
不同沙化阶段高寒草甸植物群落与表土环境因子的关系
2020-11-13 09:28王新源马仲武王小军程小云
生态学报 2020年19期
王新源,马仲武,王小军,陈 萍,唐 霞,连 杰,张 琴,程小云,刘 健,曲 浩
1 甘肃省生态环境监测监督管理局, 兰州 730020 2 甘肃省林业调查规划院,兰州 730020 3 中国科学院西北生态环境资源研究院,兰州 730000 4 中国科学院西北生态环境资源研究院乌拉特荒漠草原研究站, 兰州 730000
荒漠化是发生于干旱、半干旱和亚湿润干旱区,在自然因素和人类活动共同影响下形成的土地退化过程,其中以风为主导营力导致的,形成类似于沙漠景观的退化现象称作沙漠化[1-2]。沙漠化对生态环境的影响是动态的、负面的,常伴随着土地生产潜力下降、土壤理化性质衰退及植被的逆向演替[3-4],长期沙化将导致原生生态系统(多数为草地生态系统或绿洲生态系统)退化为沙漠生态系统,最终引发严重的生态危机。事实上,尽管风是沙漠化景观的主要塑造者,但其产生的直接诱因却是水资源的短缺,因此,沙漠化通常发生于水分受限生态系统[5-6]。
玛曲县位于青藏高原东北缘,是黄河上游重要的水源涵养区,发育其中的高寒草甸是阻碍高原风沙东移的关键生态屏障[7-8],同时也为区域内各族人民进行牧业生产提供了物源基础。但自20世纪70年代以来,伴随着全球气候变化和人类活动的加剧,玛曲县沙漠化问题趋于严重,集中表现为物种多样性减少、水资源分配格局改变、土地生产力及生产潜力下降、湿地萎缩、生态服务功能衰退等方面[8-9]。沙漠化导致玛曲县高寒草甸生态系统脆弱、敏感,极大的削弱了其生态承载力,严重制约着该区域经济社会的持续健康发展。
不同于多数沙漠生态系统,玛曲县高寒草甸沙漠化区域存在一定的地带特性,主要包括高海拔、水资源相对丰富和热量限制几个方面[10]。这种高寒草甸生态系统产生的沙漠化现象引起了诸多学者的重视,其研究内容主要集中于沙漠化土地面积及程度的时空变化[11-14]、沙漠化产生机理及风沙地貌景观格局的形成[15-16]、土地利用/覆被变化规律[17-18]和高寒草甸退化成因分析[19]等方面。然而,对于不同沙漠化生境梯度下植物群落与表土环境因子间相互关系及变化规律的研究还比较薄弱。目前,植被的恢复重建与高寒草甸生态系统功能的维持与改善仍是玛曲县沙漠化治理的主要方向,而草甸植被的生态响应区主要集中于土壤表层[20-21],因此,明晰不同沙漠化阶段植物群落与表层土壤环境因子间的内在联系,有助于明确沙漠化生境演替的生态学规律,使防治工作有的放矢。鉴于此,研究选取2018年玛曲县高清遥感影像数据,通过人工解译与实地验证,并参考相关资料[22], 将玛曲沙漠化草甸划分为潜在、轻度、中度、重度和极重度等不同的演替阶段,在此基础上,系统分析各演替阶段表层土壤环境因子与植物群落特征的动态变化,揭示沙漠化各阶段表土环境因子与植物群落结构、格局及演替的内在关联,旨在为玛曲高寒草甸沙漠化防治及生态服务功能的维持与恢复提供理论依据。
1 研究区概况
玛曲县地处甘肃省甘南藏族自治州西南部,地理坐标为33°06′30″—34°30′15″ N, 100°45′45″—102°29′00″ E,西邻青海省, 东南接四川省, 属于青藏高原高寒生态区。海拔3300—4806 m, 地势由西北向东南逐步降低,地貌类型自西向东依次为西北部高山区、中南部丘陵区及东部河岸阶地。气候方面,玛曲县冬春季漫长,寒冷多风;夏秋季短暂,多雨湿润。年均气温1.2 ℃,最冷月(1月)平均温度-8.7 ℃,最热月(7月)平均温度11.3 ℃,年均降水量615 mm,年均蒸发量1352.4 mm;年均风速2.5 m/s,最大风速36 m/s,全年大风日数77.1 d (8级以上)。自然植被类型以高寒草甸为主,镶嵌分布有高寒草原、高寒灌丛及沙化草甸等,植物种类以草本为主,包括青藏苔草(Carexmoorcroftii)、赖草(Leymussecalinus)、垂穗披碱草(Elymusnutans)、高山嵩草(Kobresiapygmaea)、二裂委陵菜 (Potentillabifurca)、乳白香青(Anaphalislacteal)、防风(Saposhnikoviadivaricata)及高山早熟禾(Poakoelzii)等[7,11,20]。
2 数据与方法
2.1 遥感影像与沙漠化等级划分
遥感数据源来自于2018年高分二号(GF-2)的全色彩高清影像,其空间分辨率达到亚米级,极大的提高了玛曲县沙漠化的监测精度。采用HCS(Hyperspherical Color Space)法对影像进行多波段融合,HCS法具有较强的光谱保真度与空间细节表达能力,可最大程度的保留原始影像的光谱信息[23]。通过二次多项式重采样法进行几何精校正,以消除挤压、扭曲、位移等影响解译精度的几何畸变,综合两类方法,将获得高对比度、高保真与高空间解析度的影像数据,这将有助于沙漠化信息的准确识别。之后,根据沙漠化土地空间分布格局与光谱景观特征[11],结合《第六次全国荒漠化和沙化监测技术规定》[22],同时参考现地相关调查因子(植被、沙化景观等),将玛曲县高寒草甸沙漠化区域划分为潜在(未沙化)、轻度、中度、重度和极重度5个演替阶段。
2.2 材料与方法
实地工作开展于2019年8月下旬至9月上旬,该时段属于玛曲县植被生长旺盛期,利于植物与表土环境指标的收集。根据玛曲沙化高寒草甸的分布情况,选择欧拉乡黄河南侧阶地、曼日玛乡黄河西侧阶地、曼日玛乡黄河北部阶地等3个立地条件相近的区域作为调查区,面积分别为5.7、5.5、6.9 hm2。在调查区内,根据草甸沙化程度设置调查样地,样地面积为20 m×20 m。其中,欧拉乡黄河南侧阶地调查区内包含样地10个(潜在、轻度、中度、重度和极重度各2个),曼日玛乡黄河西侧阶地调查区内设置样地12个(潜在、轻度和极重度各2个,中度和重度3个),曼日玛乡黄河北部阶地调查区内样地数为15个 (潜在、轻度、中度、重度和极重度各3个),共计37个样地(表 1)。之后,于每个样地中依据五点法[24]设置5个1 m×1 m的小样方,测度样方内植物组成、丰富度、多度、盖度及株高等生态学指标。同时在每个样方内,采用美国光谱公司TDR-350土壤三参数速测仪,度量表层(10 cm)土壤温度、含水量(体积)和电导率。此外,本试验样方内表土环境因子的度量时间选定在10:00—11∶00时,且距前次降水事件保持至少72小时的时间梯度,以最大限度降低时间与天气因素引起的干扰。
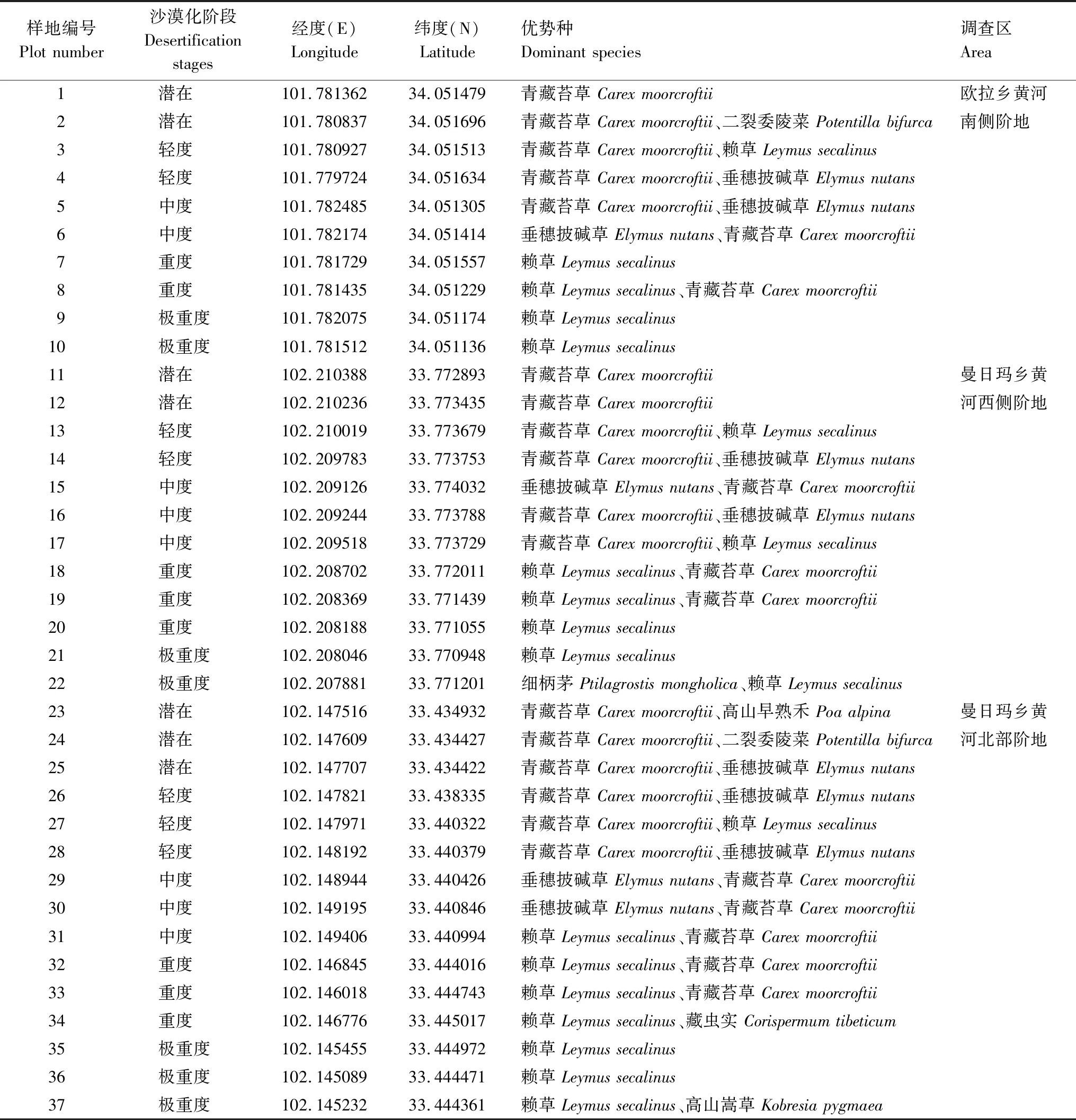
表1 样地分布与类型Table 1 Distribution and types of plots
2.3 数据处理与分析
应用Shannon-Wiener 多样性指数、Simpson 生态优势度指数及Pielou 均匀度指数来量度群落的物种多样性水平[25]
(1) 物种丰富度
R=S
式中,S为样方内物种数。
(2) 重要值
Ni=(相对盖度+相对株高+相对多度)/3
(3) Shannon-Wiener 多样性指数
式中,Ni是样方中第i种植物的重要值;N为样方所有植物重要值之和。下文中简称为Shannon-Wiener指数
(4) Simpson 生态优势度指数
Simpson 生态优势度指数(下文简称为Simpson指数)反映了群落优势种的变化情况,指数越大,物种间优势度差别越小,群落多样性水平越高。
(5) Pielou 均匀度指数
下文中简称为Pielou指数
采用EXCEL 2016和SPSS 20.0 进行数据分析,多样性指数通过R 3.5.2中“vegan”程序包diversity函数获得,应用单因素方差分析(one-way ANOVA)对不同沙化生境中植物群落及环境指标进行差异显著性分析(α=0.05),多重比较采取最小显著性差异法(LSD),所有数值均以平均值±标准差的方式表示。使用Canoco 5.0软件定量分析表土环境因子对植物群落组成与特征的影响。首先对数据进行除趋势对应分析(Detrended correspondence analysis, DCA), 获取的排序轴梯度长度(The length of the gradient axis, LGA)能够表征群落指标的变化程度。当 LGA<3时,数据适用于线性模型;LGA>4时,宜采用单峰模型;LGA介于3和4之间时,两种模型均适宜[4]。本研究不同沙化阶段中物种排序轴梯度最大值为3.9,且在区域时空尺度上,单峰模型擅长描述群落结构随沙化演替发生质变的现象(物种随生境变化出现或消失)[26],对此,应选用属于单峰模型的典范对应分析(Canonical correspondence analysis, CCA)。
3 结果与分析
3.1 不同沙化阶段高寒草甸植物群落组成
沙漠化阶段不同,物种的组成与分布亦不同。表2显示了高寒草甸沙漠化各阶段群落的组成结构与重要值。潜在沙化阶段共出现植物22种,主要优势种分别是青藏苔草、垂穗披碱草、二裂委陵菜和赖草,占群落总重要值的比例为43.97%。轻度沙化阶段植物种数较潜在阶段略微上升,为24种,其中垂穗披碱草、赖草和青藏苔草是优势种,重要值占比为44.07%。中度阶段,群落内有19种植物,优势种依次为青藏苔草、垂穗披碱草和狼毒(Euphorbiafischeriana),重要值占比为54.15%。重度阶段,有16种植物,优势种是青藏苔草和赖草,重要值占比52.26%。极重度阶段,植物种类锐减至8种,其中赖草占绝对优势地位,单一物种重要值占比达61.52%。综合可得,伴随沙化的发展,垂穗披碱草等优势种逐步衰退,狼毒等毒草侵入群落并发挥重要影响,赖草在整个沙化生境演替中均占据优势地位、表明其具有较宽广的生态位与抗干扰能力,是玛曲沙化草甸构建群落的关键物种。
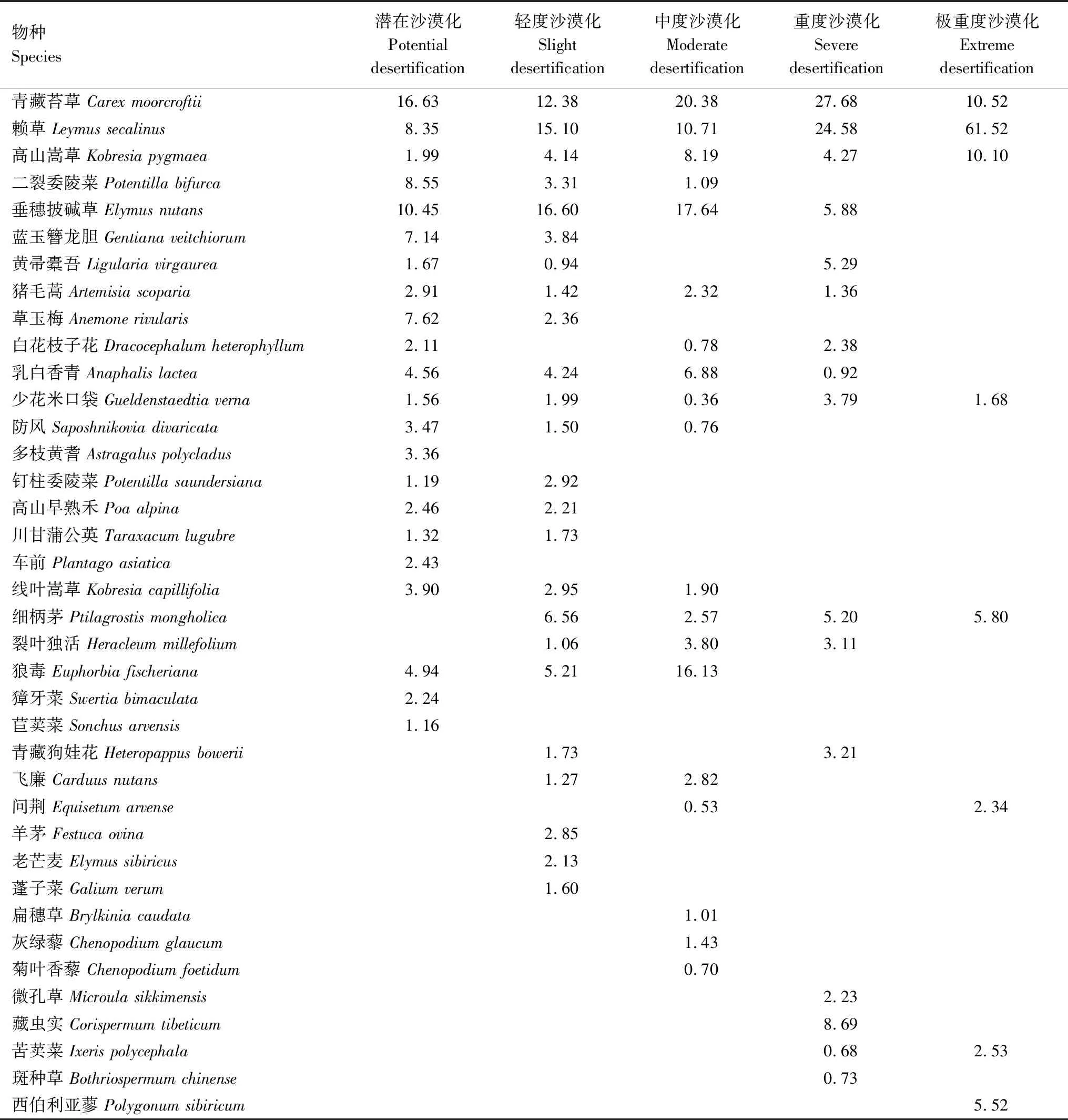
表2 不同沙化阶段高寒草甸群落的物种组成与重要值Table 2 The species composition and important value of the communities of alpine meadow in different desertification stages
3.2 不同沙化阶段高寒草甸表土环境因子变化特征
不同沙化阶段高寒草甸表土环境因子的动态变化如表3所示,表土水分在潜在沙化阶段达到峰值,为19.72%,显著高于其他沙化阶段(P<0.05),伴随沙化加剧,轻度、中度、重度和极重度阶段的表土水分依次为12.94%、11.62%、9.26%和10.51%,相互间差异不显著(P>0.05)。表土温度的变化趋势与水分不同,随沙化加剧而波动上升,其中轻度阶段最低,为24.32℃,极重度阶段最高,达30.79℃,除潜在、轻度和中度阶段,以及潜在、中度和重度阶段外,其余各阶段的差异达到显著水平(P<0.05)。表土电导率的变化倾向与水分相似,呈随沙化发展而逐步下降的动态规律,其在潜在阶段的数值最高,达0.1051 ms/cm, 且与其余各阶段间差异显著(P<0.05)。分析可见,伴随沙化的加剧,表土水分和电导率逐步降低,表土温度趋于增加,高寒草甸呈现出暖干化趋势。

表3 不同沙化阶段高寒草甸表土环境因子变化特征Table 3 Change characteristics of soil surface environmental factors of alpine meadow in different desertification stages
3.3 不同沙化阶段高寒草甸植物群落特征与物种多样性的变化
植被是沙漠化发展的指示器,伴随高寒草甸沙化程度的加剧,植物盖度、多度和物种丰富度呈不同程度的下降趋势(表4)。植物盖度在潜在阶段高达94.75%,而极重度阶段仅有4.00%,除重度和极重度之间,其余沙化阶段间差异显著(P<0.05)。物种丰富度在轻度阶段达到峰值,为9.00,极重度阶段最低,仅有2.25,处于中间值的由高至低依次是潜在、中度与重度阶段。沙漠化正向演替过程中,植物多度下降幅度剧烈,轻度、中度、重度和极重度阶段较潜在阶段分别下降了60.66%、65.08%、87.21%和95.60%,下降程度达到显著水平(P<0.05)。沙化的发展并未对植物株高产生规律性的影响,除轻度与极重度阶段外,其余各阶段间植物株高差异不显著(P<0.05)。
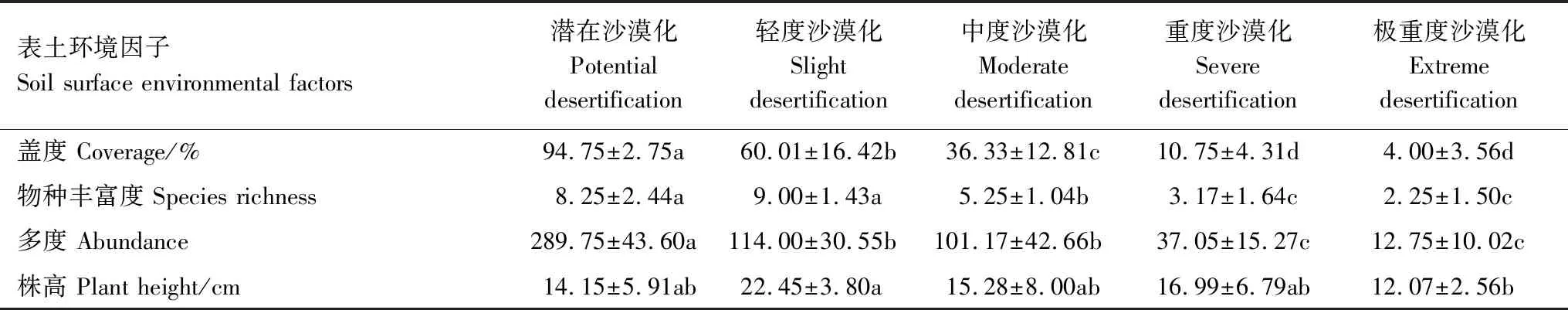
表4 不同沙化阶段高寒草甸植物群落特征Table 4 Characteristics of plant communities of alpine meadow in different desertification stages
沙化过程对高寒草甸植物群落的物种多样性、优势度及均匀度影响显著(P<0.05),且3者间的变化规律一致,均呈现出潜在至轻度阶段升高,之后伴随沙化加剧逐步下降的动态特征(图1)。各沙化生境中,潜在、轻度、中度、重度和极重度阶段Shannon-Wiener指数分别是1.25、1.54、1.12、0.68、0.43;Simpson指数依次为2.56、3.52、2.65、1.74和1.49;Pielou指数分别为0.58、0.75、0.71、0.50和0.36。综上可见,在沙化演替初期,群落中物种多样性水平有所增加,但随着生态退化的持续,多样性水平迅速下降,群落的空间异质性逐步增强。
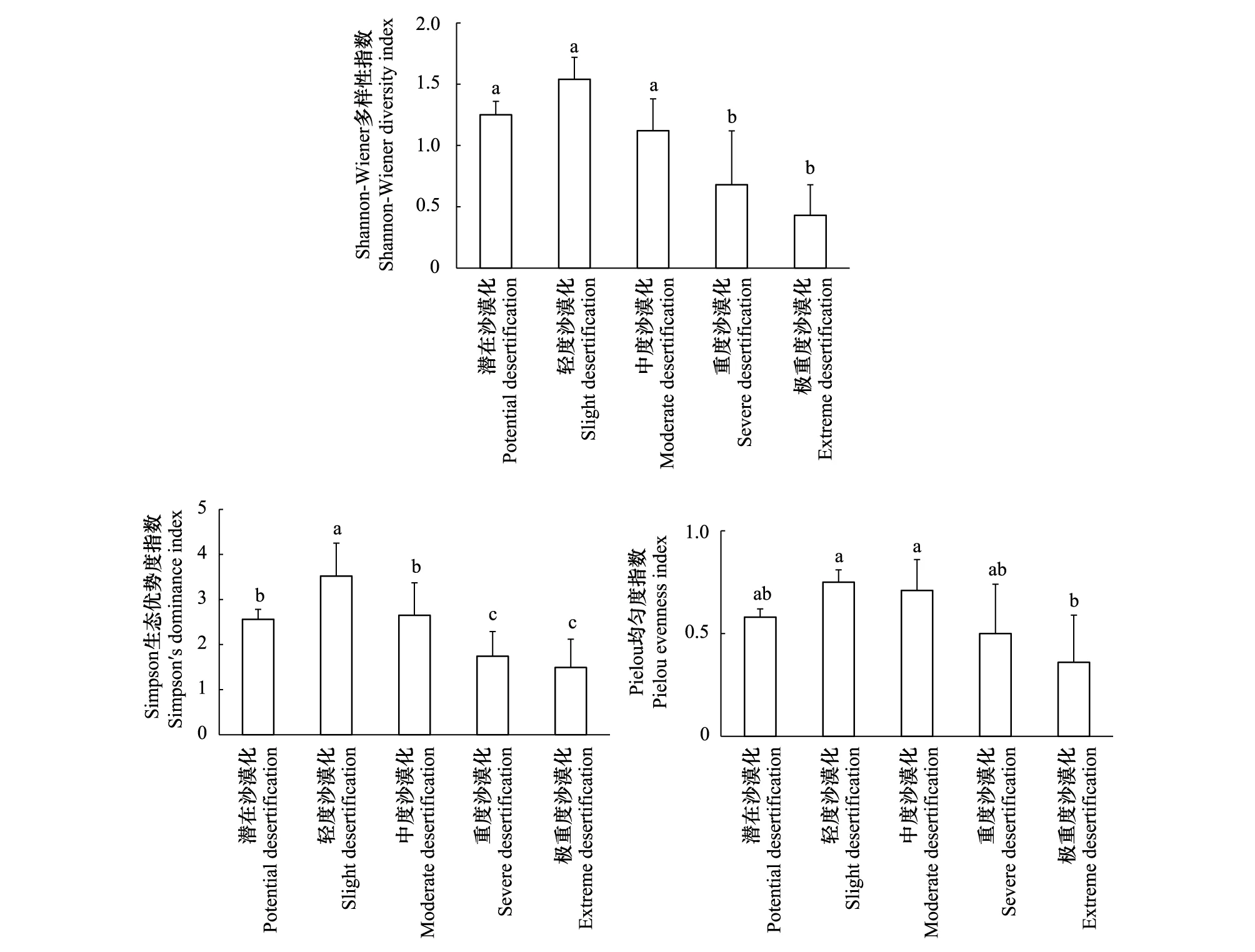
图1 不同沙化阶段高寒草甸植物多样性指数Fig.1 Shannon-Wiener index, Simpson index and Pielou indexes of alpine meadow in different desertification stages不同字母表示显著水平达到P<0.05
3.4 不同沙化阶段高寒草甸表土环境因子对群落组成的影响
排序图能够直观的描述物种组成在环境中的梯度变化。应用典范对应分析(CCA),可获得表达植物群落组成与表土环境因子相关性的二维排序图(图2)。结果表明,CCA第1轴的贡献率是44.93%,第2轴的贡献率是31.47%,累积方差贡献率达76.40%,表明排序效果较佳[27]。图中表土环境因子由带箭头的实线表示,沙漠化阶段应用实心方形表示,某一物种出现概率最高区域用对应的空心三角标注。箭头连线愈长,代表环境因子与物种分布格局的相关性越强。箭头连线交汇原点为各环境因子的均值点,箭头所指的方向表示各环境因子较平均值逐渐增加,相反方向代表较平均值不断下降。另外,将某一物种质点到环境因子的连线做垂直投影,投影点的位置可代表这些物种在该环境因子的最适值,投影点越接近箭头,最适值越高;越远离箭头,最适值愈低。由图2可知,表土水分与电导率对植物群落组成有较强的解释能力,且相互间呈正相关,而表土温度与前两者的变化趋势相反,表现出负相关性。植被方面,二裂委陵菜、 蓝玉簪龙胆(Gentianaveitchiorum)和草玉梅(Anemonerivularis)等物种定植于潜在沙化草甸的概率最高,且适生环境呈现出表土水分与电导率较高、表土温度偏低的特征。赖草、青藏苔草、垂穗披碱草、高山嵩草和裂叶独活(Heracleummillefolium)等物种出现在轻度与中度沙化草甸的频次最高,其中青藏苔草与垂穗披碱草喜生于表土水分、温度和电导率适中的环境,赖草、裂叶独活与高山嵩草更适宜表土温度较高,水分与电导率相对较低的区域。问荆(Equisetumarvense)、藏虫实(Corispermumtibeticum)、西伯利亚蓼(Polygonumsibiricum)等物种则常生长在重度和极重度沙化草甸中,生境条件呈现出表土水分与电导率较低,表土温度偏高的特点。
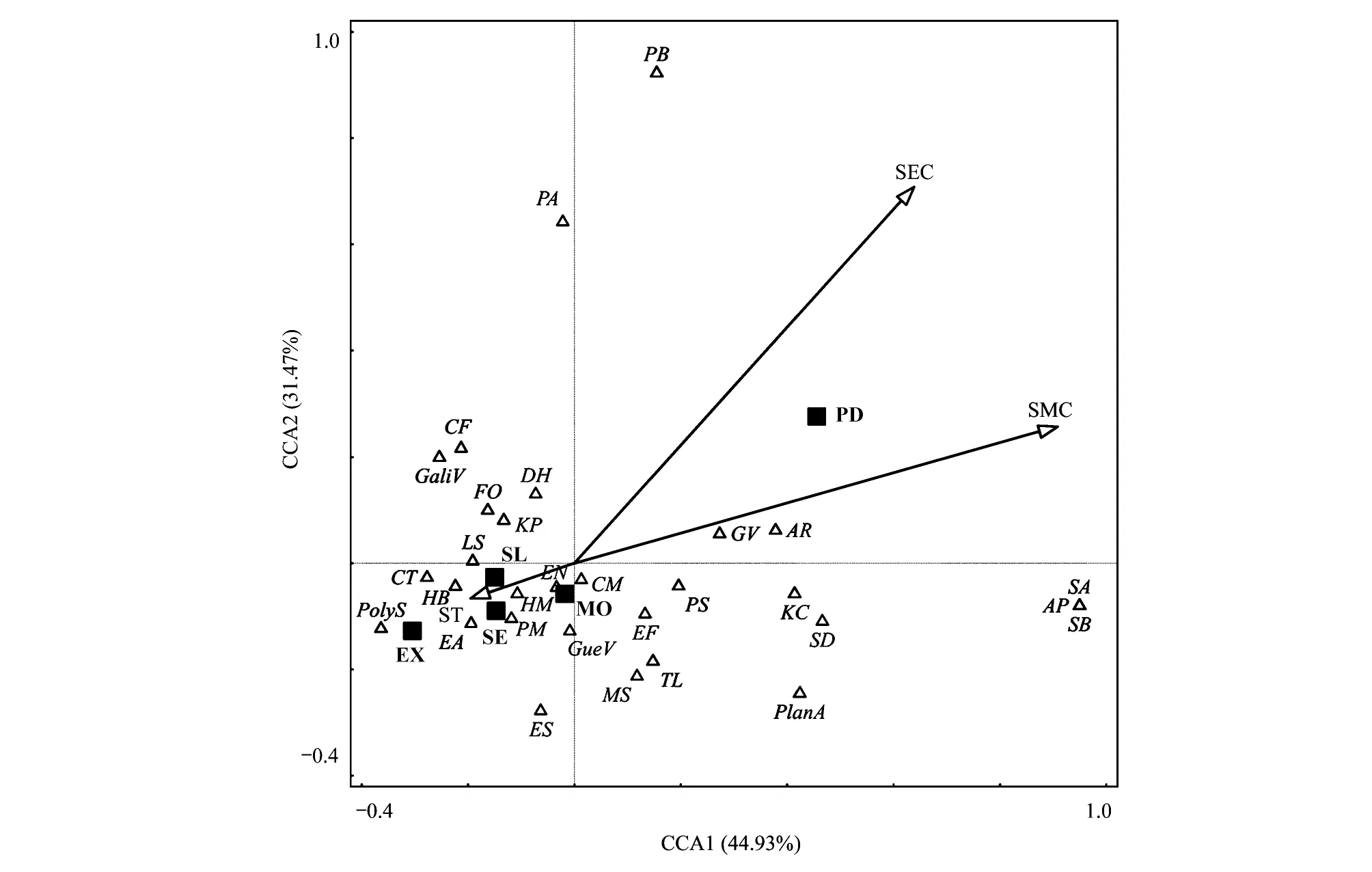
图2 不同沙化阶段高寒草甸表土环境因子与群落组成的CCA(典范对应分析)排序图Fig.2 CCA (Canonical correspondence analysis) ordination diagram of community composition and soil surface environmental factors of alpine meadow in different desertification stagesEX:极重度沙漠化, Extreme desertification; SE: 重度沙漠化, Severe desertification; MO: 中度沙漠化, Moderate desertification; SL: 轻度沙漠化, Slight desertification; PD: 潜在沙漠化, Potential desertification; SMC:表土水分, Soil moisture content; ST: 表土温度, Soil temperature; SEC: 表土电导率, Soil electric conductivity; PB: 二裂委陵菜, Potentilla bifurca; PA: 高山早熟禾, Poa alpine; CF: 菊叶香藜, Chenopodium foetidum; GaliV: 蓬子菜, Galium verum; FO: 羊茅, Festuca ovina; DH: 白花枝子花, Dracocephalum heterophyllum; KP: 高山嵩草, Kobresia pygmaea; LS: 赖草, Leymus secalinus; CT: 藏虫实, Corispermum tibeticum; HB: 青藏狗娃花, Heteropappus bowerii; HM: 裂叶独活, Heracleum millefolium; EN: 垂穗披碱草, Elymus nutans; PolyS: 西伯利亚蓼, Polygonum sibiricum; EA: 问荆, Equisetum arvense; PM: 细柄茅, Ptilagrostis mongholica; GueV: 少花米口袋, Gueldenstaedtia verna; ES: 老芒麦, Elymus sibiricus; AR: 草玉梅, Anemone rivularis; GV: 蓝玉簪龙胆, Gentiana veitchiorum; CM: 青藏苔草, Carex moorcroftii; PS: 钉柱委陵菜, Potentilla saundersiana; KC: 线叶嵩草, Kobresia capillifolia; SA: 苣荬菜, Sonchus arvensis; SB: 獐牙菜, Swertia bimaculate; AP: 多枝黄耆, Astragalus polycladus; EF: 狼毒, Euphorbia fischeriana; SD: 防风, Saposhnikovia divaricate; MS: 微孔草, Microula sikkimensis; TL: 川甘蒲公英, Taraxacum lugubre; PlanA: 车前, Plantago asiatica
应用向前筛选法(Forward selection)和蒙特卡洛置换检验(Monte Carlo permutation test)对表土环境因子进行量化分析,可获得表土环境因子对植物群落组成的解释强度(表 5)。研究表明,3个表土环境因子对群落组成的总方差解释率为31.10%,解释强度由大到小依次为表土水分、电导率和温度,其中表土水分和电导率对群落组成的影响达到极显著水平(P<0.01)。综上可以看出,表土水分和电导率对物种分布与迁移的影响较表土温度更为深远。

表5 表土环境因子对群落组成的方差解释率与贡献率Table 5 Contribution and explained variation of soil surface environmental factors to community composition
3.5 不同沙化阶段高寒草甸表土环境因子与植物群落特征及多样性水平的关系
伴随沙漠化加剧,高寒草甸表土环境因子对植物群落特征的表征能力和相关关系均呈不同程度的变化(图 3)。潜在沙化阶段,CCA前两轴的方差解释率为71.91%,排序效果良好[27],表土环境因子对植物群落特征及多样性水平的解释能力较强。环境因子中,表土水分与电导率间呈正相关,与表土温度呈负相关。Shannon-Wiener指数、Simpson指数与物种丰富度等指标有随表土水分与电导率增加而增加的趋势。植物盖度与多度同表土电导率间呈正相关,而与表土水分和温度无明显相关性。株高则与表土温度间呈轻度的正相关。轻度阶段,CCA第1、第2轴总方差解释率是55.25%,排序效果可靠,说明表土环境因子对植物群落特征及多样性指数的表征能力较高。表土水分、温度、电导率3者间的相关性与潜在沙漠化阶段相似。植物盖度、多度、物种丰富度与表土温度呈正相关,Shannon-Wiener指数、Simpson指数与表土电导率及温度间存在正向弱相关。中度、重度和极重度沙化阶段CCA前两轴的方差解释率分别为27.05%、32.04%和25.68%,整体解释率低于40%,排序效果欠佳[27]。中度阶段,Pielou指数、多度同表土环境因子间存在正相关,其余植物群落指标与表土环境因子的相关性较弱或呈负相关。重度阶段,物种丰富度、多样性指数同表土水分、电导率间存在明显正相关,而与植物生产力水平密切相关的多度、盖度则同表土温度间呈负相关。极重度阶段与重度阶段存在一定相似性,Shannon-Wiener指数、Pielou指数同表土水分、电导率间存在较好的正相关,不同之处在于多度、盖度与表土电导率的相关强度提升明显。综上,伴随沙化加剧,表土环境因子与植物群落指标、物种多样性间的相关性存在差异、其对植物群落特征和多样性水平动态变化的解释力度逐渐下降。
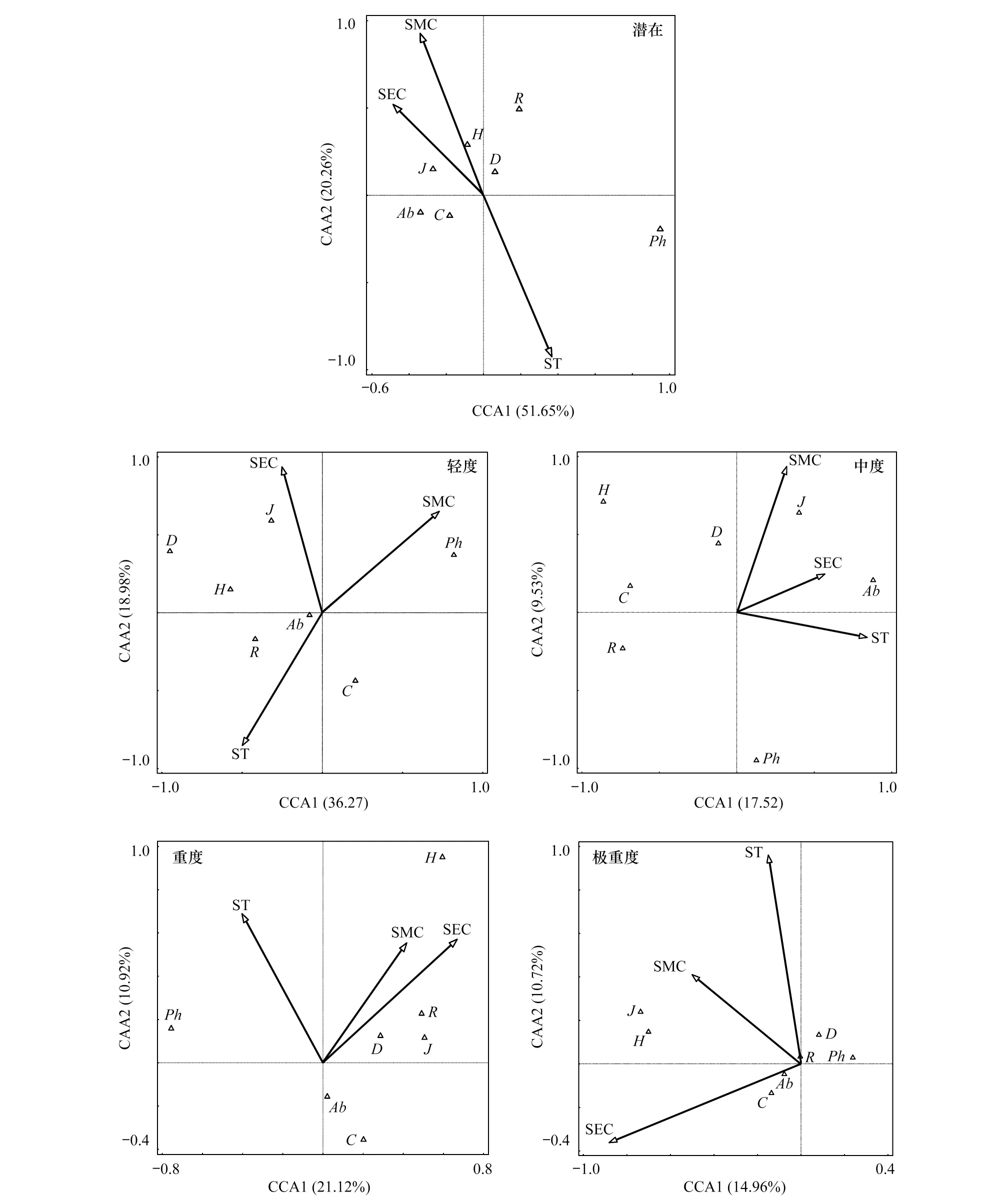
图3 不同沙化阶段高寒草甸表土环境因子与植物群落特征的CCA(典范对应分析)排序图Fig.3 CCA (Canonical correspondence analysis) ordination diagram of plant community characteristics and soil surface environmental factors of alpine meadow in different desertification stagesSMC:表土水分, Soil moisture content; ST: 表土温度, Soil temperature; SEC: 表土电导率, Soil electric conductivity; C: 盖度, Coverage; R: 物种丰富度, Species richness; Ab: 多度, Abundance; Ph: 株高, Plant height; H: Shannon-Wiener多样性指数, Shannon-Wiener index; D: Simpson 优势度指数, Simpson index; J: Pielou均匀度指数, Pielou index
4 讨论
4.1 高寒草甸沙化过程中表土环境因子变化的原因
玛曲高寒草甸生态系统中,植被类型基本为草本,根系密集而短小,决定了植被与土壤因子作用最紧密、影响最显著的区域分布在土壤表层,因此,表土环境因子对于高寒草甸植被的变化具有较强的解释能力。表土水分、温度是制约植物定植、生长、迁移与演替的关键因子[28-29],而表土电导率是土壤中可溶性离子和盐分含量的直观表征,在非盐渍化地区,一定程度上能够反映表层土壤养分的含量[30]。本研究中,表土水分和电导率随沙化的发展逐渐降低,这一发现与多个沙化生态系统的研究结果一致[31-33]。主要原因是草甸沙化过程中,风对地表土层的分选作用加强,土壤中养分含量丰富的黏粉粒遭到吹蚀与破坏,不易迁移的砂质颗粒比例上升,土质趋于粗化与贫瘠化,最终导致容重增加,渗透阻力加强,水分的存储与传输受到限制[4, 34]。表土温度的变化趋势与水分不一致,随沙化加剧而逐步增加,可能原因是沙漠化引起的植被衰退造成地表裸露,缺乏遮阴保护的土壤直接作用在太阳辐射下,导致土表热量聚集,温度上升明显。
4.2 高寒草甸沙化过程中植物群落组成与物种多样性的变化规律
高寒草甸沙漠化改变了土壤表层的物理环境与生态格局,主要表现为水分与养分的流失、土质粗化与风蚀加强[4, 29, 35],伴随这一过程,玛曲高寒草甸的群落组成、特征与物种多样性均产生了明显变化。随着沙化的正向演替,研究区植被发生退化,青藏苔草和垂穗披碱草等优势种重要值逐渐下降,二裂委陵菜、高山早熟禾、蓝玉簪龙胆和草玉梅等次优势种与伴生种逐步退出群落,这也为部分典型退化种的定植与繁衍提供了便利,如狼毒、微孔草(Microulasikkimensis)、藏虫实(Corispermumtibeticum)等。伴随着生境逐步恶化,养分匮乏、高温干旱的土壤表层已难以承载复杂、稳定的植物群落,群落结构趋于简单,优势种重要值占比增加。
沙化阶段不同,高寒草甸的植物群落特征与多样性水平亦不同。植物生产力是表征植物生态功能稳定性和区域生态承载力的关键因子,可由植物盖度、多度等群落指标直观反映[4,36-37]。本研究中,伴随玛曲高寒草甸沙化的加剧,植物盖度与多度下降明显,相较未发生实质退化的潜在沙化阶段,极重度阶段植物的盖度与多度分别下降了95.78%和95.60%,盖度与多度的大幅降低表明沙化对植物生产力和土壤承载力具有巨大的负面效应,这种消极性的破坏作用伴随了整个沙化的演替过程,使群落结构趋于分解与失衡。沙漠化演替过程中,株高的变化呈现一定的无序性,由高到低依次为轻度、重度、中度、潜在和极重度,主要原因是群落株高很大程度上取决于优势种与主要伴生种的类型,而与沙化进程关联不大,这也说明了株高对沙化阶段的指征作用不明显。不同于植物盖度与多度,沙化各阶段中,物种多样性(Shannon-Wiener指数和Simpson指数)与均匀度水平(Pielou指数)在轻度阶段最高,潜在和中度阶段次之,之后伴随沙化加剧迅速降低,主要原因是沙化轻度阶段,环境对高寒草甸原生植物的干扰是可控的,生境中的能量与资源分布在干扰下产生再分配,这种分配效应一定程度上加剧了种间竞争,为其他物种的入侵与定植提供了可能[4,38-39],在资源仍相对充足的条件下,物种的多样性和均匀度趋于增加,而当干扰持续加强,沙漠化进入重度和极重度阶段,表现为生境中的资源大量流失,多数物种退出群落,群落内多样性水平降低,优势度增加[24]。整体来看,沙化演替过程中植物群落多样性的动态变化符合“中度干扰假说”。
4.3 决定高寒草甸沙化过程中植物群落与物种多样性变化的主导因素
表土水分、温度和电导率在一定程度上解释了(31.10%)玛曲高寒草甸沙化过程中植物群落的组成结构与演替规律。伴随沙化发展,生境呈现出土表水分与养分逐渐降低、温度曲折上升的变化规律,群落出现湿生—中生—旱生的演替格局[40],喜冷湿环境的草玉梅、蓝玉簪龙胆等物种逐步退出,取而代之的是较为耐旱的微孔草、藏虫实等退化种。同时,赖草作为优势种伴随了整个退化过程,是沙化生境中维持群落的关键种。
温度和水分是高寒草甸区影响植物生长的关键环境要素[10]。本研究中(图 3),潜在沙化阶段植物的多度、盖度同表土温度和水分的相关性较差,可能源于该阶段生态系统未发生退化,生境承载力较高且基本饱和,植物生产力处于稳定水平,对外界环境的适度波动不敏感。Shannon-Wiener指数、Simpson指数和Pielou指数与表土水分、电导率呈正相关,而与温度间呈负相关,可能原因与适生于潜在沙化区域的群落类型有关,冷湿环境为蓝玉簪龙胆、草玉梅、黄帚橐吾(Ligulariavirgaurea)和苣荬菜(Sonchusarvensis)等物种的定植与繁衍提供了便利。轻度沙化阶段,植物盖度、多度、物种丰富度以及多样性指数同表土温度呈正相关,而与表土水分呈一定的负相关性。主要原因是轻度阶段表土温度为24.32 ℃,属各沙化阶段最低,温度本底值较低时,热量增加能够有效提升土壤微生物活性与植物机体代谢能力,解除了低温对植物的限制,有助于种群的繁衍与扩张[10,41-42]。同时,轻度阶段土壤水分充沛,足够植物生长所需[10],当水分持续增加时,部分中生与旱生物种的入侵与繁育将受到限制。沙化发展至中度、重度乃至极重度阶段,表土水分与电导率对植物生产力与多样性水平的正面效应愈加突出,表土温度的影响由积极转向负面或不相关。主要原因一方面是伴随沙化加剧,土壤团粒结构破坏严重,富含养分的黏粉粒大量散失,土壤质地呈明显的粗粒化与贫瘠化特征,与此同时,土壤机械阻力增加,孔隙度减少,表层水分难以留存[34],这一过程中,玛曲高寒草甸土壤的水养条件由富足转向匮乏,植物生长发育对其的需求趋于紧迫。另一方面,表土温度持续上升加剧了水分散失,且温度过高抑制了微生物活性,不利于枯落物分解与养分周转,植物生长受到限制[43]。值得注意的是,中度、重度和极重度阶段,表土环境因子对排序图的方差解释率不足40%,表明排序图虽然在一定程度上能够反映这些沙化阶段表土温度、水分、电导率与植物群落特征及多样性指数的关系,但这种关系表达并不全面,预示着其他环境要素对植物群落特征与多样性水平的影响更为深刻,依据前人研究,结合样地中频繁出现的牲畜粪便和踩踏痕迹,可判断这些影响主要源于人类的放牧活动[10, 44-46]。鉴于此,今后高寒草甸生态系统的沙漠化监测中,特别是沙化发展到中度乃至更严重阶段时,放牧活动的测度与管理应是关注的重点。
5 结论
研究明晰了玛曲高寒草甸沙漠化各阶段表土环境因子与植物群落的变化规律及两者间的相关性。
1)伴随沙化加剧,表土水分和电导率逐渐降低,表现为潜在阶段达到峰值,极重度阶段降至谷底;温度则呈波动上升趋势,其中轻度阶段最低,极重度阶段最高。
2)伴随沙化加剧,植物的盖度、多度依次降低,且下降幅度达到显著水平(P<0.05)。物种丰富度、多样性指数与均匀度指数的变化趋势基本一致,均表现为潜在至轻度阶段升高,之后不断下降的动态规律。
3)典范对应分析(CCA)表明,表土水分、温度及电导率能够解释植被分布格局的31.10%,其中表土水分和电导率对植物群落组成的影响程度达到极显著水平(P<0.01)。潜在和轻度阶段,CCA排序效果较佳,说明在沙化初期,表土水分、温度和电导率能够较好的表征植物群落特征和物种多样性的变化规律,是植物群落发育与构建的关键因子。中度、重度和极重度阶段,CCA排序效果较差,主要原因是放牧活动的干扰,这也说明沙化加剧至中度阶段后,放牧等人类活动取代表土环境因子,成为影响高寒草甸植被生长与沙漠化演替的主导因素。
由于本研究依托于全国性的荒漠化和沙化监测项目,该课题属于多尺度、大范围的科技普查项目,在充分考虑科学性、可靠度的同时需兼顾成本、适用范围等限制因素,因此在指标选取中存在一定的局限性,如应用准确度较差的电导率替代土壤有机质来表征土壤养分。鉴于此,对于某些生境条件特殊的敏感生态区,如玛曲高寒草甸沙化区,应将土壤有机质、放牧强度等关键环境因子纳入监测指标,以对高寒草甸区域沙化过程中植物群落的组成与演替提供更加精准的环境解释。
猜你喜欢
成都信息工程大学学报(2022年4期)2022-11-18
资源导刊(2022年7期)2022-11-10
青海草业(2022年2期)2022-07-23
海外文摘·文学版(2021年3期)2021-05-19
水土保持研究(2020年6期)2020-10-21
考试与评价·高二版(2020年3期)2020-09-10
考试与评价·高二版(2020年3期)2020-09-10
活力(2019年21期)2019-04-01
绿色中国(2017年7期)2017-08-02
绿色中国(2016年1期)2016-06-05
