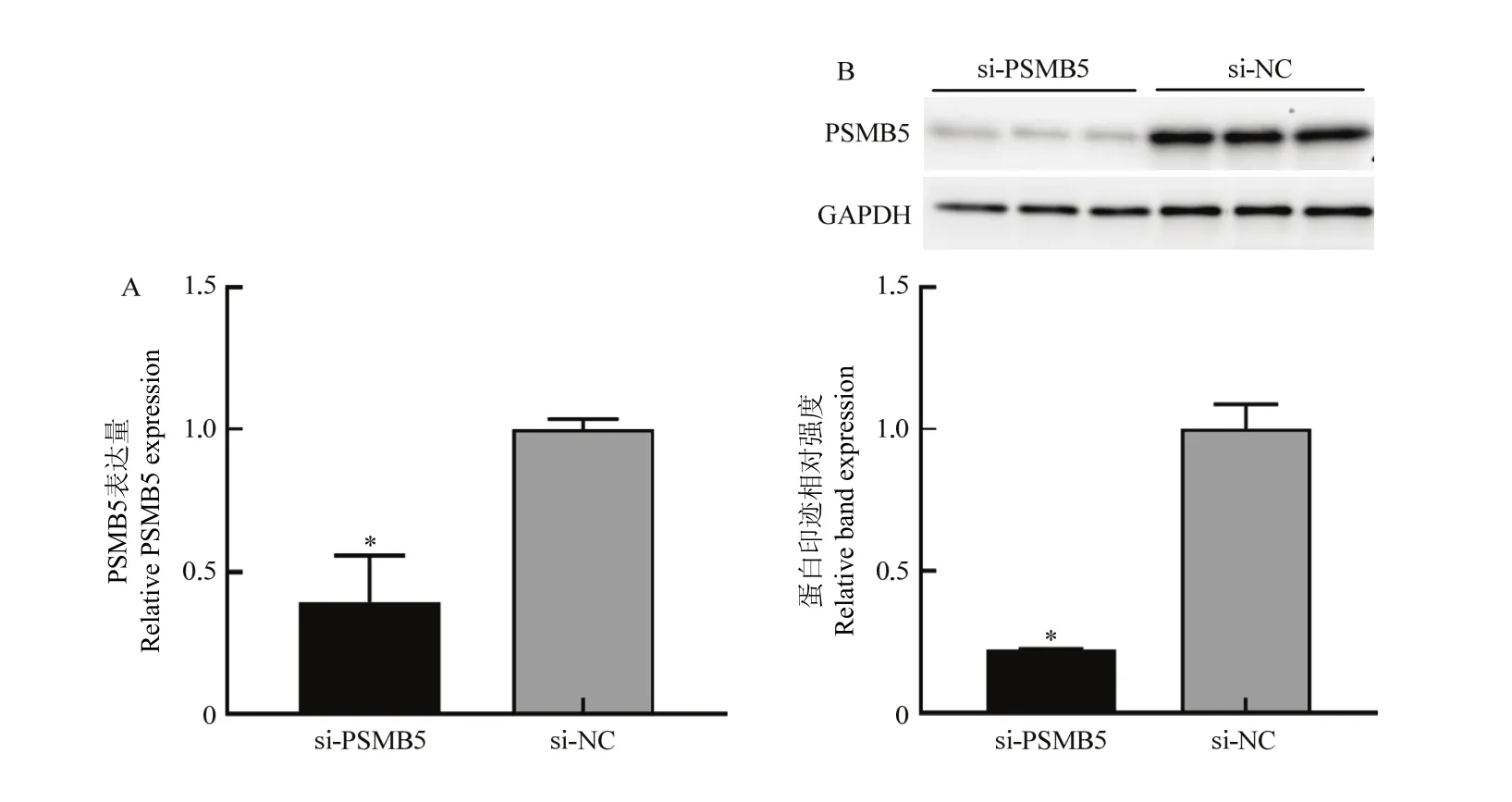
图2 siRNA的干扰效果检测Fig. 2 Detection of interference effects of si-PSMB5
3.3 PSMB5对牛骨骼肌卫星细胞增殖的影响
将 96孔板中细胞转染 si-PSMB5和 pcDNASMB5,转染处理24 h后,采用EdU法检测牛骨骼肌卫星细胞的增殖状态。结果如图4所示,干扰或过表达PSMB5后,牛骨骼肌卫星细胞EdU阳性细胞率与对照组相比差异均不显著(P>0.05)。结果表明PSMB5对牛骨骼肌卫星细胞的增殖没有明显的影响。
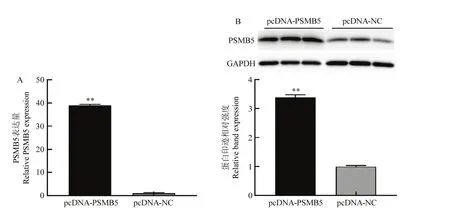
图3 PSMB5质粒载体的过表达效果检测Fig. 3 Detection of pcDNA-PSMB5 overexpression effect
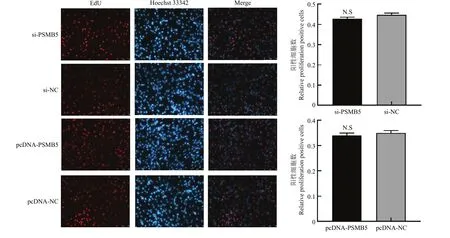
图4 EdU法检测干扰及过表达PSMB5对牛骨骼肌卫星细胞增殖的影响Fig. 4 EdU detects proliferating cells of bovine skeletal muscle satellite cells after knockdown and overexpression of PSMB5
3.4 PSMB5对牛骨骼肌卫星细胞成肌分化的影响
干扰或过表达PSMB5后,将牛骨骼肌卫星细胞诱导分化 72 h,然后采用光学显微镜观察牛骨骼肌卫星细胞肌管形成状态,Western blotting 检测分化72 h后PSMB5以及分化标记因子MyHC的蛋白水平表达。如图5所示,干扰PSMB5后,从细胞形态图来看,试验组形成的肌管数量明显少于对照组(图5-A);Western blotting检测结果显示,干扰PSMB5表达后,分化标志因子MyHC的蛋白水平显著降低(P<0.01)(图5-C);诱导分化72 h时,PSMB5蛋白水平仍显著低于对照组(P<0.01)(图5-D)。过表达PSMB5后,试验组中形成的肌管数量明显多于对照组(图5-B);Western blotting检测结果显示,过表达PSMB5后,分化标志因子MyHC的蛋白表达水平显著高于对照组(P<0.01)(图5-E);诱导分化72 h,试验组PSMB5的蛋白表达水平仍高于对照组但差异不显著(P>0.05)(图5-F)。研究结果表明干扰 PSMB5的表达会抑制牛骨骼肌卫星细胞体外成肌分化进程,而过表达 PSMB5可以促进牛骨骼肌卫星细胞体外成肌分化进程,PSMB5对牛骨骼肌卫星细胞的成肌分化过程具有显著的调控作用。
4 讨论
骨骼肌发育过程极其复杂,主要包括体节细胞增殖分化、成肌细胞增殖分化、肌管成熟以及肌纤维形成等环节。骨骼肌卫星细胞是骨骼肌中具有分化增殖潜能的肌源性干细胞,通常以静息状态存在于肌纤维肌膜与基底膜之间,在一定的条件下可以被激活,发生增殖和分化。卫星细胞在动物出生后肌肉的生长发育和再生过程中发挥着十分重要的作用[23-24]。肌卫星细胞的体外成肌分化过程很好地模拟了体内的发育过程,成为研究细胞分化和肌肉发育发生机制的良好细胞模型。泛素-蛋白酶体途径在细胞内普遍存在,从细胞膜、细胞质到细胞核等处的蛋白质都受到泛素化酶和蛋白酶体的监控[25]。有研究表明,对于处于活跃增殖期的细胞如血管平滑肌,泛素-蛋白酶体主要通过NF-κB抑制细胞凋亡,引起血管平滑肌细胞发生增殖性病变;对于已经分化并且处于非分裂期的细胞,如在不稳定的动脉粥样硬化斑块中的细胞,泛素-蛋白酶体抑制细胞凋亡的作用明显减弱[26]。此外,蛋白酶体可以降解凋亡相关蛋白,抑制蛋白酶体活性,可以促使 p53、凋亡蛋白Bax和蛋白激酶C(PKC)等集聚,进而促使心肌细胞凋亡[27]。在小鼠骨骼肌卫星细胞的再生过程中,蛋白酶体的活性上升[28]。另外,泛素-蛋白酶体系统在肌肉萎缩和神经损伤变性过程中都发挥了重要作用。应用蛋白酶体抑制剂MG-132能抑制离体神经肌细胞蛋白降解,延缓离体神经轴突变性的发生[29]。综合以上研究发现,泛素-蛋白酶体系统在细胞增殖、分化中起着非常重要的作用。PSMB5为蛋白酶体中具有水解活性的亚基,在泛素-蛋白酶体中发挥着重要作用,它可能在细胞分化和肌肉发育过程中发挥了重要的调控作用。目前,PSMB5在牛骨骼肌卫星细胞增殖分化中的表达模式和调控作用还不清楚,因此,笔者检测了牛肌卫星细胞增殖分化过程中PSMB5的mRNA和蛋白表达水平的变化,检测结果发现 PSMB5在牛骨骼肌卫星细胞分化前后表达水平存在显著差异,PSMB5在牛骨骼肌卫星细胞分化期的表达水平较高,推测其对牛骨骼肌卫星细胞的增殖和分化过程可能具有一定的影响。
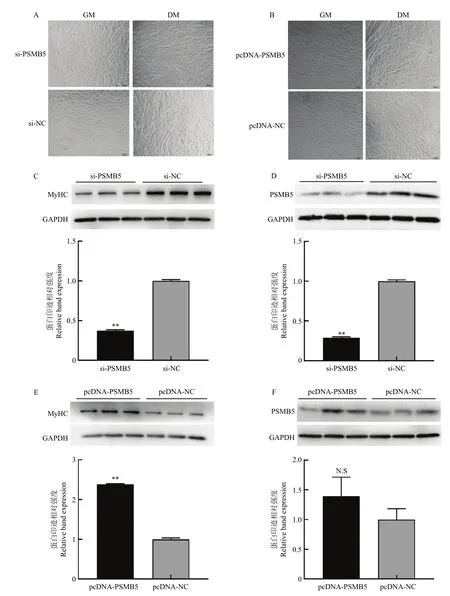
图5 干扰和过表达PSMB5对牛骨骼肌卫星细胞分化的影响Fig. 5 Effects of interference and overexpression of PSMB5 on the differentiation of bovine skeletal muscle satellite cells
有研究报道,选择性敲除小鼠肌卫星细胞特异性19S蛋白酶体调节亚基(Rpt3)基因,会阻碍小鼠体内肌再生过程,同时还会抑制肌再生过程中肌卫星细胞的增殖与分化[30]。PSMB5基因沉默可降低神经干细胞蛋白酶体活性抑制小鼠神经干细胞的增殖分化能力[19]。PSMB5过表达能够促进人骨髓间充质干细胞向神经元样细胞分化[17]。在本研究中,通过模拟体内肌卫星细胞增殖分化过程,成功建立体外牛肌卫星细胞增殖分化模型,设计si-RNA抑制PSMB5的表达,干扰PSMB5后发现,细胞分化能力减弱,肌管数量明显减少,同时牛骨骼肌卫星细胞分化标志基因 MyHC的蛋白表达量下调,试验结果表明,干扰 PSMB5表达会抑制牛肌卫星细胞的分化;构建 PSMB5过表达载体,转染细胞后发现,过表达 PSMB5的牛骨骼肌卫星细胞分化能力增强,肌管数量增多,且MyHC蛋白表达量上调,说明过表达 PSMB5后能够促进牛骨骼肌卫星细胞的成肌分化。本研究结果表明,PSMB5对牛骨骼肌卫星细胞的成肌分化过程具有显著的调控作用,与骨髓间充质干细胞和小鼠神经干细胞上研究类似,PSMB5在牛骨骼肌卫星细胞上表现出了对细胞分化的调控作用,但 PSMB5在牛骨骼肌卫星细胞上没有表现出在骨髓间充质干细胞和神经干细胞上类似的对增殖的调控作用,可能是其在不同细胞类型中发挥作用的途径不一致,具体的调控作用机制还有待进一步研究。
5 结论
本研究发现PSMB5蛋白对牛骨骼肌卫星细胞的成肌分化过程具有显著的调控作用。干扰PSMB5表达可以抑制牛骨骼肌卫星细胞的成肌分化进程,而过表达 PSMB5可以促进牛骨骼肌卫星细胞体外成肌分化进程。研究探明了PSMB5蛋白对牛骨骼肌卫星细胞增殖和成肌分化的具体调控作用,为进一步开展 PSMB5在牛成肌分化中的调控机制研究奠定了基础。
猜你喜欢
中国生物化学与分子生物学报(2022年4期)2022-09-07
生物化学与生物物理进展(2022年8期)2022-08-20
体育科技文献通报(2022年3期)2022-05-23
清华金融评论(2022年4期)2022-04-13
医学综述(2021年16期)2021-12-01
心血管病学进展(2021年8期)2021-09-13
国际放射医学核医学杂志(2021年10期)2021-02-28
湖北农业科学(2020年24期)2021-01-21
房地产导刊(2020年7期)2020-08-24
中国外汇(2019年14期)2019-10-14
