
New chroniosuchian materials from Xinjiang, China
2020-11-11 12:59:42LIUJun
古脊椎动物学报(中英文) 2020年4期
LIU Jun
(1 Key Laboratory of Vertebrate Evolution and Human Origins of Chinese Academy of Sciences, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences Beijing 100044)
(2 CAS Center for Excellence in Life and Paleoenvironment Beijing 100044 liujun@ivpp.ac.cn)
(3 College of Earth and Planetary Sciences, University of Chinese Academy of Sciences Beijing 100049)
Abstract Chroniosuchians have the earliest representatives in Gansu, China, but the Chinese records were scarcer compared to Russia, especially the Triassic one. In Xinjiang, there was only one specimen reported from the upper part of the Guodikeng Formation. Here three new chroniosuchian specimens are reported from three new stratigraphic horizons: the Quanzijie Formation (middle Permian), the base of the Guodikeng Formation (upper Permian), and the Jiucaiyuan Formation (Lower Triassic). The osteoderm from the Jiucaiyuan Formation represents the first definite Triassic chroniosuchian from China. The new findings increase the chroniosuchian diversity and their time range in China. The bystrowianid chroniosuchian specimens from the Guodikeng and Jiucaiyuan formations demonstrated that this group survived in the end-Permian mass extinction in Xinjiang, China.
Key words Xinjiang, China; Permian, Triassic; Quanzijie Formation, Guodikeng Formation,Jiucaiyuan Formation; Bystrowianidae, chroniosuchians
1 Introduction
The chroniosuchians were an enigmatic clade of non-amniotic tetrapods with uncertain phylogenetic position (Clack and Klembara, 2009; Schoch et al., 2010; Buchwitz et al., 2012;Witzmann and Schoch, 2018; Marjanović and Laurin, 2019). This group can be divided into two families, Bystrowianidae and Chroniosuchidae (Ivakhnenko and Tverdokhlebova, 1980;Novikov et al., 2000; Witzmann et al., 2019). The earliest representatives of both families are known from the middle Permian Dashankou fauna, Gansu, China (Li and Cheng, 1999; Jiang et al., 2017). The latest representative of Chroniosuchidae is known only from a single Triassic taxon,Madygenerpeton pustulatus, from the Ladinian or Carnian of Kyrgyzstan (Schoch et al.,2010); while those of Bystrowianidae are known from Middle Triassic of Russia and Germany(Novikov and Shishkin, 2000; Witzmann et al., 2008; Witzmann and Schoch, 2018).
This group was first reported from the late Permian Jiyuan fauna in China, i.e.,Bystrowiana sinica(Young, 1979); later, two more species were named and referred to Bystrowianidae for this fauna (Jiyuanitectum flatumandDromotectum largum) (Liu et al., 2014). Some postcranial bones were identified as bystrowianid from the Permo-Triassic Guodikeng Formation of Jimusar,Xinjiang, China (Liu and Abdala, 2017), but no definite Triassic chroniosuchian is known from China up to now. Our recent fieldwork showed that chroniosuchians were a diverse and persistent group in Xinjiang: they survived from the late Permian to at least the Early Triassic here. In this paper we reported three new chroniosuchian specimens from Xinjiang (Fig. 1), including the first definite Triassic one from the Jiucaiyuan Formation.

Fig. 1 Permian to Triassic stratigraphic sequence within Turpan-Hami Basin, Xinjiang, showing the horizons of studying specimens
AbbreviationsIVPP, Institute of Vertebrate Paleontology and Paleoanthropology,Chinese Academy of Sciences; SMNS,Staatliches Museum für Naturkunde Stuttgart,Germany.
2 Systematic paleontology
Chroniosuchia Tatarinov, 1972 Chroniosuchia indet.
MaterialIVPP V 26539, a left femur (Fig. 2).
Localityand horizonDalongkou section, Santai, Jimusar, Xinjiang, China; upper part of Quanzijie Formation (middle Permian).
DescriptionThe femur is slightly dorsoventrally compressed, and is not so curved as the femur of IVPP V 23295 (Liu and Abdala, 2017). It is slender and waisted as other chroniosuchians (Clack and Klembara, 2009; Liu and Abdala, 2017). The anterior margin is more concave than the posterior margin. The length of the bone is 40 mm, and the width is 13,15 mm respectively for the proximal and distal ends. Both expanded ends are incompletely ossified. The unfinished proximal articular surface is convex in outline dorsally and slightly concave ventrally when viewed mesially (Fig. 2D). It extends anteroventrally, forms a continuous surface with the unfinished internal trochanter. The proximal articular surface decreases in dorsoventral height backwards and forms a pointed angle posteriorly.
The dorsal surface of the femur bears striations on the proximal side (Fig. 2A). The dorsal surface of the distal end of the femur is divided by a relatively broad intercondylar groove (Fig. 2E). The anterior condyle extends slightly wider than the posterior one. Due to the incomplete ossification, the anterior condyle projects distally similar to the posterior condyle.The anterodorsal surface of the anterior condyle, near the middle, bears distinct longitudinal ridges for ligaments.
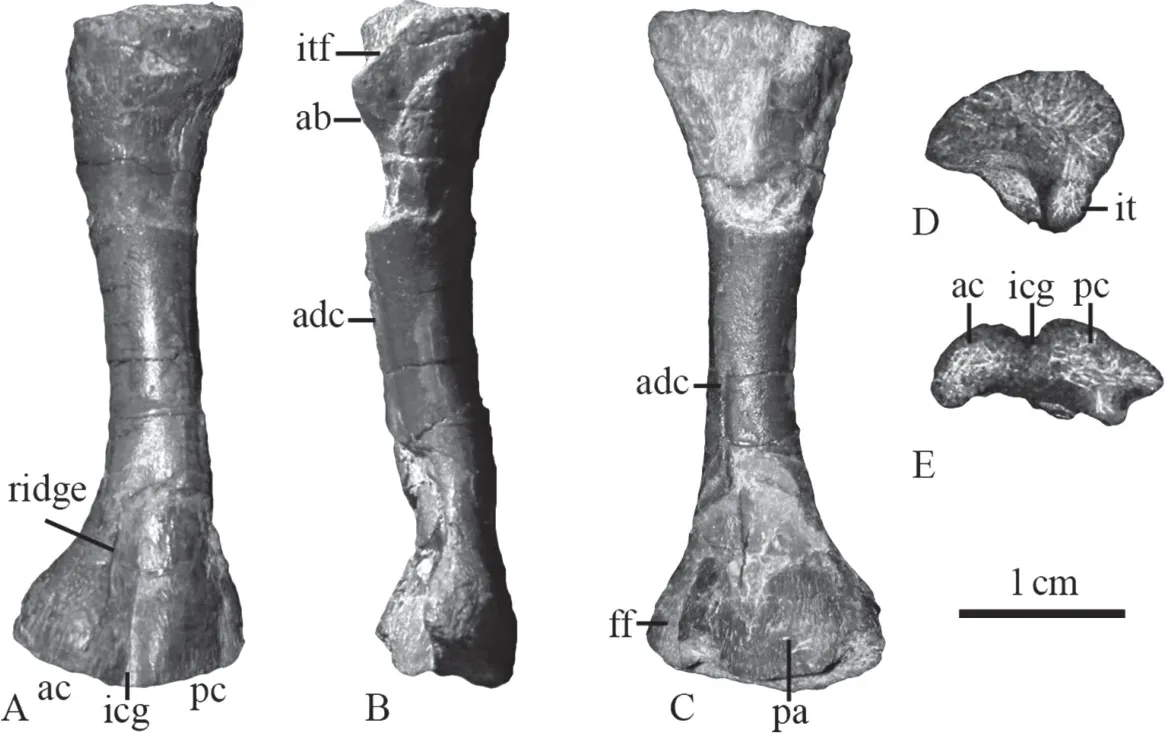
Fig. 2 Chroniosuchian left femur (IVPP V 26539) from the Quanzijie Formation in Jimusar, Xinjiang in extensor (A), anterior (B), flexor (C), proximal (D), and distal (E) views
The adductor blade is low, directed posteroventrally, forming the anterior margin of the large intertrochanteric fossa (Fig. 2B). The anterior surface of the adductor blade is rugose.Distal to the adductor blade, a low but distinct adductor crest is slightly curved, runs distally near the posterior margin of the shaft on the ventral surface, and ends by a crack (Fig. 2C).Distal to the crest, there is a low ridge to a point close to the triangular popliteal area. However,it is likely separated from the adductor crest. Posterior to it, another crest runs on the posterior side of the shaft ending on the distal posterior corner. Two crests frame the small fibula fossa on the posteroventral corner of the distal end.
DiscussionThe shape of this femur is primitive among tetrapods. However, it is slender,similar to that of temnospondyls likeTrimerorhachis(Pawley, 2007) or chroniosuchians (Clack and Klembara, 2009; Liu and Abdala, 2017; Witzmann and Schoch, 2018), rather than robust as in many other basal tetrapod groups (Witzmann and Schoch, 2018; Bazzana et al., 2020). As in IVPP V 23295 andBystrowiella schumanni, the adductor crest follows a diagonal course,ending distally close to the posterior margin of the shaft (Liu and Abdala, 2017; Witzmann and Schoch, 2018). In most Permian tetrapods, adductor crest generally runs towards, not posterior to the popliteal area (Fox and Bowman, 1966; Klembara and Bartík, 2000; Pawley, 2007). This could be a diagnostic character of chroniosuchians.
IVPP V 26539 also shares the following features with other chroniosuchian femurs (V 23295 and SMNS 96948): the adductor blade is directed posteroventrally, and the posterior end of the proximal articular surface is pointed. So this specimen can be referred to Chroniosuchia.
This specimen has unfinished ends, as in the much longer femur of SMNS 96948, may indicated juvenile state of both specimens. Meanwhile, a medium sized femur of V 23295 is well-ossified, indicated that species has a smaller adult size.
Chroniosuchia Tatarinov, 1972 Bystrowianidae Vjuschkov, 1957 Bystrowianidae indet.
MaterialIVPP V 26540, four vertebrae, one rib, and several scutes (Fig. 3).
Locality and horizonTaoshuyuan (Taoxigou) section, Turpan, Xinjiang; base of the Guodikeng Formation (upper Permian).
DescriptionThere are four articulated and one isolated vertebrae, two of them are nearly complete (Fig. 3). They are identified as the vertebrae around sacral region. The vertebrae have a basic shape of Bystrowianidae (Liu et al., 2014; Liu and Abdala, 2017;Witzmann and Schoch, 2018).
IntercentraOnly two articulated intercentra are exposed (Fig. 3A, B). The anterior one shows smooth periosteal bone on ventral and lateral surfaces. Its lateral surface bears part of the parapophyses for articulation with the capitulum near the posterior margin, and ventral surface has no haemal arch. In lateral view, its length relative to the pleurocentrum is similar to IVPP V 23295, much narrower than inChroniosaurus dongusensis(Clack and Klembara,2009).
PleurocentraThe lengths of the pleurocentra are approximately 10 mm, while the heights are about 9 mm (Fig. 3A, B). The pleurocentra are massive with a round cross-section,and they are not perforated by the notochord (Fig. 3C, D). The ventral surface is relatively flat (slightly convex on the anterior one, slightly concave on the posterior one) with faint ventral ridges on three articulated larger pleurocentra. Two longitudinally aligned, low ridges are clear on the narrow ventral face of the smaller pleurocentrum of the isolated vertebra.This feature was proposed as a bystrowianid characteristic (Witzmann and Schoch, 2018).The neural arches are fused to the pleurocentra, but their suture is clear; so this specimen looks like in a younger stage than V 23295, in which the suture is absent. This suture runs across the diapophysis in three articulated vertebrae, indicating the pleurocentrum participates in formation of the diapophysis. On two posterior pleurocentra, the anterior half of the ventrolateral surface participates in formation of the parapophysis, which is separated from the diapophysis by a narrow groove. So they are identified as the sacral and the first caudal vertebrae, as inKotlassia(Bystrow, 1944) andProterogyrinus(Holmes, 1984).
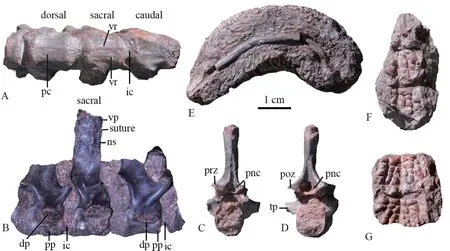
Fig. 3 Bystrowianid specimen (IVPP V 26540) from the Guodikeng Formation in Turpan, Xinjiang
Neural archesThe prezygapophyses are oriented dorsomedially and slightly anteriorly,and the postzygapophyses are oriented ventrolaterally and posteriorly (Fig. 3B-D). The paired openings of the paraneural canals are visible between and ventral to the zygapophyses on anterior and posterior surface in the better prepared isolated vertebra as in other bystrowianids(Tatarinov, 1972; Novikov and Shishkin, 2000; Witzmann et al., 2008; Liu et al., 2014; Liu and Abdala, 2017; Witzmann and Schoch, 2018). The openings are large as inAxitectum(Novikov and Shishkin, 2000),Bystrowiana(Liu et al., 2014) and V 23295 (Liu and Abdala,2017), much bigger than those ofSynesuchus(Novikov and Shishkin, 2000) andBystrowiella(Witzmann et al., 2008; Witzmann and Schoch, 2018).
The transverse processes are very short and massive, directed ventrolaterally. The facets of the diapophyses are strongly enlarged in the sacral and first caudal. In the isolated vertebra,the transverse processes are completely formed by the neural arch. It is probably derived from the anterior tail region.
The neural spines are only nearly complete in the sacral vertebra and one isolated caudal vertebra. The neural spines are anteroposteriorly slightly shorter than the pleurocentrum. In the sacral, its anterior and posterior margins are nearly parallel. Its height is less than twice the height of the pleurocentrum. Its dorsal tip is connected with the osteoderm by interdigitating sutures. In the isolated caudal, the neural spine widens dorsally near the tip then narrows. Its dorsal tip does not carry an osteoderm.
RibA curved incomplete rib measures more than 5 cm in original length (Fig. 3E). The proximal side is not preserved.
OsteodermsThere are two pieces of impression of the dermal surface of the osteoderms,one with three osteoderms, and the other with two osteoderms (Fig. 3F, G). Neither of the osteoderms is complete, so it is unsure on the width of the osteoderms. The distinct dermal sculpturing is composed of ridges and depressions. A mid-ridge is preserved. The pits have different sizes on two sides of the ridge, and they likely arrange in posterolaterally radiated line. No prominent parasagittal ridge is observed.
DiscussionThis specimen is diagnosed as a bystrowianid chroniosuchian for the presence of the sculptured dorsal osteoderms which are sutured with the neural spine, paired deep paraneural canals on anterior and posterior surfaces of the neural arch. It cannot be further diagnosed for the poor preservation of the osteoderms.
Chroniosuchia Tatarinov, 1972 Bystrowianidae Vjuschkov, 1957 unnamed new taxon
MaterialIVPP V 26541, an incomplete dorsal osteoderm (Fig. 4).
Locality and horizon18J15 (N43°56′6.8″, E88°54′19.5″) (4 km east to the Dalongkou section), Santai, Jimusar, Xinjiang; Jiucaiyuan Formation (Lower Triassic).
DescriptionThe single osteoderm is slightly convex dorsally. Most of the margins are broken other than right anterolateral margin (Fig. 4). The complete width should be approximately 3 cm.

Fig. 4 Bystrowianid osteoderm (IVPP V 26541) from the Jiucaiyuan Formation in Jimusar, Xinjiangin dorsal (A), ventral (B), and posterior (C) views
The dermal sculpture is not symmetric, and it is dominated by different-sized polygonal pits formed mainly by longitudinally oriented and oblique ridges. Transversely elongated sculptural depressions are only present near the posterior area. The sculpture has no axial crest or any parasagittal ridge, as inSynesuchus muravjeviorDromotectum spinosum(Novikov and Shishkin, 2000). The right anterior wing is preserved its posterior portion, with a smooth dorsal facet for the facies alaris (ventral facets) of the preceding osteoderm. The right accessory process is also partially preserved. The maximum width of one anterior wing is estimated as 9 mm, slightly smaller than the width of accessory processes.
The incomplete median articular plate measures 10 mm in width. It lies posteriorly and slightly ventral to the sculptured dorsal plate. The median crista decreases in width backwards and is triangular in shape in dorsal view. The broad lateral cristae are longer than and located at the same level as the median crista. Most parts of these cristae are formed by tapered extensions of the posterior margin of the sculptured dorsal plate. As most bystrowianids,two deep longitudinal grooves (sulci articulares) lies between the median and lateral crests,for accommodation of the accessory processes and associated ligaments of the successive osteoderm (Novikov and Shishkin, 2000; Shishkin et al., 2014). The area marginalis (marginal zone) is incompletely preserved on the right side. It extends anteriorly ventral to the dorsal plate as inBystrowianaandJiyuanitectum(Novikov and Shishkin, 2000; Liu et al., 2014).
On the ventral surface, the shallow median depression, between the anterior wings back to the base of the ventral process, is preserved for most of the right side and nearly posterior half of the left side. The ridge extends from the right accessory process is distinct. The sulcus medius (median groove) should be narrow. A broad, low but distinct crista obliqua (oblique crest) extends along the ventral surface of the osteoderm posteromedially from the anterior wings toward the region of the ventral process, and continuous to the margin zone of the median articular plate. Lateral to the ventral margin, a shallow depression on the posterior part of the ventral surface is the ventral facet that overlapped the dorsal facet of the anterior wing of the succeeding osteoderm.
The base of the ventral process is located in the posterior half of the osteoderm, and its posterior margin extends posteriorly beyond the boundary of the sculptured surface of the osteoderm. The process is ovate in transverse section and lacks anterior and posterior extensions.
DiscussionIVPP V 26541 can be referred to Bystrowianidae based on the unpaired posterior articular plate and weakly expanded plate bearing broadly separated anterior facets.This specimen is coded for the osteoderm characters of Witzmann et al. (2019) (Appendix 1),and the coding of character 1 ofDromotectumis changed to 1&2 because the ratio is greater than 1.5 inD. abditum(Shishkin et al., 2014). A single most parsimonious tree (length=33)is recovered and nearly identical to figure 5 of Witzmann et al. (2019) other than IVPP V 26541 forming the sister-group ofDromotectum. However, no synapomorphy supports this clade. IVPP V 26541 shares one synapomorphy with the clade (Vyushkoviana+ (Dromotectum+Axitectum)): the absence of parallel to subparallel transversal bars present in the dorsal ornamentation (3). It also shares two of four synapomorphies with the clade (Dromotectum+Axitectum): axial process of posterior articular plate is positioned at the same level as the lateral processes (13) and posterior part of the base of the ventral process positioned posterior to the boundary of the ornamented region (21). However, it is different from them by the osteoderm plate with an angle between neural spine and osteoderm plate greater than 70°(19). IVPP V 26541 represents a new taxon closely related toDromotectum. However, it is not named here and should be named on more complete material. Its close relationship withDromotectumis not surprise, becauseDromotectumexisted in the upper Permian of Henan,China (Liu et al., 2014).
3 Discussion and conclusion
The new specimens described here increase the diversity of Permo-Triassic tetrapods.Previously, only one tetrapod species,Kunpania scopulusa, was reported from the top of the Quanzijie Formation (Sun, 1978). Also, only one chroniosuchian specimen was reported from the Guodikeng Formation (Liu and Abdala, 2017). Although chroniosuchian should have existed from middle Permian in Xinjiang based on their known distributions, this is the first evidence which confirmed its existence from the Quanzijie Formation. The bystrowianian chroniosuchian specimens from the base and the top of the Guodikeng Formation and the Jiucaiyuan Formation demonstrated that this group survived in the end-Permian mass extinction here in Xinjiang, together withLystrosaurus.
AcknowledgementsThanks to field crew to Xinjiang in 2018 and 2019, especially Liu Yu-Dong and Wang Ming-Li who discovered two of the studied specimens. FU Hua-Lin and Lan Li-Li prepared the specimens. Jozef Klembara and Igor V. Novikov kindly reviewed the paper. This work is supported by Strategic Priority Research Program of Chinese Academy of Sciences (XDB26000000), National Natural Science Foundation of China (41572019,41661134047), and National Natural Science Foundation of USA (NSF EAR 1714749).

- 古脊椎动物学报(中英文)的其它文章
- Ancient DNA molecular identification and phylogenetic analysis of Cervinae subfossils from Northeast China
- New skulls of ctenodactyloids from the Early Oligocene of Ulantatal, Nei Mongol, China
- Taxonomic revision of Anthracokeryx thailandicus Ducrocq, 1999 (Anthracotheriidae, Microbunodontinae)from the Upper Eocene of Thailand
- A new species of Luganoia (Luganoiidae, Neopterygii)from the Middle Triassic Xingyi Biota, Guizhou, China
- Subdivision and age of the Silurian fish-bearing Kuanti Formation in Qujing, Yunnan Province
- 总 目 次