
伐藤对五指山市低海拔次生林群落特征的影响
2020-11-03 07:55林承博陈国德张何信符生波杜尚嘉童思远余雪标
中南林业科技大学学报 2020年10期
林承博,陈国德,张何信,符生波,杜尚嘉,童思远,余雪标
(1.海南大学 林学院,海南 海口 570203;2.海南省林业科学研究院,海南 海口 571100)
藤本植物作为一类必须依靠其他植物或其他支持物的物理支撑向上攀援的植物[1],是热带地区森林的重要组成部分之一,在受干扰的生境(如次生林)中多度更大[2-4]。因为生态特性与林木不同,藤本植物自身不能构成群落,必须依附他物而生,导致其生长环境变化不定,由于生长过程受到支持木的限制,藤本植物经常在林冠攀援或在林下匍匐生长[5-9],其独特的生物学特性,常使它们在生态适应性和种间竞争上具有明显优势[1]。部分藤本植物特别是一些大型木质藤本的蔓延扩散能显著滞缓甚至改变次生林的演替进程[10-11]。
目前对于热带森林藤本植物的研究较少,刘奇等[4]通过调查西双版纳热带雨林,报道了该地区木质藤本的多样性、径级分布、攀援方式及对支持木的攀援情况;赵科等[12]通过对海南霸王岭原始山地雨林区木质藤本垂直结构特征的调查,发现支持木的高度是影响木质藤本多度、丰富度和物种组成的重要因素;刘晋仙等[13]调查了霸王岭热带山地原始林与伐后林中支持木与攀附在其上的藤本植物,证明了木质藤本对支持木在科和种的水平上具有选择性。目前的研究主要集中在热带雨林地区,而对于次生林研究较少。
五指山市低海拔次生林群落处于演替的初级阶段[14],由于频繁遭受台风的侵袭以及严重的人为干扰等因素,森林生态系统的抵抗力和稳定性较差。该地区的藤本植物呈爆发式增长,其中以金钟藤、葛等为最。随着全球气候变暖,藤本植物的多度将呈增加的趋势,必然会影响共存的乔灌草的生长、死亡、多样性等[4,15-17]。本研究以五指山市低海拔次生林为研究对象,设置3 种伐藤处理与一组对照,分析群落特征、物种多样性以及藤本植物的群落特征与乔灌草多样性的相关性,了解藤本植物对整个森林群落特征的影响,为是否需要对五指山地区的藤本植物进行防治提供参考,为推动五指山地区次生林的恢复提供理论基础。
1 研究区概况与研究方法
1.1 研究区概况
研究区位于海南省五指山市畅好乡(18.71°~18.73°N,109.47°~109.49°E),属热带季风气候,海拔高度360~480 m。该地区森林资源丰富,以倒吊笔Wrightia pubescens、琼榄Gonocaryum lobbianum、银柴Aporosa dioica、乌心楠Phoebe tavoyana等常绿阔叶树种为主,冬春干旱,夏秋多雨,雨旱分明,雨季为5—10月,旱季从11月至翌年4月。兼有台风,秋末日照少,日夜温差大;年活动积温8 166.2~8 411.5 ℃,年平均气温22.7 ℃,夏季平均气温25.9 ℃,冬季平均气温17 ℃,年极端最高气温37.1 ℃,年极端最低气温0.1 ℃;年平均日照时数2 122.6 h;年平均降水量1 771.8 mm,最大年降水量2 810.4 mm,最小降水量1 055.5 mm[8,14]。
1.2 样地设置与调查
通过实地踏查等方式,在研究区挑选3 块藤本植物干扰较为严重的次生林群落并进行不同的伐藤处理,分别记为:未伐藤(未伐藤的次生林群落);伐藤2 a(人工伐除样地内所有藤本植物,自然恢复2 a 的次生林群落);伐藤4 a(伐藤后自然恢复4 a 的次生林群落);另在附近选择立地条件相近的未被藤本植物覆盖的次生林群落作为对照。每块样地布设3 块总计12 块20 m×20 m的样方,每块样地的坡位、坡向及海拔高度等立地条件基本一致。用样方法对样地内的乔木、灌木、草本、匍匐藤本、攀援藤本[18-20]进行调查,记录每一物种名称。
将株高大于1.5 m、胸径大于2 cm 的树种记为乔木;株高大于0.5 m 小于1.5 m 记为灌木;株高小于0.5 m 记为草本;倾斜度>45°、高度>1.5 m 的含有主干的死亡木记为枯立木[21],记录枯立木的径粗和株高。将藤本植物分为匍匐藤本和攀援藤本,在地上匍匐生长的、还未攀附在支持木上或攀附高度低于30 cm 的藤本植物记为匍匐藤本,记录其在样方内的长度,并测量径粗;攀附在支持木上且攀附高度大于30 cm 的藤本植物记为攀援藤本,测量其攀爬高度及地径。
1.3 群落特征值与物种多样性计算
群落特征包括种类特征(物种数、优势种、重要值等)和乔木层树高、胸径、冠幅、多度;灌木层株高、盖度、冠幅、多度;草本层多度、盖度、高度;匍匐藤本的平均径粗、多度、长度;攀援藤本的地径、多度、攀援高度。所有林分群落特征指标均取平均值,在Excel 中计算不同样方间的平均值与标准偏差。
区分4 种伐藤处理次生林群落的乔木层、灌木层、草本层、匍匐藤本、攀援藤本,物种多样性采用马克平等[22-23]的重要值、丰富度指数、多样性指数及均匀度指数计算公式进行计算。

式中:Pi为第i种的个体数占所有种个体数的比例;S为种i所在样方中物种总数。
1.4 数据分析
运用SAS 9.2 分析群落特征和物种多样性指数的差异显著性和藤本植物的群落特征与乔灌草物种多样性之间的相关性。
2 结果与分析
2.1 不同伐藤处理次生林的群落特征
2.1.1 不同伐藤处理乔灌草的种类特征
研究区域次生林群落乔、灌、草层共出现植物212 种,隶属于86 科166 属,其中,大戟科Euphorbiaceae 种数最多,为17 种;其次是茜草科Rubiaceae,有13 种;樟科Lauraceae 11 种。乔木层植物共102 种,分属于49 科86 个属,其中,樟科和大戟科种数最多,各为9 种,其次是芸香科Rutaceae 6 种,茜草科5 种,无患子科Sapindaceae 5 种。不同伐藤处理次生林群落中乔灌草优势种如表1所示。未伐藤的次生林有乔木31 种,分属于22 科29 属,倒吊笔是重要值最大的物种,其次是琼榄、猫尾木、无患子、破布叶;倒吊笔、猫尾木、无患子等阳性树种重要值之和为0.719,因此未伐藤的次生林群落乔木层以阳性树种为主。伐藤2 a的次生林乔木32 种,分属于23 科31 属,重要值最大的物种依次为琼榄、银柴、龙眼、破布叶、黄牛木,其中银柴、龙眼、黄牛木等阳性树种所占比例减小为0.556;伐藤4 a 的次生林中有乔木55 种,隶属于45 科49 属,重要值由大到小分别为银柴、琼榄、乌心楠、厚皮树、山柑算盘子,其中银柴、厚皮树等阳性树所占比例为0.536;对照的次生林中乔木共67 种,分属于33 科53 属,优势种则变为了乌心楠、黄椿木姜子、美叶菜豆树、海南锥、银柴,其中银柴、厚皮树等阳性树种所占比例为0.512,阳性树种占比继续减少,阴性和中性树种逐渐增加。
灌木层共出现110 种植物,隶属于48 科88 属。其中,大戟科11 种为最多,其次是茜草科8 种,无患子科6 种,樟科6 种。未伐藤的次生林中灌木共39 种,共计25 科35 属,重要值最大的5 个种依次是破布叶、心叶黄花稔、苎麻、倒吊笔、鲫鱼胆,阳性树种重要值之和为0.397;伐藤2 a的次生林有28 种灌木,分属于19 科27 属,优势种为大青、银柴、倒吊笔、破布叶、草豆蔻,阳性树种重要值之和为0.476;伐藤4 a 的次生林灌木层物种数为52,分属于25 科46 属,优势种依次为银柴、琼榄、牛眼睛、假黄皮、猪肚木,阳性树种重要值之和为0.4;对照的次生林灌木层物种数为43,分属于25 科40 属,重要值从大到小依次是九节、黧蒴锥、糖棕、苎麻、乌心楠,阳性树种重要值之和为0.367。
草本层共有86 种植物,隶属于50 科76 属。其中,菊科Asteraceae 8 种、茜草科8 种为最多,其次是大戟科6 种,禾本科5 种、桑科4 种。未伐藤的次生林中草本植的物种数为33,分属于21科31 属,重要值最大的5 个种依次是马唐、心叶稷、苎麻、柔枝莠竹、飞机草;伐藤2 a 的次生林出现草本植物29 种,分属于17 科28 属,优势种为马唐、藿香蓟、心叶稷、飞机草、柔枝莠竹;伐藤4 a 的次生林中出现草本植物38 种,分属于27 科37 属,优势种依次为马唐、银柴、飞机草、猪肚木、九节;对照的次生林中草本植物的物种数较少,仅18 种,分属于15 科16 属,重要值从大到小依次是莎草蕨、淡竹叶、糖棕、九节、狭叶凤尾蕨。
2.1.2 不同伐藤处理藤本植物的种类特征
本次调查的样方中,匍匐藤本的物种数为48,分属35 科39 属。其中,旋花科Convolvulaceae和茜草科各4 种为最多,其次是豆科Leguminosae、百合科Liliaceae 均为3 种。不同伐滕处理次生林群落中藤本植物优势种如表2所示。未伐藤的次生林中匍匐藤本物种数为10,分属于8 科8 属,重要值最大5 个种依次是金钟藤、葛、榄形风车子、山蒟、畏芝;伐藤2年的次生林中有17 种匍匐藤本,分属于13 科16 属,优势种为葛、榄形风车子、金钟藤、海南崖豆藤、掌叶鱼黄草;伐藤4 a 的次生林中匍匐藤本的物种数为19,分属于14 科17属,优势种依次为葛、金钟藤、榄形风车子、华马钱、山蒟;对照的次生林中的匍匐藤本的种类数为29,分别属于24 科24 属,重要值从大到小依次是锡叶藤、葛、菝葜、木防己、金钟藤。各种处理组中,金钟藤、葛、榄形风车子重要值均较高,仅对照组中匍匐的榄形风车子重要值较低,为0.002,未伐藤、伐藤2 a、伐藤4 a、次生林中金钟藤、葛、榄形风车子重要值之和分别为0.940、0.844、0.606、0.236。

表1 不同伐藤处理次生林群落中乔木层、灌木层、草本层优势种(重要值最大的5 个种)Table 1 The dominant species in the arbor, shrub, herb of the secondary forest community under different recovery time after eliminate lianas (IV≥5)
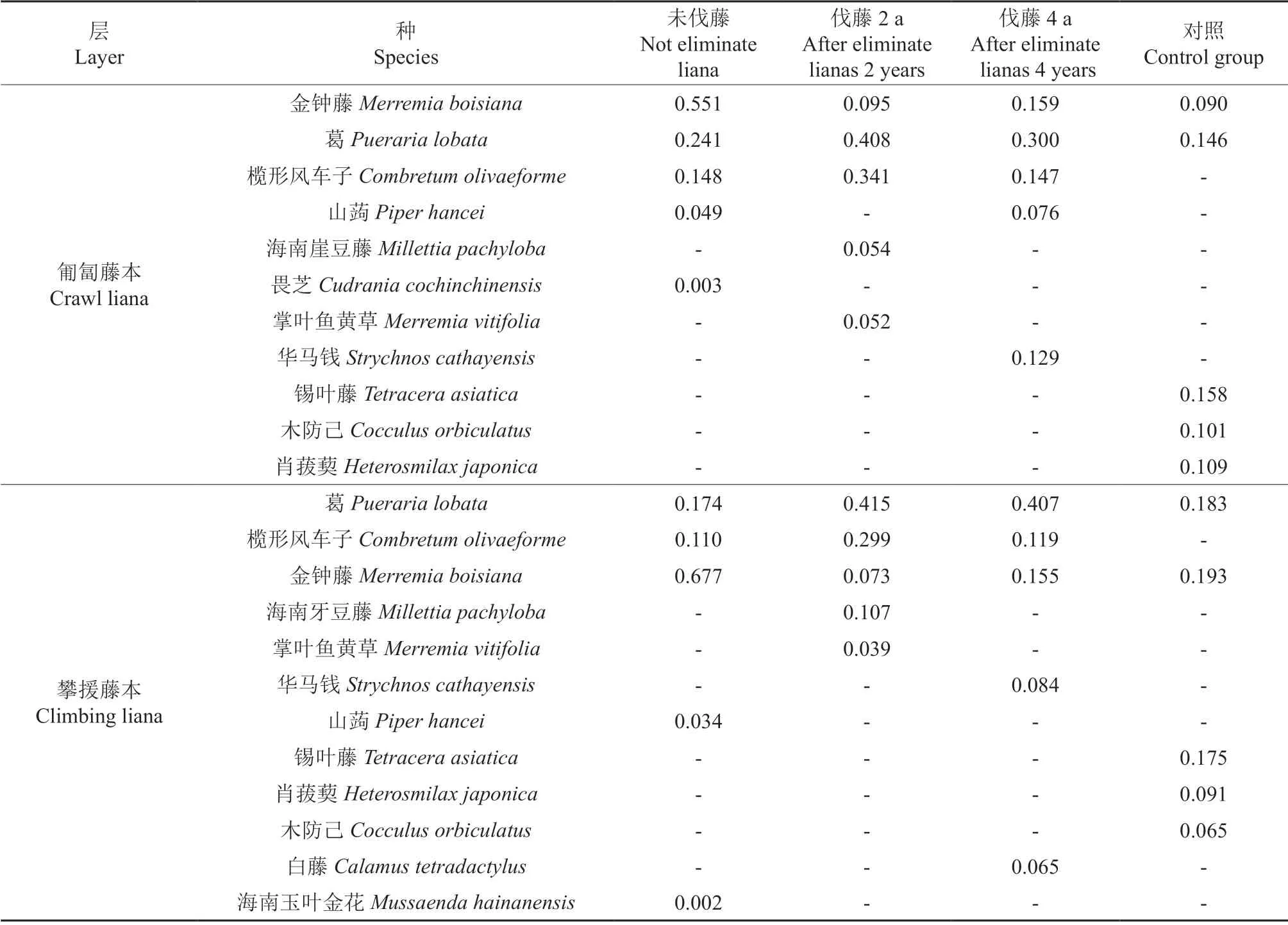
表2 不同伐藤处理次生林群落中藤本植物优势种(重要值最大的5 个种)Table 2 The dominant species in the lianas in the secondary forest community under different recovery time after eliminate lianas (IV≥5)
攀援藤本的物种数为46,分属34 科38 属。其中,旋花科4 种、茜草科4 种并列最多,其次是豆科3 种,百合科Liliaceae 3 种。未伐藤的次生林内攀援藤本有9 种,分属于6 科6 属,重要值最大的5 个种依次是金钟藤、葛、榄形风车子、山蒟、海南玉叶金花;伐藤2 a 的次生林中出现的攀援藤本共计13 种,分属于9 科12 属,优势种为葛、榄形风车子、金钟藤、海南崖豆藤、掌叶鱼黄草;伐藤4 a 的次生林内出现的攀援藤本的物种数为19,分属于15 科15 属,优势种依次为葛、金钟藤、榄形风车子、华马钱、白藤;对照组的次生林中攀援藤本的物种数最多,为29 种,分属于22 科23 属,重要值从大到小依次是金钟藤、葛、锡叶藤、菝葜、木防己。4 种伐藤处理的次生林群落中金钟藤、葛、榄形风车子重要值也较高,3 种藤本植物的重要值之和表现为未伐藤(0.961)、伐藤2 a(0.787)、伐藤4 a(0.681)、次生林(0.376)。伐藤前,金钟藤重要值最大;伐藤后,优势种变为葛。
2.1.3 不同伐藤处理乔木层枯立木
不同伐藤处理下次生林群落中乔木层的枯立木胸径和多度见表3。由表3可知,伐藤2 a 的次生林群落中枯立木多度略少于未伐藤的次生林群落,两者间差异不显著;枯立木胸径呈未伐藤>伐藤2 a>伐藤4 a,三者间差异极显著,说明伐藤后,大量小径阶的乔木死亡。

表3 乔木层枯立木(平均值±标准差)†Table 3 Snags of arbor(Mean ± SD)
2.1.4 不同伐藤处理乔木层、灌木层和草本层特征差异比较分析
不同伐藤处理的样方之间群落特征存在着差异,表4分别对4 种处理的样地乔木层、灌木层、草本层、匍匐藤、攀援藤的特征值进行比较。
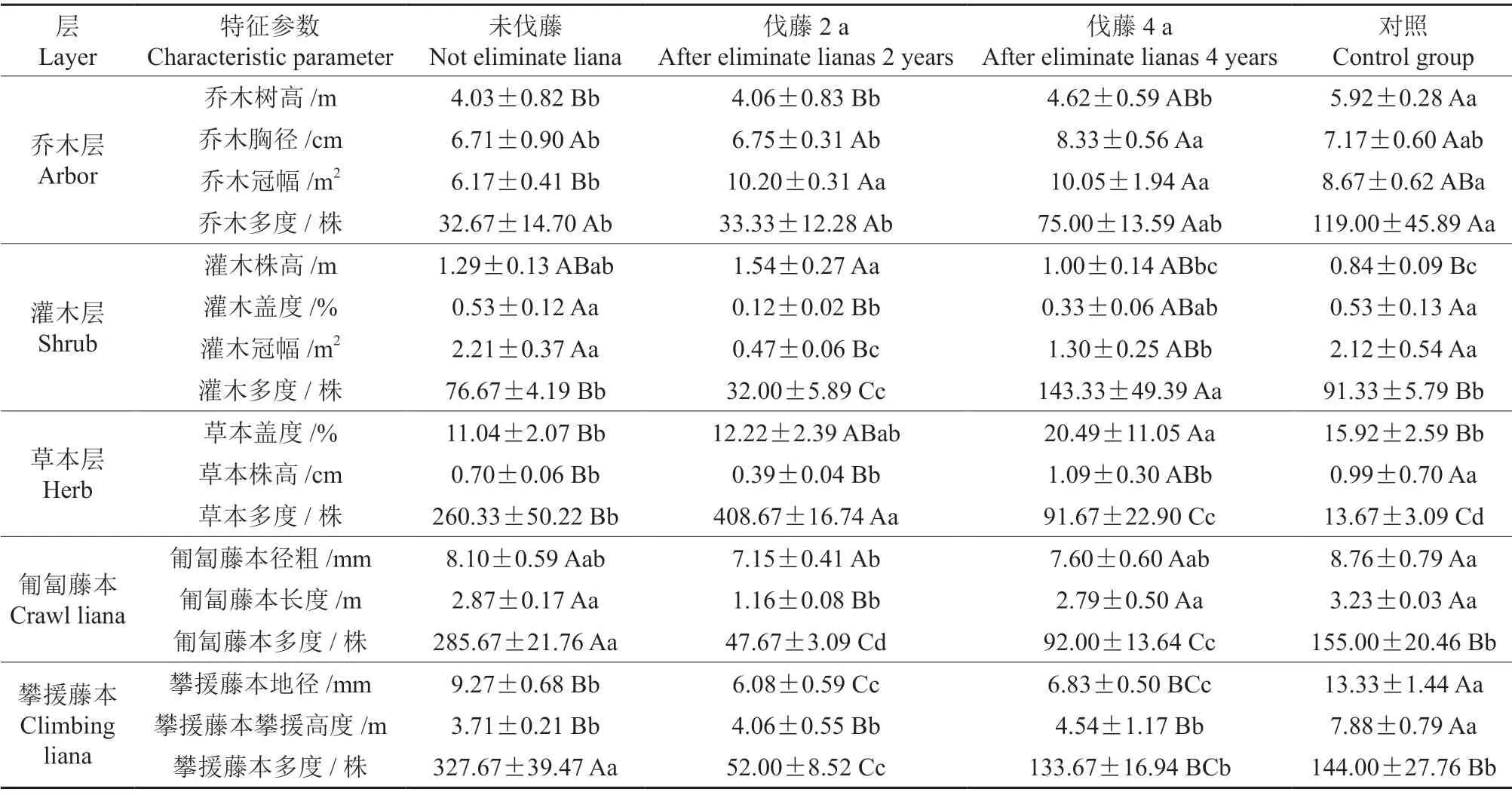
表4 不同伐藤处理下次生林群落特征值(平均值±标准差)†Table 4 The community characteristic values of the secondary forestcommunity under different recovery time after eliminate lianas (Mean±SD)
乔木层的平均树高、多度表现对照最大,未伐藤最小,且差异达到显著水平(P<0.05)。
灌木层的多度表现为伐藤4 a>未伐藤>伐藤2 a,且差异极显著(P<0.01),即伐藤后,次生林中的灌木表现为先减少再增多的趋势。
草本层的多度表现为伐藤2 a>未伐藤>伐藤4 a>次生林,4 种伐藤处理的差异极显著。
匍匐藤本的多度表现为未伐藤>对照>伐藤4 a>伐藤2 a,各种处理间多度差异显著。
攀援藤本的多度、径粗表现出与匍匐藤本相同的变化趋势;攀爬高度因为支持木的关系表现出与乔木层的树高相一致的规律,即对照显著多于其他3 种处理。
2.2 不同伐藤处理次生林群落物种多样性特征比较分析
表5为不同伐藤处理次生林群落的物种多样性指数的比较。
与未伐藤相比,随着恢复年限的增加,伐藤后次生林群落乔木层的Margalef 物种丰富度指数(RT)、Simpson 指数(DT)、Shannon-Wienner指数(HT′)、Pielou 指数(JT)均出先增长,但都低于对照。
对照灌木层的Margalef物种丰富度指数(S-R)、Shannon-Wienner 指数(S-H′)显著多于其它3 种次生林。
草本层的Margalef 物种丰富度指数(H-R)、Simpson指数(DH)、Shannon-Wienner 指数(H-HH′)、Pielou 指数(H-J)均表现为对照最大,从未伐藤到伐藤4 a,草本层的各项指数均出现了显著增长。
匍匐藤本的Margalef 物种丰富度指数(RCrL)、Simpson 指数(DCrL)、Shannon-Wienner 指数(HCrL′)均随着伐藤年限的增长而增长,但都显著低于对照。
与匍匐藤本相同,攀援藤本的的Margalef 物种丰富度指数(RClL)、Simpson 指数(DClL)、Shannon-Wienner 指数(HClL′)、Pielou 指数(JClL)也随着恢复时间的增加而增加,伐藤4 a>未伐藤,且都显著低于对照。
2.3 藤本植物的群落特征与次生林群落物种多样性的相关性
将乔灌草的物种多样性、丰富度指数、均匀度指数与藤本植物的群落特征进行相关性分析,结果如表6所示。
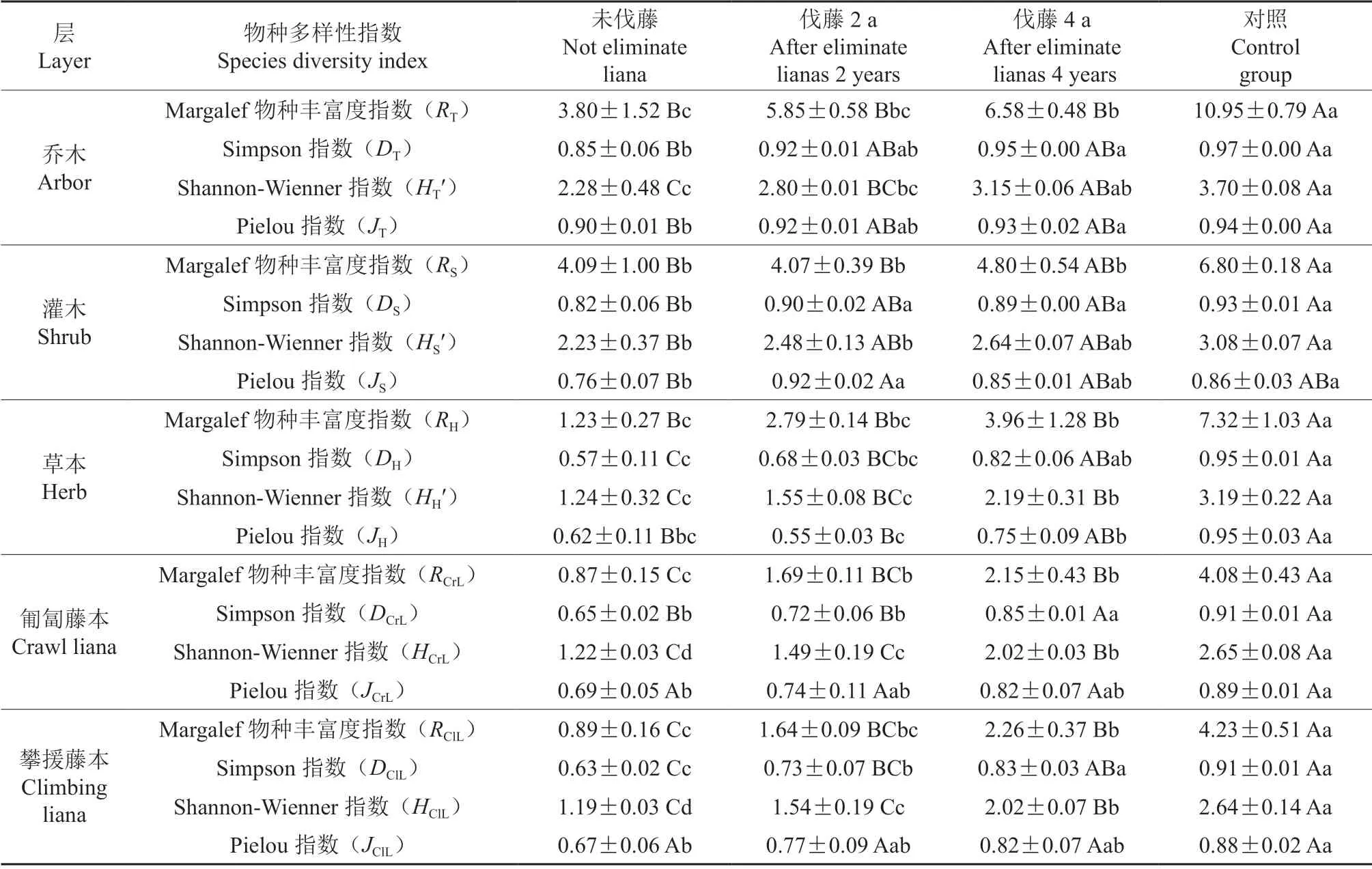
表5 不同伐藤处理次生林群落的多样性指数、均匀度和丰富度指数(平均值±标准偏差)Table 5 Richness, diversity and evenness indexes of the secondary forest community under different recovery time after eliminate lianas (Mean±SD)
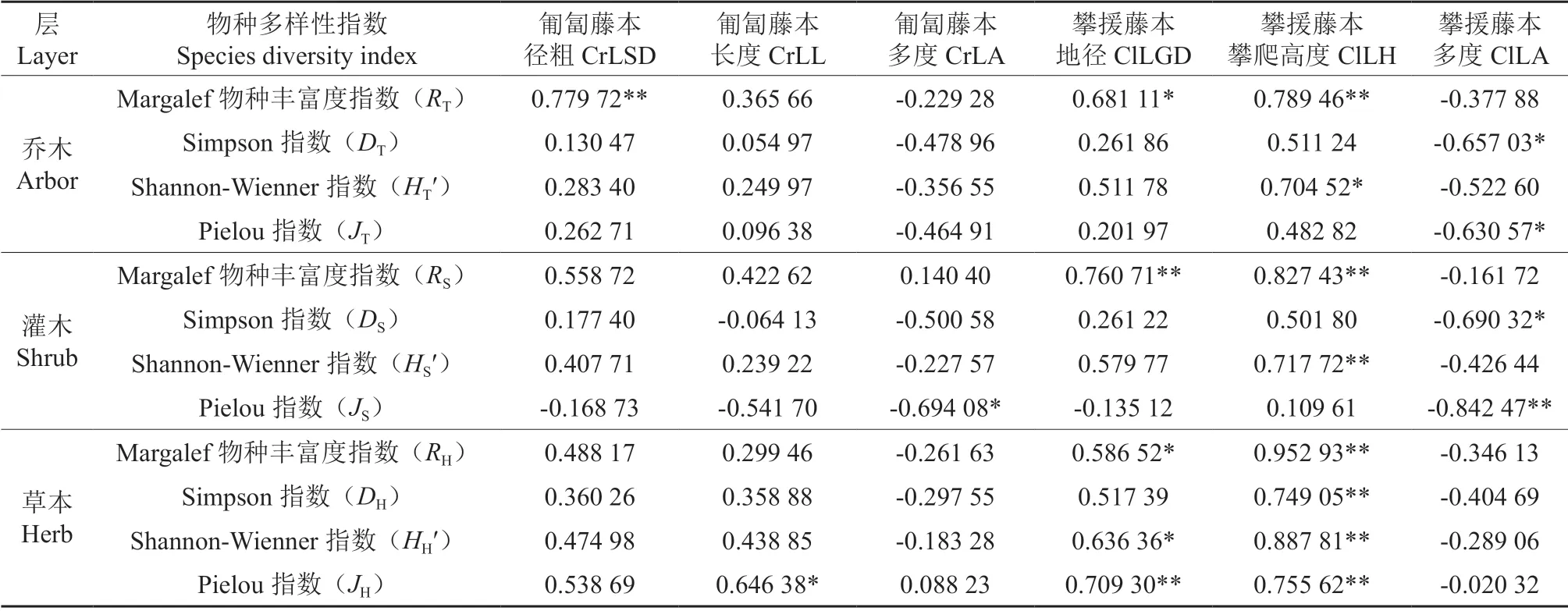
表6 藤本植物的群落特征与乔灌草物种多样性的相关性†Table 6 The relevance of lianas’ community characteristics with diversity indices of the arbor, shrub and herb
乔木层的Margalef 物种多样性指数RT与攀援藤本的地径(ClLGD)、攀爬高度(ClLH)呈较强的正相关关系;攀援藤本的攀爬高度(ClLH)与Shannon-Wienner 指数HT′呈正相关关系;攀援藤本的多度与Simpson 指数DT、Pielou 指数JT呈较强的负相关关系。
灌木层与乔木层相似。
攀援藤本的地径和攀爬高度与草本层的各项物种多样性指数、丰富度指数、均匀度指数呈较强的正相关关系。
3 结论与讨论
3.1 五指山市低海拔次生林的群落特征
在本次调查的4 种次生林群落的攀援藤本和匍匐藤本中,金钟藤、葛、榄形风车子的重要值均较大,伐藤后,重要值最大的藤本植物由伐藤前的金钟藤变为葛。在对不同次生林枯立木的多度和胸径调查中发现伐藤后大量小径阶乔木死亡。伐藤后,乔木层、灌木层多度出现先减少再增多的趋势,草本层则呈现先增多再减少的趋势。4 种次生林群落内阳性树种的重要值之和占比较大,随着伐藤后次生林的恢复,乔木层和灌木层的阳性树种重要值之和都出现降低的趋势。
森林群落的演替是以群落结构的变化为表现特征的,并首先表现为种类结构的变化,因而可根据群落种类结构变化来对群落演替现状进行判断[24-25]。程汉亭等[26]研究发现金钟藤种子的自然萌发率较低,而葛种子在雨水充足、温度适宜的情况下比较容易自然萌发[27],因此本调查中发现伐藤后葛的重要值超过金钟藤的主要原因或与其种子萌发率有关。赵俊卉等[28]认为耐荫树种依赖于上层林木对温度和湿度等的调节,保留大量上层林可以促进阴性树种的发育,Webster 等[29]认为单木择伐形成的林隙会导致耐阴性树种的多样性降低,伐藤后产生的林窗一定程度上导致了部分小径阶阴性树种的死亡,从而使伐藤后的次生林中小径阶枯立木增多。适度择伐有助于森林更新和多样性恢复,因此乔、灌层呈增加状态,次生林群落逐渐恢复,随之郁闭度逐增加,草本层呈现出减少的状态,农友等[30]也认为适当间伐有助于林下植被恢复和林木自身生长,而郁闭度过大时则会导致草本层呈现消亡趋势。4 种次生林群落内阳性树种的重要值之和占比均较大,表明该区域的次生林群落处于以阳性树种为主的长绿阔叶林阶段,即演替的初级阶段。随着伐藤后的恢复,阴性和中性树种所占比例逐渐增加,表明对于被藤本植物干扰较为严重的的次生林群落来说,藤本植物的大量繁殖使得次生林群落出现了一定程度的退化,伐藤推动了森林的正向演替。
3.2 物种多样性比较分析
伐藤后,次生林群落中乔木、灌木、草本、匍匐藤本、攀援藤本的物种丰富度指数、多样性指数大都出现了显著增长,未伐藤次生林群落的各项指标均为最低,随着伐藤后恢复时间的增加,物种丰富度指数和物种多样性指数均随之增长。
童思远等[31]在对不同伐藤处理下次生林群落土壤种子库的特征及演替趋势研究中发现,在次生林群落土壤种子库中的种子密度和丰富度上,做伐藤处理的明显高于未做伐藤处理的,本调查的结果也显示伐藤后物种丰富度指数和多样性指数显著增长,说明伐藤处理一定程度上确实可以促进次生林的物种多样性。而伐藤后的样地内开始渐出现新物种,丰富度指数和物种多样性指数随之增大,更有力地说明,对于藤本数量过多的次生林群落,伐藤处理能够对增加次生林群落的物种丰富度、多样性,增大复杂程度,增强次生林群落的稳定性等起到明显的推动作用。
3.3 相关性分析
除群落特征和物种多样性外,调查还发现攀援藤本的群落特征与乔灌草的物种多样性之间存在明显的相关关系:攀援藤本的地径和攀爬高度与乔灌木的Margalef 物种丰富度指数呈正相关,攀援高度与乔灌木Shannon-Wienner 指数呈正相关;攀援藤本的多度则与乔灌木的Simpson 指数和Pielou 指数表现为明显的负相关;攀援藤本的地径和攀爬高度的增加对草本层的各项指数表现为正相关。
颜立红等[32]在对藤本植物与支持木之间的关系调查中发现除少数耐荫藤本外绝大多数藤本的生长会随群落中的乔木一起向上生长,并认为虽然藤本植物胸径大小与支持木胸径之间的关系受光照、土壤肥力、群落结构、群落演替等诸多因素的影响,但无论在哪种条件下,它们的胸径大小都存在一定的相关关系,当群落演替到物种多样性和密闭性较高时,乔木会通过生长高度来获取更多的光照和水分,从而使得攀附藤本的地径和高度随之生长,一定程度上也说明了其正相关性。刘晋仙等[33]在海南霸王岭山地原始森林与伐后林的调查中发现,伐后林比原始林的树木拥有更高的物种多度和丰富度,但藤本的物种丰富度和多度差异不显著,而平均每木藤本数与支持木高度之间有着明显的正相关,都是随着树木高度的增加攀附的藤本数也随之增加,而伐后林的增长幅度明显高于原始林。藤本植物的过量增加会抢占其他植被的生态位,导致乔灌木多样性、均匀度降低。择伐干扰后冠层郁闭度相对降低,一定程度上促进了森林底层植被的生长[33-34]。
4 展 望
藤本植物是热带地区森林生态系统中的重要组成部分,在维持热带森林生物多样性、结构和功能上有着极为重要的作用[7]。然而,本次调查发现藤本植物的大量繁殖对次生林群落的正常生长发育有一定的抑制作用,会降低乔灌木的多样性与均匀度,从而阻碍次生林群落的天然更新与正常演替。本研究采用的是空间代替时间的方法,对伐藤后样方内树种的长期变化规律研究具有一定的局限性。今后,可对样方内的物种进行长期观测,以推测该区域次生林的演替趋势,为避免该区域内的藤本植物造成过量干扰,特别是控制金钟藤、葛的数量,从而提高该区域内植被恢复潜力,促进次生林的天然更新,推动该区域次生林向顶级群落演替。
猜你喜欢
现代农村科技(2022年5期)2022-11-18
安徽农业科学(2021年7期)2021-04-29
环球时报(2021-01-27)2021-01-27
老年教育(2020年5期)2020-05-02
少儿科学周刊·儿童版(2019年5期)2019-08-01
读友·少年文学(清雅版)(2018年2期)2018-09-10
现代农业研究(2017年11期)2018-01-12
环球时报(2017-12-19)2017-12-19
热带农业科学(2017年5期)2017-06-12
现代农业研究(2016年8期)2016-12-02
