
川西鹧鸪山典型林线杜鹃-冷杉林的物种多样性研究
2020-11-04 11:09李桾溢才晓源张远东缪宁
四川林业科技 2020年5期
李桾溢, 才晓源, 张远东, 缪宁*
1. 四川大学生命科学学院,生物资源与生态环境教育部重点实验室,四川 成都 610064;
2. 中国林业科学研究院森林生态环境与保护研究所,国家林业和草原局森林生态环境重点实验室,北京 100091
林线(Treeline)一般是指郁闭森林与树种线之间的生态过渡带[1]。对于林线的研究起源于极地林线的研究,最早可以追溯到1555年,Conrad Gessner对瑞士阿尔卑斯山前缘森林界限的描述[2]。对于高山林线,一般认为低温是其主要限制因素[1,3],森林所需要的水分无法满足也同样限制着森林的分布[4]。由于林线的特殊地理位置和极端环境条件以及对气候变化响应的敏感性[5-7],可以作为全球气候变化生物监测系统的理想“预警线”[8]。林线的位置、结构和动态对气候变化有着重要的意义,随着20世纪80年代左右的全球气候变暖,关于林线的研究成为研究热点,包括林线对气候变化的响应[9-13]、林线动态变化机制[14-16]、林线植被生理生态特征[17-21]、林线植物群落物种多样性[5,22-23]等。
目前,对林线植物群落物种多样性的研究集中在群落和种群特征研究[5,22-25]和物种多样性对气候变化的响应[13,26-28]。对林线群落和种群特征研究不同海拔梯度下林线物种多样性的变化[24,29]、林线交错带的边缘效应物种多样性研究[5,25]、不同坡向下林线物种多样性的变化[24]等。全球气候变暖可能导致林线及附近的生态系统的物种组成和群落结构变化[28],一方面是由于低海拔物种向高海拔迁移使得物种多样性增加[30-32],另一方面则是气候变暖使得原有的生物因生境变化而丧失[33]。但目前关于林线杜鹃-冷杉林的物种多样性的研究较少,本文旨在通过对林线杜鹃-岷江冷杉林的调查分析,了解林线环境下的物种多样性,为林线生物多样性研究和保护工作提供基础资料。
1 研究区概况
研究地点位于四川省阿坝藏族羌族自治州理县米亚罗镇的米亚罗林区的鹧鸪山垭口林线附近(N31°24′—31°55′,E102°35′—103°4′),海拔3 968 m。地貌类型为典型的高山峡谷,坡度多在30°以上。该区属于青藏高原气候区,受太平洋、印度洋及青藏高原三大气团影响,年降水量为600~1 100 mm,年蒸发量为1 000~1900 mm,1月均温为−8 ℃,7月均温为12.6 ℃,温度≥10 ℃年积温为1 200~1 400 ℃[34]。米亚罗林区植被垂直成带明显,原生森林分布于海拔2 400~4 200 m处,以亚高山暗针叶林为主,主要优势树种为岷江冷杉(Abies faxoniana)。
2 研究方法
2.1 样方设置
于2018年7月在米亚罗林区的林线附近的原始林中设置了一个100 m×100 m的样地,划分为25个20 m×20 m的乔木调查样方,记录每个样方中所有乔木物种的种名、高度、胸径以及数量等数据。将每个乔木调查样方划分为16个5 m×5 m的小样方(共400个小样方),并选取样地左上角到右下角对角线上的20个小样方和随机选取的48个样方作为灌木样方调查(共68个小样方),在样地左上角到右下角对角线上的20个和随机选取的16个灌木样方中选取大小为1 m×1 m的草本样方。记录每个样方中所有草本和灌木植物的种名、多度、盖度、高度等数据。
2.2 物种多样性测度方法
物种多样性作为群落的基本特征,既表征群落的组成结构,也是对环境状况的指示,分别反映了植物群落的不同方面的特点,选取的物种多样性测度方法如下[35]:
(1)物种丰富度(S):

(2) Shannon-wiener 多样性指数:

(3) Simpson 优势度指数:
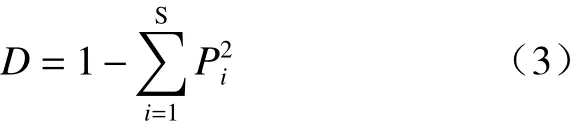
(4) Pielou 均匀度指数:
式中,Pi为种i的重要值,灌木和草本重要值的计算式为:

3 结果与分析
3.1 群落概况和物种组成
研究发现在100 m×100 m的样地中,共记录乔木、灌木与草本植物17 220株,其中乔木植物16 509株,灌木植物397株,草本植物314株。本次调查中共记录植物32种,分属24科,31属。乔木共3种,隶属于2科3属;灌木植物共8种,隶属于7科8属;草本植物共21种,隶属于18科21属。
乔木层郁闭度为0.47,乔木树种包括:岷江冷杉、理县杜鹃(Rhododendron trichogynum)和红杉(Larix potaninii)等。灌木层植物盖度较低,总盖度在0.06左右,最高总盖度为0.25,平均高度0.98 m,灌木植物包括:陕甘花楸(Sorbus koehneana)、冰川茶藨子(Ribes glaciale)、唐古特忍冬(Lonicera tangutica)、红 刺 悬 钩 子(Rubus rubrisetulosus)、红背杜鹃(Rhododendron rufescens)、香柏(Juniperus pingiivar. wilsonii)、柳属(Salixsp.)等。
草本层盖度为0.27,高于灌木层,近30%的样方总盖度达0.5以上,40%的样方总盖度在0.05以下,平均高度0.14 m。草本植物有:耳柄蒲儿根(Sinosenecio euosmus)、山酢浆草(Oxalis griffithii)、紫花碎米荠(Cardamine purpurascens)、洱源囊瓣芹(Pternopetalum molle)、微唇马先蒿(Pedicularis minutilabris)、葱状灯心草(Juncus allioides)、独叶草(Kingdonia uniflora)、丁座草(Boschniakia himalaica)、长鞭红景天(Rhodiolafastigiata)、石松(Lycopodium japonicum)、阴地蕨(Botrychium ternatum)、川甘火绒草(Leontopodium chuii)、弯齿风毛菊(Saussurea przewalskii)、糙野青茅(Deyeuxia scabrescens)、东方草莓(Fragaria orientalis)、林地早熟禾(Poa nemoralis)、蓼属(Polygonumsp.)、蹄盖蕨属(Athyriumsp.)、堇菜属(Violasp.)、莎草属(Cyperussp.)和石竹科(Caryophyllaceae sp.)等。苔藓植物分布较为均匀,盖度0.9,厚度约为9 cm。林内长松萝(Usnea longissima)附生现象明显,密布于岷江冷杉枝条上[1]。
3.2 物种多样性
乔木层以岷江冷杉和理县杜鹃为优势种,重要值分别为49.72%和46.96%,红杉的重要值仅占3.33%;灌木层以陕甘花楸为优势种,重要值占灌木层的57.79%,且75%以上的植物重要值在10%以下;草本层相对优势的物种为山酢浆草和糙野青茅,重要值较低,仅为28%和25.94%,且85%以上的植物物种重要值在10%以下(见表1−3)。

表1 乔木树种的重要值Tab.1Importance value of tree species
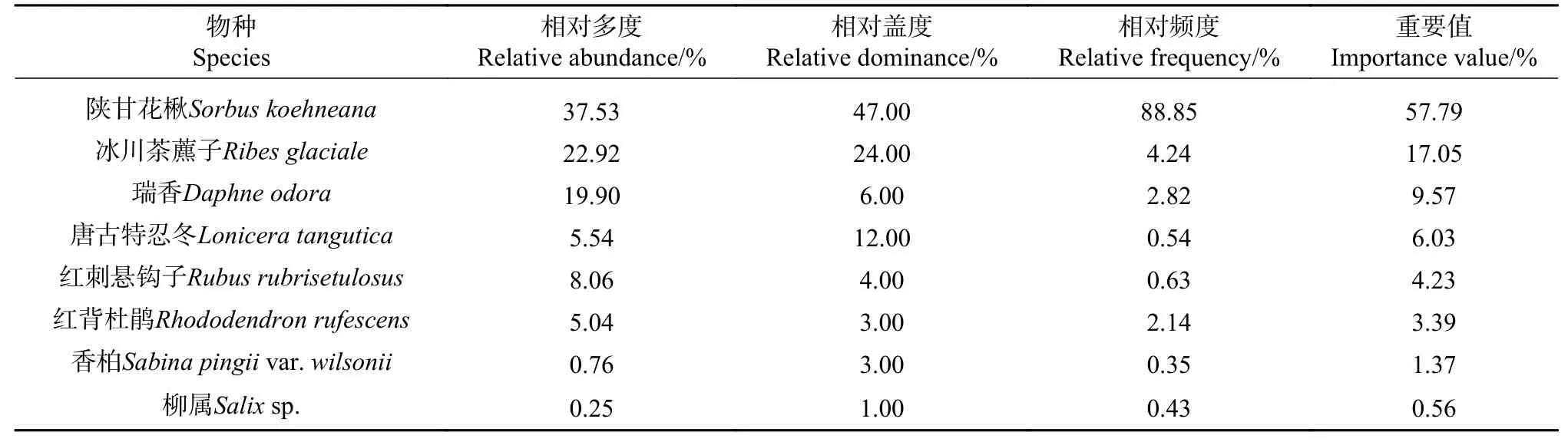
表2 灌木植物的重要值Tab.2Importance value of shrub species
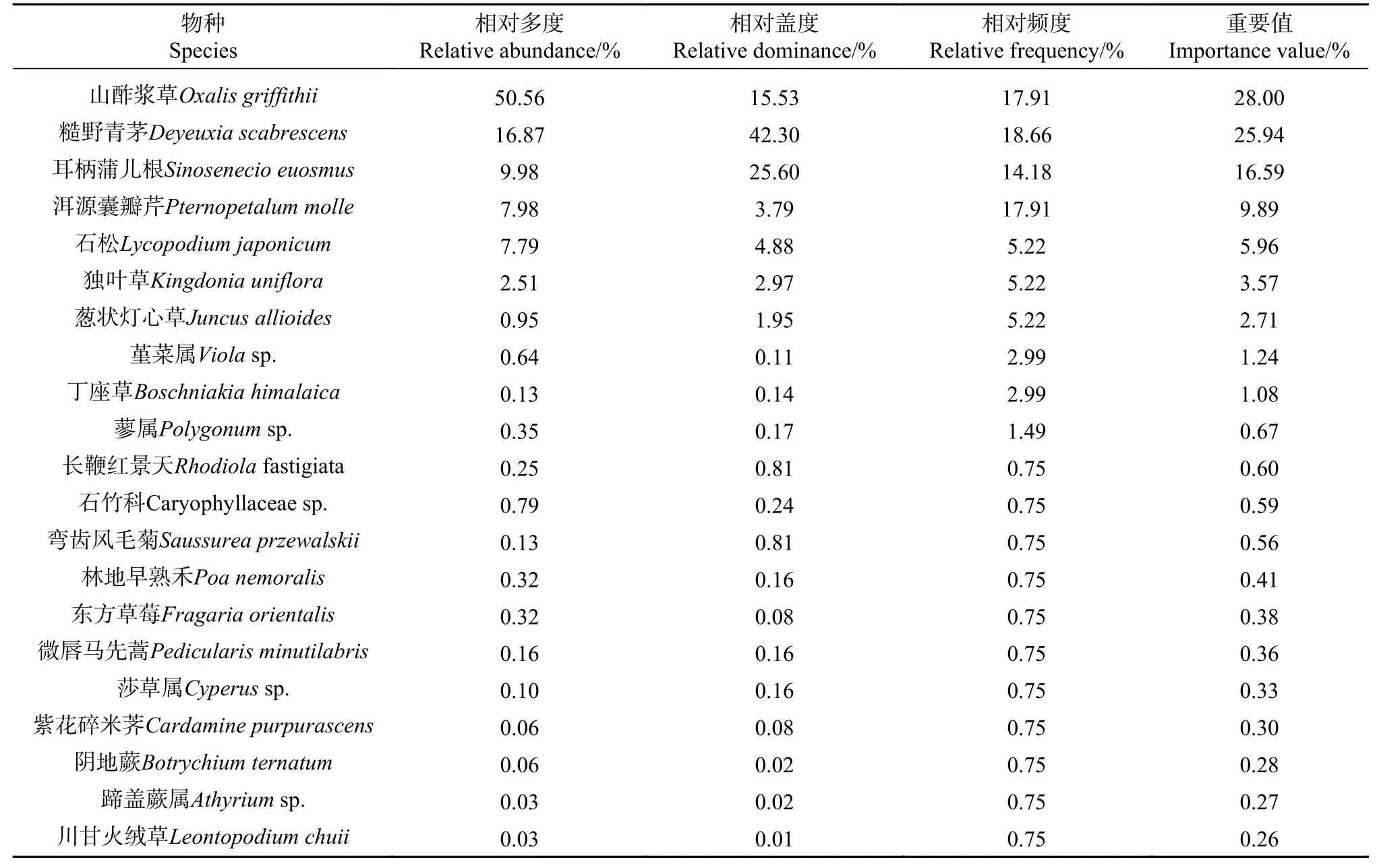
表3 草本植物的重要值Tab.3Importance value of herb species
在杜鹃-岷江冷杉林中,乔木层中属于A级频度的物种占33.33%;无B、C和D级频度的物种;属于E级频度的物种,即优势种和建群种的占比为66.67%。灌木层中属于A级频度的物种占全部灌木的75.00%;B和D级频度的种类较少,均占12.50%;无C和E级频度的物种。草本层中属于A级频度的物种占全部草本植物的80.95%;C级的物种较少,占4.76%;属于D级频度的物种,占全部草本植物的14.29%;无B和E级频度的物种(见图1)。由于群落均匀度与E级的比例成正比,乔木层、灌木层和草本层对比来看,乔木层的均匀度更高。灌木层和草本层的频度分布属于A级频度的物种最多的,B、C和D频度级的物种较少,表明物种分布较为均匀。

图1 乔木层、灌木层和草本层物种的频度分布Fig.1Frequency distribution of tree species, shrub species, and herb species
在林线附近的岷江冷杉-杜鹃林中,Shannon-Wiener指数、物种丰富度和Simpson优势度指数最大的均为草本层(H′= 1.99,S= 21,D= 0.81),其次为灌木层(S= 8,H′= 1.35,D= 0.62),乔木层最小(S=3,H′= 0.82,D= 0.53),乔木层、灌木层和草本层的Shannon-Wiener指数和物种丰富度表明在岷江冷杉-杜鹃林中,草本层的丰富度和群落复杂程度最高,灌木层其次,乔木层最小;且相比草本层和灌木层,乔木层的优势种的地位最突出。乔木层Pielou均匀度指数最大(J= 0.74),草本层和灌木层的Pielou均匀度指数相似(J= 0.65)(见表4),表明乔木层物种分布最均匀,草本层和灌木层物种分布的均匀度相似。
4 讨论
在林线杜鹃-岷江冷杉林中,乔木层的优势种岷江冷杉和杜鹃二者重要值之和高达96%,陕甘花楸的重要值也在灌木层中超过一半,这与林线的极端环境相关,符合Whittaker提出的生态位优先占领假说(Niche-preemption hypothesis)[36],即,第一位优势种首先占领主要的空间生态位,第二位的占领其余的大部分空间,依此类推,到末位的只能占留下的很少空间。而草本层以山酢浆草和糙野青茅为优势种,但重要值较乔木和灌木优势种偏低分别为28.00和25.94,林线乔、灌、草各层次的物种组成和数量相对稳定,频率分布均较为均匀(见图1,表4),与石培礼[5]等在四川省卧龙巴朗山林线的发现相似,岷江冷杉林线交错带群落生境相对均匀,草本层优势种及其地位不明显。这是由于草本层比乔木层和灌木层的物种组成更丰富,重叠的生态位以及种间竞争抵消了优势种的地位。

表4 岷江冷杉-杜鹃林各层次物种多样性指数Tab.4Species diversity index of different growth form layers in the Abies faxoniana-Rhododendron forest
与同在米亚罗林区的28~33年生的云杉(Picea asperata)天然林(海拔3 179~3 270 m)相比[37],岷江冷杉-杜鹃天然林的乔木层和灌木层Shannon-Wiener指数都较小,草本层的Shannon-Wiener指数较高;乔木层的Simpson优势度指数偏低,灌木层和草本层Simpson优势度指数均与云杉天然林相似。由于云杉天然林的海拔较低,其水热条件较林线处的环境更好,使得云杉天然林的乔木层和灌木层物种更为丰富,而由于乔木层和灌木层的竞争作用抑制了草本层的发育。岷江冷杉-杜鹃天然林的不同层次的Pielou均匀度指数均较云杉天然林更高,说明林线各层次物种空间分布更为均匀。
荒漠地区和林线环境均属于极端环境,整体的生物多样性均处于较低水平,但也有差异。与阜康北部荒漠的梭梭(Haloxylon ammodendron)群落和白梭梭(Haloxylon persicum)群落的灌木层相比[38],岷江冷杉-杜鹃天然林的灌木层物种更多,Shannon-Wiener指数和Pielou均匀度指数均更高,Simpson优势度指数:梭梭群落(D=0.95)>岷江冷杉-杜鹃林>白梭梭群落(D=0.37)。说明岷江冷杉-杜鹃天然林灌木层的生物多样性较高,空间分布较梭梭群落和白梭梭群落更为均匀,群落更为稳定。但内蒙古额济纳绿洲的天然胡杨(Populus euphratica)林成熟林草本层(H'=0.21,D=0.21)与灌木层(H'=0.24,D=0.14)的物种多样性均小于林线处岷江冷杉-杜鹃天然林[39]。与胡杨林相比,在整体上生物多样性均较低,乔、灌 、草三个层次植物种数均不多,其中,都以灌草层植物种数占绝对数量优势。这与胡杨林分布于干旱少雨、冬季寒冷干燥、土壤盐分较高的生境[40],而位于米亚罗林区林线附近的岷江冷杉-杜鹃林的水分条件较胡杨林的荒漠地区更好相关。
同样位于林线条件下,西藏色季拉山林线阴坡的杜鹃-急尖长苞冷杉(Abies georgeivar.smithii)林(海拔4 320~4 390 m)灌木层(H′= 0.818,D=0.489)和草本层(H′= 0.654,D= 0.466)物种多样性指数均小于位于米亚罗林线的杜鹃冷杉林的草本层和灌木层[24]。同样位于阴坡的苔藓急尖长苞冷杉林(海拔4 000~4 320 m)灌木层(H′= 2.095,D= 0.874)物种多样性指数较米亚罗林线的杜鹃冷杉林更大,草本层(H′= 1.026,D= 0.624)则更小,这可能是由于苔藓急尖长苞冷杉林中的苔藓盖度和高度较高,抑制了草本层的发育生长[24],而灌木层不易受苔藓层的影响;与之相反,米亚罗林线的杜鹃冷杉林的草本层与灌木层的相互竞争使得该灌木层的物种多样性低于色季拉山林线的苔藓急尖长苞冷杉林的灌木层。而由于位于阳坡林线的方枝柏(Sabina saltuaria)+急尖长苞冷杉林(海拔4 200~4 320 m)和杜鹃-方枝柏林(4 320~4 570 m)的林下光线充足,其灌木层(方枝柏+急尖长苞冷杉林:H′= 2.202,D= 0.889;杜鹃-方枝柏林:H′= 2.272,D= 0.893)和草本层(方枝柏+急尖长苞冷杉林:H′= 2.616,D=0.915;杜鹃-方枝柏林:H′= 3.519,D= 0.975)的物种多样性均高于米亚罗林线的杜鹃-岷江急尖长苞冷杉林[24]。
5 结论
本次在米亚罗鹧鸪山林线处1 hm2的杜鹃-冷杉林中调查中共记录植物32种分属24科31属。调查样地区域整体呈现出物种多样性低,灌木、草本植物种类稀少的特征,与青藏高原东缘原始林林线附近脆弱、敏感的生态环境相关。其中,乔木层的物种多样性指数较低,而灌木和草本植物的物种多样性则较高,且各层的物种多样性空间分布较为均匀。
猜你喜欢
散文诗世界(2021年12期)2021-12-17
青年文学家(2021年16期)2021-08-05
少儿美术(2020年8期)2020-11-06
林业与生态(2019年9期)2019-10-07
岷峨诗稿(2019年4期)2019-04-20
花火A(2019年1期)2019-03-22
现代养生·上半月(2017年7期)2017-07-07
发明与创新·大科技(2017年6期)2017-06-20
廉政瞭望(2016年13期)2016-08-11
中华奇石(2015年12期)2015-07-09
